
281 results in 35Kxx
Travelling wavefronts for the Belousov–Zhabotinsky system with non-local delayed interaction
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 10 October 2024, pp. 1-37
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Boundary crossing problems and functional transformations for Ornstein–Uhlenbeck processes
- Part of
-
- Journal:
- Journal of Applied Probability / Volume 62 / Issue 1 / March 2025
- Published online by Cambridge University Press:
- 07 October 2024, pp. 395-421
- Print publication:
- March 2025
-
- Article
-
- You have access
- HTML
- Export citation
Stabilization in a chemotaxis system modelling T-cell dynamics with simultaneous production and consumption of signals
- Part of
-
- Journal:
- European Journal of Applied Mathematics , First View
- Published online by Cambridge University Press:
- 18 September 2024, pp. 1-14
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
In a smoothly bounded domain
 $\Omega \subset \mathbb{R}^n$,
$\Omega \subset \mathbb{R}^n$,  $n\ge 1$, this manuscript considers the homogeneous Neumann boundary problem for the chemotaxis systemwith parameter
$n\ge 1$, this manuscript considers the homogeneous Neumann boundary problem for the chemotaxis systemwith parameter \begin{eqnarray*} \left \{ \begin{array}{l} u_t = \Delta u - \nabla \cdot (u\nabla v), \\[5pt] v_t = \Delta v + u - \alpha uv, \end{array} \right . \end{eqnarray*}
\begin{eqnarray*} \left \{ \begin{array}{l} u_t = \Delta u - \nabla \cdot (u\nabla v), \\[5pt] v_t = \Delta v + u - \alpha uv, \end{array} \right . \end{eqnarray*} $\alpha \gt 0$ and with coincident production and uptake of attractants, as recently emphasized by Dallaston et al. as relevant for the understanding of T-cell dynamics.
$\alpha \gt 0$ and with coincident production and uptake of attractants, as recently emphasized by Dallaston et al. as relevant for the understanding of T-cell dynamics.It is shown that there exists
 $\delta _\star =\delta _\star (n)\gt 0$ such that for any given
$\delta _\star =\delta _\star (n)\gt 0$ such that for any given  $\alpha \ge \frac{1}{\delta _\star }$ and for any suitably regular initial data satisfying
$\alpha \ge \frac{1}{\delta _\star }$ and for any suitably regular initial data satisfying  $v(\cdot, 0)\le \delta _\star$, this problem admits a unique classical solution that stabilizes to the constant equilibrium
$v(\cdot, 0)\le \delta _\star$, this problem admits a unique classical solution that stabilizes to the constant equilibrium  $(\frac{1}{|\Omega |}\int _\Omega u(\cdot, 0), \, \frac{1}{\alpha })$ in the large time limit.
$(\frac{1}{|\Omega |}\int _\Omega u(\cdot, 0), \, \frac{1}{\alpha })$ in the large time limit.








The roles of nonlinear diffusion, haptotaxis and ECM remodelling in determining the global solvability of a cancer invasion model
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 13 September 2024, pp. 1-32
-
- Article
-
- You have access
- HTML
- Export citation
-
In this paper, we consider the following PDE-ODE system modelling cancer invasion with slow diffusion and ECM remodelling,
\[ \begin{cases} u_t=\Delta u^m-\chi\nabla\cdot(u\nabla v)-\xi\nabla\cdot(u\nabla\omega)+\mu u(1-u-\omega), \\ v_t=\Delta v+u-v, \\ \omega_t={-}v\omega+\eta \omega(1-u-\omega). \end{cases} \]For the special case $\eta =0$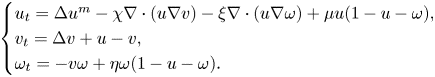
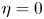 , fruitful results have been achieved since Tao and Winkler's work in 2011. However, there is no any progress for the general case $\eta >0$
, fruitful results have been achieved since Tao and Winkler's work in 2011. However, there is no any progress for the general case $\eta >0$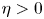 in the past ten years. In this paper, we analysed some commonly used research methods when $\eta =0$
in the past ten years. In this paper, we analysed some commonly used research methods when $\eta =0$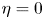 , and found that these methods are completely unsuitable for situations where $\eta >0$
, and found that these methods are completely unsuitable for situations where $\eta >0$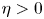 . By introducing some new forms of functionals, we reconstruct the relationship between the haptotactic term and the nonlinear diffusion term, and ultimately prove the global existence of weak solutions. This result improves and perfects a series of works previously presented in the literature.
. By introducing some new forms of functionals, we reconstruct the relationship between the haptotactic term and the nonlinear diffusion term, and ultimately prove the global existence of weak solutions. This result improves and perfects a series of works previously presented in the literature.
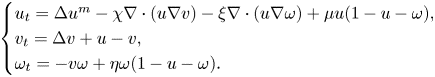
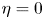
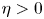
Asymptotic profiles of a nonlocal dispersal SIS epidemic model with saturated incidence
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 14 May 2024, pp. 1-33
-
- Article
-
- You have access
- HTML
- Export citation
Unified framework for the separation property in binary phase-segregation processes with singular entropy densities
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 36 / Issue 1 / February 2025
- Published online by Cambridge University Press:
- 09 May 2024, pp. 40-67
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
This paper investigates the separation property in binary phase-segregation processes modelled by Cahn-Hilliard type equations with constant mobility, singular entropy densities and different particle interactions. Under general assumptions on the entropy potential, we prove the strict separation property in both two and three-space dimensions. Namely, in 2D, we notably extend the minimal assumptions on the potential adopted so far in the literature, by only requiring a mild growth condition of its first derivative near the singular points
 $\pm 1$, without any pointwise additional assumption on its second derivative. For all cases, we provide a compact proof using De Giorgi’s iterations. In 3D, we also extend the validity of the asymptotic strict separation property to the case of fractional Cahn-Hilliard equation, as well as show the validity of the separation when the initial datum is close to an ‘energy minimizer’. Our framework offers insights into statistical factors like particle interactions, entropy choices and correlations governing separation, with broad applicability.
$\pm 1$, without any pointwise additional assumption on its second derivative. For all cases, we provide a compact proof using De Giorgi’s iterations. In 3D, we also extend the validity of the asymptotic strict separation property to the case of fractional Cahn-Hilliard equation, as well as show the validity of the separation when the initial datum is close to an ‘energy minimizer’. Our framework offers insights into statistical factors like particle interactions, entropy choices and correlations governing separation, with broad applicability.

Threshold dynamics scenario of a plants-pollinators cooperative system with impulsive effect on a periodically evolving domain
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 35 / Issue 6 / December 2024
- Published online by Cambridge University Press:
- 02 May 2024, pp. 797-822
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Global boundedness and large time behaviour in a higher-dimensional quasilinear chemotaxis system with consumption of chemoattractant
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 30 April 2024, pp. 1-26
-
- Article
-
- You have access
- HTML
- Export citation
-
This paper deals with the following quasilinear chemotaxis system with consumption of chemoattractant
\[ \left\{\begin{array}{@{}ll} u_t=\Delta u^{m}-\nabla\cdot(u\nabla v),\quad & x\in \Omega,\quad t>0,\\ v_t=\Delta v-uv,\quad & x\in \Omega,\quad t>0\\ \end{array}\right. \]in a bounded domain $\Omega \subset \mathbb {R}^N(N=3,\,4,\,5)$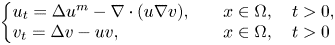
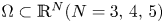 with smooth boundary $\partial \Omega$
with smooth boundary $\partial \Omega$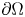 . It is shown that if $m>\max \{1,\,\frac {3N-2}{2N+2}\}$
. It is shown that if $m>\max \{1,\,\frac {3N-2}{2N+2}\}$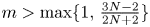 , for any reasonably smooth nonnegative initial data, the corresponding no-flux type initial-boundary value problem possesses a globally bounded weak solution. Furthermore, we prove that the solution converges to the spatially homogeneous equilibrium $(\bar {u}_0,\,0)$
, for any reasonably smooth nonnegative initial data, the corresponding no-flux type initial-boundary value problem possesses a globally bounded weak solution. Furthermore, we prove that the solution converges to the spatially homogeneous equilibrium $(\bar {u}_0,\,0)$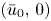 in an appropriate sense as $t\rightarrow \infty$
in an appropriate sense as $t\rightarrow \infty$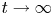 , where $\bar {u}_0=\frac {1}{|\Omega |}\int _\Omega u_0$
, where $\bar {u}_0=\frac {1}{|\Omega |}\int _\Omega u_0$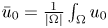 . This result not only partly extends the previous global boundedness result in Fan and Jin (J. Math. Phys. 58 (2017), 011503) and Wang and Xiang (Z. Angew. Math. Phys. 66 (2015), 3159–3179) to $m>\frac {3N-2}{2N}$
. This result not only partly extends the previous global boundedness result in Fan and Jin (J. Math. Phys. 58 (2017), 011503) and Wang and Xiang (Z. Angew. Math. Phys. 66 (2015), 3159–3179) to $m>\frac {3N-2}{2N}$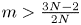 in the case $N\geq 3$
in the case $N\geq 3$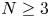 , but also partly improves the global existence result in Zheng and Wang (Discrete Contin. Dyn. Syst. Ser. B 22 (2017), 669–686) to $m>\frac {3N}{2N+2}$
, but also partly improves the global existence result in Zheng and Wang (Discrete Contin. Dyn. Syst. Ser. B 22 (2017), 669–686) to $m>\frac {3N}{2N+2}$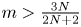 when $N\geq 2$
when $N\geq 2$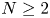 .
.
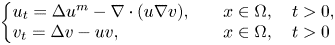
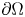
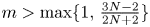
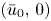
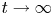
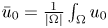
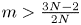
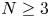
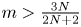
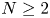
Eternal solutions to a porous medium equation with strong non-homogeneous absorption. Part I: radially non-increasing profiles
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 14 March 2024, pp. 1-22
-
- Article
-
- You have access
- HTML
- Export citation
-
Existence of specific eternal solutions in exponential self-similar form to the following quasilinear diffusion equation with strong absorption
\[ \partial_t u=\Delta u^m-|x|^{\sigma}u^q, \]posed for $(t,\,x)\in (0,\,\infty )\times \mathbb {R}^N$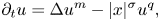
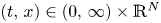 , with $m>1$
, with $m>1$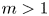 , $q\in (0,\,1)$
, $q\in (0,\,1)$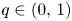 and $\sigma =\sigma _c:=2(1-q)/ (m-1)$
and $\sigma =\sigma _c:=2(1-q)/ (m-1)$ is proved. Looking for radially symmetric solutions of the form\[ u(t,x)={\rm e}^{-\alpha t}f(|x|\,{\rm e}^{\beta t}), \quad \alpha=\frac{2}{m-1}\beta, \]we show that there exists a unique exponent $\beta ^*\in (0,\,\infty )$
is proved. Looking for radially symmetric solutions of the form\[ u(t,x)={\rm e}^{-\alpha t}f(|x|\,{\rm e}^{\beta t}), \quad \alpha=\frac{2}{m-1}\beta, \]we show that there exists a unique exponent $\beta ^*\in (0,\,\infty )$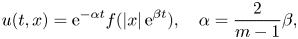
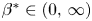 for which there exists a one-parameter family $(u_A)_{A>0}$
for which there exists a one-parameter family $(u_A)_{A>0}$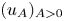 of solutions with compactly supported and non-increasing profiles $(f_A)_{A>0}$
of solutions with compactly supported and non-increasing profiles $(f_A)_{A>0}$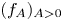 satisfying $f_A(0)=A$
satisfying $f_A(0)=A$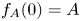 and $f_A'(0)=0$
and $f_A'(0)=0$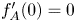 . An important feature of these solutions is that they are bounded and do not vanish in finite time, a phenomenon which is known to take place for all non-negative bounded solutions when $\sigma \in (0,\,\sigma _c)$
. An important feature of these solutions is that they are bounded and do not vanish in finite time, a phenomenon which is known to take place for all non-negative bounded solutions when $\sigma \in (0,\,\sigma _c)$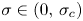 .
.
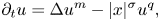
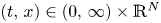
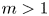
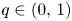

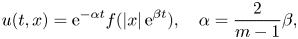
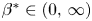
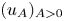
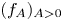
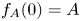
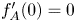
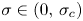
Decay at infinity for solutions to some fractional parabolic equations
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 14 March 2024, pp. 1-37
-
- Article
-
- You have access
- HTML
- Export citation
-
For $s\in [\tfrac {1}{2},\, 1)$
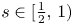 , let $u$
, let $u$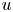 solve $(\partial _t - \Delta )^s u = Vu$
solve $(\partial _t - \Delta )^s u = Vu$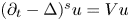 in $\mathbb {R}^{n} \times [-T,\, 0]$
in $\mathbb {R}^{n} \times [-T,\, 0]$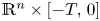 for some $T>0$
for some $T>0$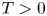 where $||V||_{ C^2(\mathbb {R}^n \times [-T, 0])} < \infty$
where $||V||_{ C^2(\mathbb {R}^n \times [-T, 0])} < \infty$ . We show that if for some $0<\mathfrak {K} < T$
. We show that if for some $0<\mathfrak {K} < T$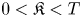 and $\epsilon >0$
and $\epsilon >0$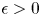 \[ {\unicode{x2A0D}}-_{[-\mathfrak{K},\, 0]} u^2(x, t) {\rm d}t \leq Ce^{-|x|^{2+\epsilon}}\ \forall x \in \mathbb{R}^n, \]then $u \equiv 0$
\[ {\unicode{x2A0D}}-_{[-\mathfrak{K},\, 0]} u^2(x, t) {\rm d}t \leq Ce^{-|x|^{2+\epsilon}}\ \forall x \in \mathbb{R}^n, \]then $u \equiv 0$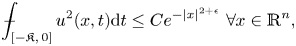
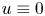 in $\mathbb {R}^{n} \times [-T,\, 0]$
in $\mathbb {R}^{n} \times [-T,\, 0]$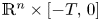 .
.
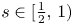
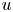
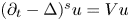
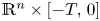
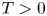

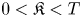
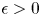
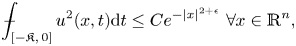
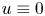
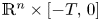
Unbounded Sturm attractors for quasilinear parabolic equations
- Part of
-
- Journal:
- Proceedings of the Edinburgh Mathematical Society / Volume 67 / Issue 2 / May 2024
- Published online by Cambridge University Press:
- 08 March 2024, pp. 542-565
-
- Article
-
- You have access
- HTML
- Export citation
Bifurcations and pattern formation in a host–parasitoid model with nonlocal effect
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 04 March 2024, pp. 1-40
-
- Article
-
- You have access
- HTML
- Export citation
On determining and breaking the gauge class in inverse problems for reaction-diffusion equations
- Part of
-
- Journal:
- Forum of Mathematics, Sigma / Volume 12 / 2024
- Published online by Cambridge University Press:
- 26 February 2024, e25
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We investigate an inverse boundary value problem of determination of a nonlinear law for reaction-diffusion processes, which are modeled by general form semilinear parabolic equations. We do not assume that any solutions to these equations are known a priori, in which case the problem has a well-known gauge symmetry. We determine, under additional assumptions, the semilinear term up to this symmetry in a time-dependent anisotropic case modeled on Riemannian manifolds, and for partial data measurements on
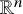 ${\mathbb R}^n$.
${\mathbb R}^n$.Moreover, we present cases where it is possible to exploit the nonlinear interaction to break the gauge symmetry. This leads to full determination results of the nonlinear term. As an application, we show that it is possible to give a full resolution to classes of inverse source problems of determining a source term and nonlinear terms simultaneously. This is in strict contrast to inverse source problems for corresponding linear equations, which always have the gauge symmetry. We also consider a Carleman estimate with boundary terms based on intrinsic properties of parabolic equations.
Global dynamics and spatiotemporal heterogeneity of a preytaxis model with prey-induced acceleration
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 35 / Issue 5 / October 2024
- Published online by Cambridge University Press:
- 26 January 2024, pp. 601-633
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
On degenerate reaction-diffusion epidemic models with mass action or standard incidence mechanism
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 35 / Issue 5 / October 2024
- Published online by Cambridge University Press:
- 22 January 2024, pp. 634-661
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
The fully parabolic multi-species chemotaxis system in
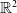 $\mathbb{R}^{2}$
$\mathbb{R}^{2}$
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 35 / Issue 5 / October 2024
- Published online by Cambridge University Press:
- 19 January 2024, pp. 675-706
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
This article is devoted to the analysis of the parabolic–parabolic chemotaxis system with multi-components over
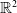 $\mathbb{R}^2$. The optimal small initial condition on the global existence of solutions for multi-species chemotaxis model in the fully parabolic situation had not been attained as far as the author knows. In this paper, we prove that under the sub-critical mass condition, any solutions to conflict-free system exist globally. Moreover, the global existence of solutions to system with strong self-repelling effect has been discussed even for large initial data. The proof is based on the modified free energy functional and the Moser–Trudinger inequality for system.
$\mathbb{R}^2$. The optimal small initial condition on the global existence of solutions for multi-species chemotaxis model in the fully parabolic situation had not been attained as far as the author knows. In this paper, we prove that under the sub-critical mass condition, any solutions to conflict-free system exist globally. Moreover, the global existence of solutions to system with strong self-repelling effect has been discussed even for large initial data. The proof is based on the modified free energy functional and the Moser–Trudinger inequality for system.
Adhesion and volume filling in one-dimensional population dynamics under Dirichlet boundary condition
- Part of
-
- Journal:
- Proceedings of the Royal Society of Edinburgh. Section A: Mathematics , First View
- Published online by Cambridge University Press:
- 08 January 2024, pp. 1-49
-
- Article
-
- You have access
- HTML
- Export citation
Solvability of Hessian quotient equations in exterior domains
- Part of
-
- Journal:
- Canadian Journal of Mathematics / Volume 77 / Issue 1 / February 2025
- Published online by Cambridge University Press:
- 14 December 2023, pp. 118-148
- Print publication:
- February 2025
-
- Article
- Export citation
-
In this paper, we study the Dirichlet problem of Hessian quotient equations of the form
 $S_k(D^2u)/S_l(D^2u)=g(x)$ in exterior domains. For
$S_k(D^2u)/S_l(D^2u)=g(x)$ in exterior domains. For  $g\equiv \mbox {const.}$, we obtain the necessary and sufficient conditions on the existence of radially symmetric solutions. For g being a perturbation of a generalized symmetric function at infinity, we obtain the existence of viscosity solutions by Perron’s method. The key technique we develop is the construction of sub- and supersolutions to deal with the non-constant right-hand side g.
$g\equiv \mbox {const.}$, we obtain the necessary and sufficient conditions on the existence of radially symmetric solutions. For g being a perturbation of a generalized symmetric function at infinity, we obtain the existence of viscosity solutions by Perron’s method. The key technique we develop is the construction of sub- and supersolutions to deal with the non-constant right-hand side g.


Analysis and numerical simulations of travelling waves due to plant–soil negative feedback
- Part of
-
- Journal:
- European Journal of Applied Mathematics / Volume 35 / Issue 4 / August 2024
- Published online by Cambridge University Press:
- 07 December 2023, pp. 554-565
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Anisotropic flow, entropy, and
 $L^p$-Minkowski problem
$L^p$-Minkowski problem
- Part of
-
- Journal:
- Canadian Journal of Mathematics / Volume 77 / Issue 1 / February 2025
- Published online by Cambridge University Press:
- 28 November 2023, pp. 1-20
- Print publication:
- February 2025
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We provide a natural simple argument using anistropic flows to prove the existence of weak solutions to Lutwak’s
 $L^p$-Minkowski problem on
$L^p$-Minkowski problem on  $S^n$ which were obtained by other methods.
$S^n$ which were obtained by other methods.




