



Introduction
During development, orthodox seeds acquire a set of physiological traits of significance to survival and regeneration: the ability to germinate before and after drying and the capacity to survive long periods of air-dry storage (Bewley et al., Reference Bewley, Bradford, Hilhorst and Nonogaki2013; Waterworth et al., Reference Waterworth, Bray and West2015). However, seed deterioration begins soon after seed dispersal and then during storage. Over time, viability loss is inevitable (Bewley et al., Reference Bewley, Bradford, Hilhorst and Nonogaki2013), even under conditions recommended for genebanks, likely as a result of cumulative oxidative stress (Kranner et al., Reference Kranner, Birtić, Anderson and Pritchard2006). Uncertainty in how long a seed lot of a species might survive in storage makes it critical to periodically monitor viability during storage (FAO, 2014). It is also helpful to understand inter- and intra-specific differences in seed longevity and to potentially customise monitoring intervals so that seed lots expected to lose viability sooner are tested before those expected to remain viable for longer (Hay and Whitehouse, Reference Hay and Whitehouse2017). To assess relative seed longevity within a reasonable time frame (i.e. weeks to months), samples of seeds are aged under standard, relatively high moisture and temperature conditions (Hay et al., Reference Hay, Valdez, Lee and Sta Cruz2019). This has been used to rank relative seed longevity of hundreds of species (Probert et al., Reference Probert, Daws and Hay2009; Mondoni et al., Reference Mondoni, Probert, Rossi, Vegini and Hay2011; White et al., Reference White, Hay, Abeli and Mondoni2023) and hundreds of accessions destined for genebank storage (Lee et al., Reference Lee, Velasco-Punzalan, Pacleb, Valdez, Kretzschmar, McNally, Ismail, Sta. Cruz, Sackville Hamilton and Hay2019).
As seeds age in storage, their ability to produce normal healthy seedlings may decline before the reduction in radicle emergence (Rao et al., Reference Rao, Roberts and Ellis1987; Bradford et al., Reference Bradford, Tarquis and Duran1993; Bewley et al., Reference Bewley, Bradford, Hilhorst and Nonogaki2013). Therefore, relying solely on radicle protrusion as an estimate of survival after storage might overestimate the seeds' ability to produce a normal healthy seedling (Ellis and Roberts, Reference Ellis and Roberts1981). According to the guidelines provided by the International Seed Testing Association (ISTA), normal germination, which refers to a seed's ability to develop into a satisfactory plant (ISTA, 2018), is considered the most reliable and recommended test of seed quality over radicle emergence. This principle is also supported by findings in some wild plants. For example, in Delphinium and Salvia seeds, cotyledon following by radicle emergence was identified as a better indicator of seed viability after storage (Kwong et al., Reference Kwong, Stodolski, Mari, Gurusinghe and Bradford2001), as well as in Salix seeds after various storage treatments, including cryopreservation (Popova et al., Reference Popova, Kim, Han, Moltchanova, Pritchard and Hong2013). However, in most, if not all, seed longevity studies on wild plants, radicle emergence has been the main criterion used to determine a seed's viability (see e.g. Long et al., Reference Long, Panetta, Steadman, Probert, Bekker, Brooks and Adkins2008; Kochanek et al., Reference Kochanek, Steadman, Probert and Adkins2009; Probert et al., Reference Probert, Daws and Hay2009; Mondoni et al., Reference Mondoni, Probert, Rossi, Vegini and Hay2011; Merritt et al., Reference Merritt, Martyn, Ainsley, Young, Seed, Thorpe, Hay, Commander, Shackelford, Offord and Dixon2014; Bernareggi et al., Reference Bernareggi, Carbognani, Petraglia and Mondoni2015; Seglias, Reference Seglias2022; Sommerville et al., Reference Sommerville, Newby, Yenson and Offord2023; White et al., Reference White, Hay, Abeli and Mondoni2023). In this context, germination studies on wild plant species do not adhere to the guidelines on crop species. Different standards have also been applied in other studies, such as those from the European Native Seed Conservation Network (ENSCONET, 2009), the Australia Flora Bank Guidelines (FloraBank, 1999) and the US Seeds of Success program (Bureau of Land Management, 2018). The reasons for having different standards for native seeds as opposed to crops are extensively discussed by Pedrini and Dixon (Reference Pedrini and Dixon2020). It is unquestionable that checking normal seedling development is a valuable means of assessing post-storage viability as it is a better indicator of potential seedling establishment. But it is not always feasible to do this for wild plants, in part because what is considered a ‘normal seedling’ that is able to grow into a healthy plant has not been documented for many wild species. The attrition, or loss, of individuals as germination progresses from radicle emergence to early seedling emergence (i.e. radicle and cotyledon) has the potential to impact the interpretation of seed viability, depending on which parameter is measured. Indeed, there is the possibility of overestimating seed viability when using radicle emergence alone. The extent of this problem may depend on the species and seed age (Ellis and Roberts, Reference Ellis and Roberts1981; Tarquis and Bradford, Reference Tarquis and Bradford1992; Popova et al., Reference Popova, Kim, Han, Moltchanova, Pritchard and Hong2013). Of particular concern is any overestimation of seed longevity (OESL) for the most vulnerable plant species, including those already identified as producing short-lived seeds in storage, for example, species from cool wet climates (Probert et al., Reference Probert, Daws and Hay2009) or, more specifically, understorey herbs and alpine species (Ali et al., Reference Ali, Probert, Hay, Davies and Stuppy2007; Mondoni et al., Reference Mondoni, Probert, Rossi, Vegini and Hay2011). In this study, we examine the longevity of seeds from 33 alpine species (including two subspecies of one species, Silene vulgaris) under standard storage conditions, evaluating both radicle emergence (R) and radicle plus cotyledon emergence (R + C) to provide insights into the potential OESL.
Material and methods
Studied species and seed collection
The studied species inhabit two of the Natura 2000 habitat types: ‘6230 – Nardus-rich species grasslands’, formed on various types of siliceous rocks with acidic pH and ‘6170 – Alpine and subalpine calcareous grasslands’, formed on calcareous bedrocks with alkaline pH (92/43/CEE ‘Habitat’ Directive classification) (European Commission, 2007).
Fresh mature seeds of the target species were collected based on their occurrence and abundance in these two habitats at the time of natural seed dispersal at the Stelvio National Park (close to Ubraill pass, 46°32′30.1″N, 10°25′46.1″E; 2,565 m a.s.l.) and in Dosdè Valley (46°25′06.5″N, 10°11′59.8″E; 2,225 m a.s.l.) (Sondrio Province, Italy) during August 2015 following the ENSCONET (2009) guidelines for responsible seed collection practices. After collection, seeds were held in the drying room [15% relative humidity (RH), 15°C] at the Plant Germplasm Bank of the University of Pavia (Italy) for 1–2 months until the commencement of the experiments. Seed viability of all species was determined through standard germination testing and 33 species (14 families; Table 1) having highly viable seeds were selected for this study. Seed mass (average seed weight of three replicates of 50 seeds, converted to 1000 seed weight) of each species was measured and seed morphology type of each species (at least to the genus level) was derived from Martin (1946) (Table 1).
Table 1. Details of the species used in the study and seed characteristics, together with the accepted model when comparing the post-storage germination survival curves (equation 1) when the germination criterion was radicle emergence (R) or radicle and cotyledon emergence (R + C)

Experimental storage
Seed longevity was determined using a standard rapid ageing protocol (Newton et al., Reference Newton, Hay and Probert2009). Samples of seeds from each species were removed from the drying room after 1–2 months and equilibrated to the laboratory environment over 5 d, monitored using a data logger (approximately 20°C; 50% RH, TinyTag DataLogger). Thereafter, seeds of each species were placed separately inside a Petri dish as a single seed layer and sealed in a 300 × 300 × 130 mm electrical enclosure box (Ensto UK Ltd, Southampton, UK), over a non-saturated solution of LiCl (anhydrous, Laboratory Reagent Grade, Fisher Scientific UK Ltd.) at 60% RH (Hay et al., Reference Hay, Adams, Manger and Probert2008) and placed in an oven (Binder FD53, Tuttlingen, Germany) in the dark at 45°C. One sample of 60 seeds of each species was retrieved from the large sample of seeds in the electrical enclosure box after 0, 1, 3, 5, 7, 10, 15, 20, 30, 40 and 50 d for germination testing. For each species, the seeds were sown as three subsamples of 20 seeds on 1% agar in distilled water containing 0.00072 mol L−1 GA3 (Sigma-Aldrich Company Ltd., Dorset, UK) in 55 mm-diameter Petri dishes and incubated at 25°C light (12 h) and 15°C dark (12 h) in a LMS 250A cooled incubator (LMS Ltd., Sevenoaks, UK). Radicle emergence (R) of 1–2 mm in length and subsequent fully opened cotyledonary leaf emergence (R + C) were recorded separately every 5 d for 40 or 70 d after sowing, depending on species and seed quality. Excess water accumulating on the Petri dish lids during the incubation period was removed in order to minimize the risk of damping off.
Data analysis
Probit analysis was performed using the FITNONLINEAR directive in GenStat 19th Edition (VSN International Ltd., Hemel Hempstead, UK) to simultaneously fit the seed viability equation to the data for both the number of seeds showing radicle emergence (R) and the number of seeds going on to produce fully open cotyledons (R + C):

where v is the viability according to R or R + C (in normal equivalent deviates, NED) of the seed lot after p days in storage, Ki is the initial viability (NED) of the seed lot and σ is the time (d) for viability to fall by 1 NED (i.e. the standard deviation of the normal distribution of seed deaths over time) (Ellis and Roberts, Reference Ellis and Roberts1980). In the same analysis, the effects of constraining Ki and/or σ to the same value for the different germination criteria on the residual deviance were compared using approximate F-tests. In addition, estimates were made of the time for viability to fall to 75 or 50% (p 75 and p 50, respectively), again using the two different criteria. The p 75 was estimated as it is the viability standard for the seed bank at the Department of Earth and Environmental Sciences, University of Pavia; when the viability falls below 75%, recollection of the species/population will be considered, according to conservation priorities.
When it was not possible to constrain the parameters of equation (1) (Ki and σ) to the same values for the two germination criteria (R or R + C) due to a significant increase in residual deviance, further analyses were implemented. In these cases, the OESL resulting from using R as the germination criterion (instead of R + C) was estimated both in relation to the number of days and percentage using the following equations:


with x equal to 50 or 75. These calculations were also made in GenStat and hence the standard errors of the estimates of OESLx, based on all the germination data used to fit each survival curve, were also provided.
In addition, when it was not possible to constrain Ki and σ to the same values for the two germination criteria (R or R + C), the two fitted versions of equation (1) were used to estimate the viability using R as the germination criterion as a function of the viability using R + C as the germination criterion, that is,

Results
Seed survival curves
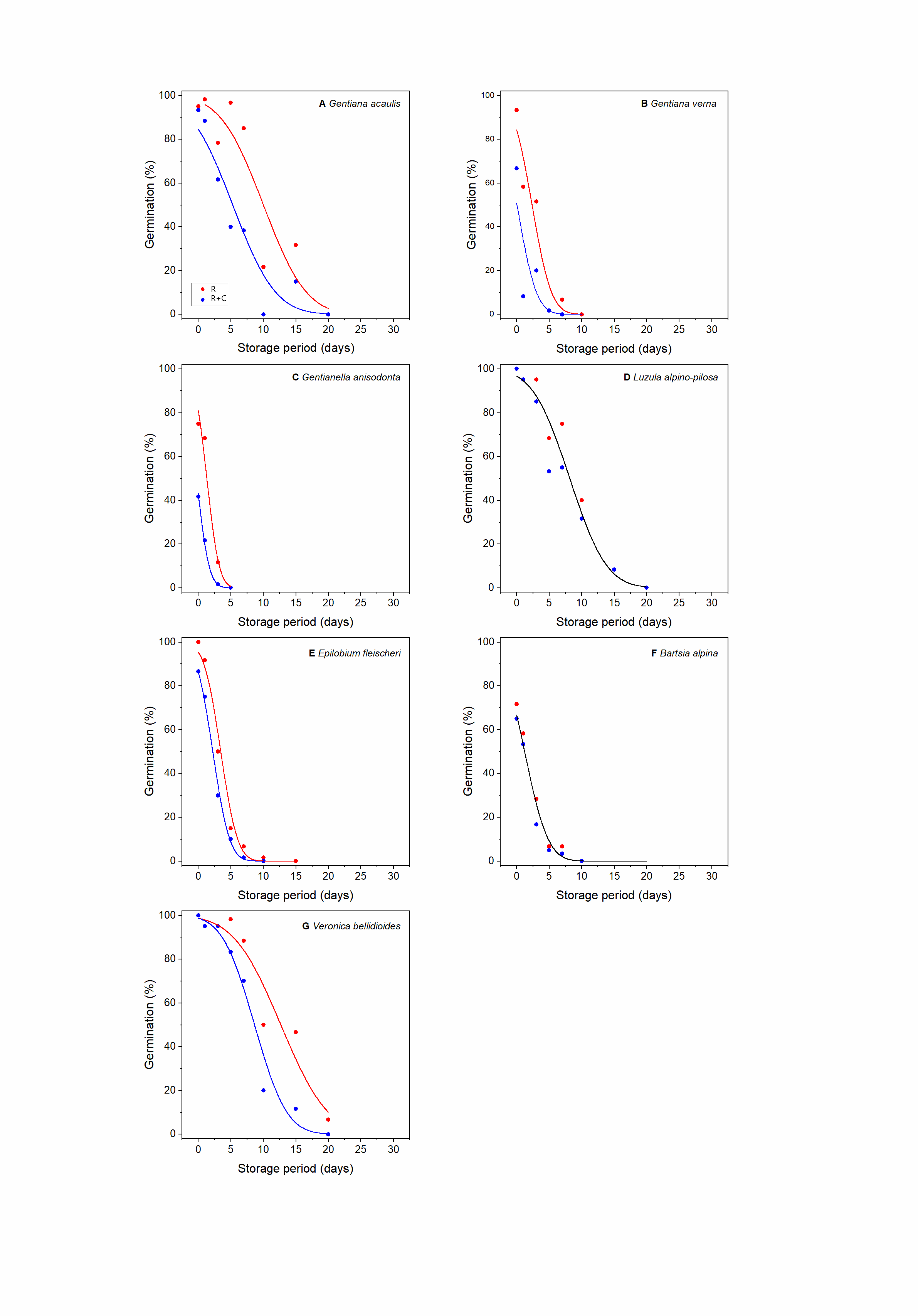
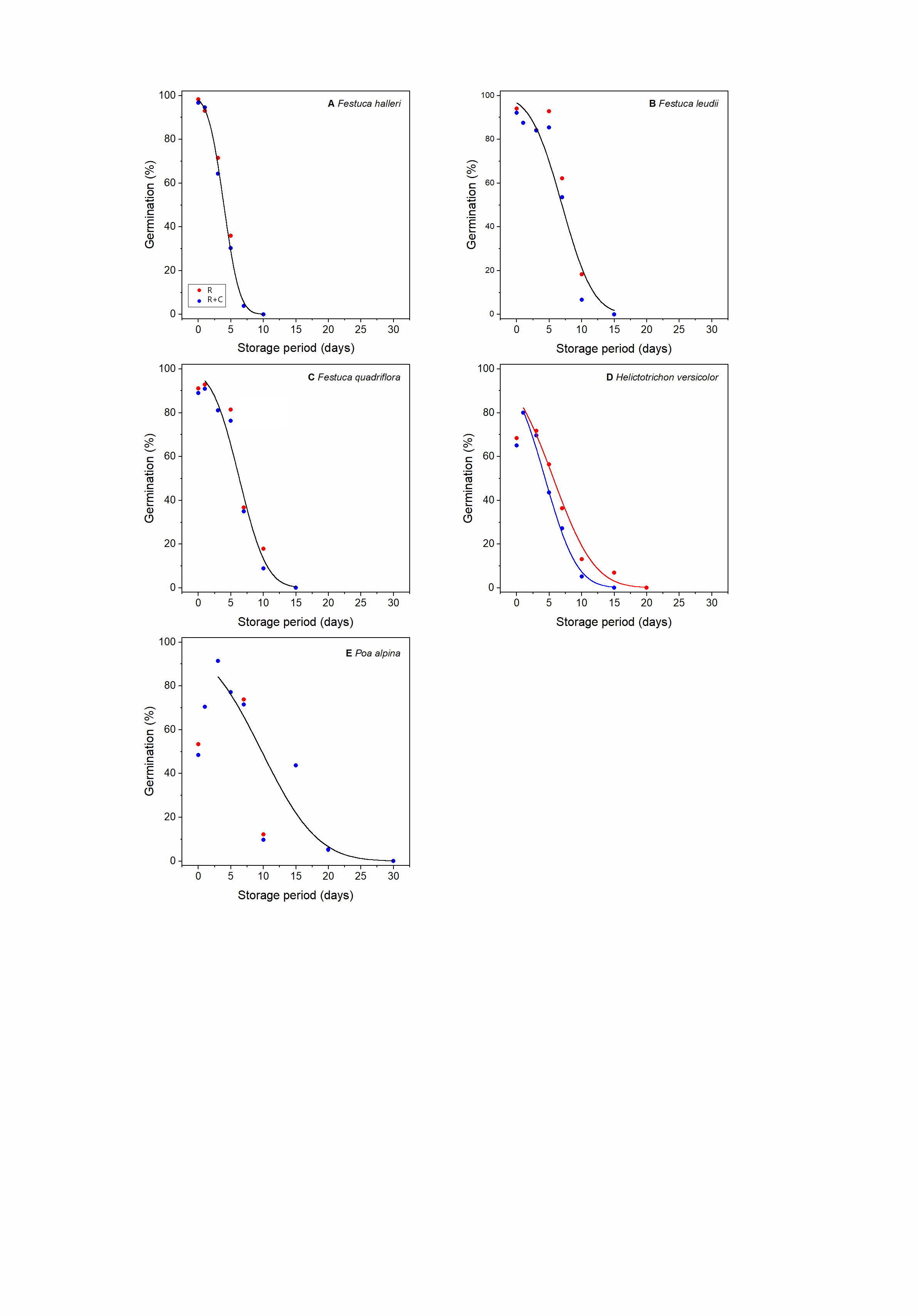
Seed viability declined with increasing duration of the ageing treatment in all species (Supplementary Figs S1–S6). For every sampling time and across all species, R was higher (most samples) or equal to the count of R + C (Supplementary Figs S1–S6; Fig. 1). In some cases, the differences were large (e.g. Silene rupestris, see Supplementary Fig. S3B), whereas for 16 species, the differences were very small throughout the sampling period and it was possible to fit a single survival curve (‘one line’) to the R and R + C data without a significant increase in residual deviance (P > 0.05); this group includes all Rosaceae, four out of five Poaceae, five out of seven Asteraceae and the single representative species of the Juncaceae, Orobanchaceae and Scrophulariaceae (Table 1). For those species where a single line could not be fitted, a common slope model was the most frequently accepted model (12 species), while the common intercept model was accepted for just four species. Only for Saxifraga stellaris, was it not possible to constrain either the slope or the intercept of the R and R + C survival curves.

Figure. 1. Observed and fitted (equation 4) relationships between radicle (R) and radicle plus cotyledon emergence (R + C) for seeds of 33 alpine species from 14 different families, as indicated. Seeds were aged for different periods of time at 45°C and 60% RH. The black line is the line of equality (i.e. R = R + C); the curved lines show the difference between the fitted survival curves for R and R + C for the respective species where it was not possible to constrain to a common line for R and R + C.
Estimates and OESL
Some seed lots, most notably those of Bartsia alpina, Gentianella anisodonta and Gentiana verna, had low initial viability, hence estimates for viability to fall to 75 or even 50% were negative (Tables 2–3). Using R as the criterion for germination, the estimate for p 50(R) ranged from 1.24 d for seeds of B. alpina (Table 2) to 41.71 d for seeds of Silene vulgaris ssp. glareosa (Table 3).
Table 2. Estimates (±SE) of seed longevity parameters, Ki and σ (equation 1) and the time for viability to fall to 75 and 50% (p 75 and p 50) from laboratory seed storage experiments in which seeds were stored at 60% RH and 45°C with samples taken at regular intervals for a germination test

The parameters shown are the results of fitting a model with the same estimates for Ki and σ (‘common line’) when the germination criterion was radicle (R) or radicle plus cotyledon emergence (R + C) (Supplementary Figs S1–S6).
a Estimate was negative (initial viability <75%).
Table 3. Estimates (±SE) of seed longevity parameters, Ki and σ (equation 1), the time for viability to fall to 75 and 50% (p 75 and p 50) from laboratory seed storage experiments, and the overestimation of p 75 and p 50 (OESL) when the germination criterion used was radicle emergence (R) rather than radicle plus cotyledon emergence (R + C) (equations 2 and 3)

Seeds were stored at 60% RH and 45°C with samples taken at regular intervals for a germination test. The parameters shown are the results of fitting a model with the same estimate for σ (‘common slope’) when the germination criterion was radicle (R) or radicle plus cotyledon emergence (R + C) (Supplementary Figs S1, S2 and S4–S6).
a Estimate was negative (initial viability <75%).
b Estimate was negative (initial viability <50%).
n.a., calculation could not be performed due to the negative estimate of p 50 or p 75.
Since p 50 is usually used as the relative measure of longevity in comparative studies, we considered the species rankings depending on whether R or R + C was used as the germination criterion. The species with the largest rank shift was Primula farinosa, changing from the 8th longest-lived seeds based on R to the 13th shortest-lived seeds when using R + C (Fig. 2). Other species that also fell in the ranking were Gentiana acaulis (from 11th to 18th longest-lived) and Phyteuma orbiculare (from 22nd to 29th). Species that moved up the ranking included Campanula barbata, Luzula alpino-pilosa and Dryas octopetala (moving from 14th to 9th, 15th to 11th and 21st to 17th, respectively). There were some changes in the rankings and in the shift in rankings depending on the germination criterion when p 75 rather than p 50 was used as the measure of longevity (cf. Fig. 2 and Supplementary Fig. 7).

Figure. 2. Species ranking for relative seed longevity depending on whether the germination criterion is radicle emergence (R) or radicle plus cotyledon emergence (R + C). The y-axis scale shows p 50, the time for viability to fall to 50% when seeds are stored at 45°C and 60% RH (Tables 2–5). The scale is logarithmic. The numbers next to the species name refer to the ranking. The p 50 estimates for Gentiana verna and Gentianella anisodonta when R + C was the criterion were <1 (see the black arrow).
There were 16 species for which the common line model could be accepted for both criteria (Table 1). Conversely, in species where the common slope model or the common intercept model was applicable for both germination criteria, the extent of OESL by using R rather than R + C varied among species, both in terms of percentage and the number of days (Tables 3–5). For example, the OESL50 (days) varied widely, ranging from 0.54 d for Oxyria digyna to 9.01 d for Silene rupestris. Similarly, the OESL50 (%) exhibited substantial variation, ranging from 12.51% for Hieracium pilosella to 4196% for Gentiana verna. Overall, of the 16 seed lots for which estimates of OESL50 (%) were determined, 13 (81%) had OESL50 values between 10 and 90% with the remaining three above this range (Tables 3–5).
Table 4. Estimates (±SE) of seed longevity parameters, Ki and σ (equation 1), the time for viability to fall to 75 and 50% (p 75 and p 50) from laboratory seed storage experiments, and the overestimation of p 75 and p 50 (OESL) when the germination criterion used was radicle emergence (R) rather than radicle plus cotyledon emergence (R + C) (equations 2 and 3)

Seeds were stored at 60% RH and 45°C with samples taken at regular intervals for a germination test. The parameters shown are the results of fitting a model with the same estimate for Ki (‘common intercept’) when the germination criterion was radicle (R) or radicle plus cotyledon emergence (R + C) (Supplementary Figs S2–S5).
Table 5. Estimates of seed longevity parameters for Saxifraga stellaris (Saxifragaceae)

Estimates (±SE) were made of Ki and σ (equation 1), the time for viability to fall to 75 and 50% (p 75 and p 50) from laboratory seed storage experiments, and the overestimation of p 75 and p 50 (OESL) when the germination criterion used was radicle emergence (R) rather than radicle plus cotyledon emergence (R + C) (equations 2 and 3). Seeds were stored at 60% RH and 45°C with samples taken at regular intervals for a germination test. For this species, it was not possible to constrain either Ki or σ without a significant increase in residual deviance (Supplementary Fig. S4).
Interspecies variation in seed longevity
When R + C was used as the germination criterion, p 50 ranged between −0.25 d (i.e. initial fitted viability was less than 50%) for Gentianella anisodonta and 33.98 d for Silene vulgaris subsp. glareosa (Tables 2–5, though both species are reported in Table 3; Fig. 2). There were some taxonomic trends in longevity, for example, the four Silene taxa all had p 50 estimates >20 d; other species with p 50 estimates >10 d belonged to the Asteraceae. The relative rankings in longevity were slightly changed when the measure of seed longevity was p 75 rather than p 50 (Supplementary Fig. S7). For example, Primula farinosa was ranked 8th and 21st according to p 50(R) and p 50(R + C) (Fig. 2) but 10th and 27th according to p 75(R) and p 75(R + C) (Supplementary Fig. S7).
Discussion
In this study, we used both radicle emergence (R) and subsequent cotyledon emergence (R + C) as criteria for assessing germination in 33 species after different experimental storage periods at 45°C and 60% RH, to understand their relative seed longevity. Since radicle emergence normally occurs before cotyledons appear, the position of the survival curves for R + C are shifted to the left (i.e. to shorter storage periods) than those for R (Supplementary Figs S1–S6). This shift was negligible for about half the species so that a single survival curve could be fitted to the R and R + C data without a significant increase in residual deviance (Table 2). Consequently, for these species, germination criterion did not affect the prediction of seed longevity, that is, either R or R + C can be used as a proxy of germination in seed longevity studies. However, for the other species, a single line model could not be fitted. Rather, a common slope or a common intercept model could be accepted, depending on the species (Table 1). For one species, neither of these constrained models could be accepted, that is, the accepted model was one with independent estimates for Ki and σ for the two germination criteria. Given these results, the OESL from using R rather than R + C showed considerable variation in terms of days and as a percentage.
Differences in the survival curves depending on germination criteria have long been recognized, for example, by Rao et al. (Reference Rao, Roberts and Ellis1987) for lettuce seeds, with the curve for normal germination [seeds that produced healthy seedlings sensu ISTA (2018)] being shifted to shorter storage periods than the survival curve for total germination (normal plus abnormal seedlings). Furthermore, Ellis and Roberts (Reference Ellis and Roberts1981) described how all seeds go through a ‘defective’ phase, as the last stage before they lose the ability to germinate, and Ellis et al. (Reference Ellis, Td and Roberts1985) provided advice on evaluation of seedlings when testing the viability of seeds stored in a genebank. Nevertheless, most of the recent literature exploring correlates of seed longevity in wild species has relied on R as a measure of seed viability (e.g. Probert et al., Reference Probert, Daws and Hay2009; Mondoni et al., Reference Mondoni, Probert, Rossi, Vegini and Hay2011; Merritt et al., Reference Merritt, Martyn, Ainsley, Young, Seed, Thorpe, Hay, Commander, Shackelford, Offord and Dixon2014; Bernareggi et al., Reference Bernareggi, Carbognani, Petraglia and Mondoni2015; Seglias, Reference Seglias2022; Sommerville et al., Reference Sommerville, Newby, Yenson and Offord2023; White et al., Reference White, Hay, Abeli and Mondoni2023), indicating a possible OESL for some species. Adoption of R is easier when dealing with wild species since it is often unknown what would be considered abnormal, that is, a seedling that is not able to develop into a mature, reproductive plant. Nonetheless, this bias in the estimation of longevity could mistakenly target species for more frequent viability monitoring and vice versa and alter the perception of successful ex situ conservation of wild species if the ability to regenerate plants from seeds is not better assessed. This concern seems particularly relevant given the growing role of seed banks and other collections/stocks in the provision of propagation material for habitat restoration and species recovery (White et al., Reference White, Hay, Abeli and Mondoni2023). For example, Mondoni et al. (Reference Mondoni, Probert, Rossi, Vegini and Hay2011) reported that seeds of alpine plants have very short, short or medium longevity, based on seed storage experiments with R as the germination criterion. According to our results, seed longevity for seedling growth of several alpine species may be lower than previously reported. For example, two species (Veronica bellidioides and Primula farinose) previously classified as having medium longevity (10 d < p 50 ≤ 100 d; Mondoni et al., Reference Mondoni, Probert, Rossi, Vegini and Hay2011) according to p 50(R) should be reclassified as short-lived (1 d < p 50 ≤10 d) according to p 50(R + C). Similarly, and more alarmingly, seeds of two other species (Gentiana verna and Gentianella anisodonta) classified as having short seed longevity according to p 50(R) were found to be very short-lived (p 50 ≤ 1 d) based on p 50(R + C). Ex situ storage of short-lived seeds such as those of alpine species may, therefore, be more problematic than currently thought.
The range in p 50 estimates observed for these taxa are similar to those observed by Probert et al. (Reference Probert, Daws and Hay2009) and Mondoni et al. (Reference Mondoni, Probert, Rossi, Vegini and Hay2011), for seeds stored under identical conditions with radicle emergence (R) as the germination criterion. Both these previous studies emphasized a correlation between climate and seed longevity, with seeds from species originating from cooler, wetter habitats ranked lower in terms of seed longevity. Our results here, however, again serve to emphasize that there is also variation in seed longevity even when seeds are collected from localities with very similar climates. Thus, more research is clearly needed to further tease out the genetic and environmental controls – and their interaction – on seed longevity.
We introduced a coefficient, denoted as OESL, for species exhibiting the difference of seed longevity based on germination criteria (i.e. R vs R + C). This coefficient provides a deeper understanding of the criticality in assessing the R + C criterion over R. It can be used to prioritize species that require R + C as the germination criterion in seed storage studies. Indeed, even a slight difference in seed longevity between R and R + C is significant for short-lived species in comparison with medium or long-lived seeds. These subtle distinctions do not hold the same significance for long-lived seeds. In general, OESL (%) is more useful than OESL (days). For example, Epilobium fleischeri seeds can be classified as short-lived, with a p 50 of 3.5 d for R and 2.2 d for R + C, while Silene vulgaris subsp. glareosa seeds fall into the medium-lived category, with a longevity of 41.7 d for R and 33.9 d for R + C. Considering OESL in days, the choice of germination criterion has a greater impact for the medium-lived species compared with the short-lived species (OESL of 7.7 and 1.2 d, respectively; Table 3). However, when examining the OESL coefficient (%), a different picture emerges, with short-lived seeds revealing a 54.7% overestimation, while that for the medium-lived seeds being just 22.7%. Hence, this coefficient plays a crucial role in prioritizing species within seed banks, allowing us to determine the most suitable criterion for each species. This approach enables effective screening of species, facilitating the selection of the most appropriate germination criterion, thereby enhancing the overall efficiency and precision of seed storage studies. We have focused on OESL50 since p 50 has become our standard measure of longevity (Walters et al., Reference Walters, Wheeler and Grotenhuis2005; Hay et al., Reference Hay, Valdez, Lee and Sta Cruz2019). If the R and R + C survival curves do not have the same slope, the divergence is greater when viability has fallen to 50% than it is when viability has fallen to just 75% (Tables 4 and 5). Accordingly, there were some changes in the relative rankings of p 75 seed longevity among the taxa considered here, depending on whether the germination criterion was R or R + C (Supplementary Fig. S7). This also needs to be considered if the seed bank recollection standard is 75%.
Finally, the OESL resulting from using R instead of R + C has implications on understanding the ageing process itself. Depending on the germination criteria, for some species the ageing process resulted in the same proportion of abnormal seedlings over the course of viability loss of the seed lot (i.e. common slope model; Table 3), whereas for others, it appears that an increasing proportion of the population of seeds can accumulate damage that is phenotypically expressed before they lose the ability to germinate (R) (i.e. common intercept model; Table 4). Furthermore, the differences between R and R + C during seed ageing may be not only species-specific, but also affected by other factors, such as conservation conditions. Rao et al. (Reference Rao, Roberts and Ellis1987) noted that a common slope model is acceptable in some storage conditions and a common intercept model is acceptable for others, with no apparent trend according to moisture content or temperature, or ‘rate’ of ageing.
Our results illustrate the need to be more discerning when deciding whether a seed is viable or not when conducting seed storage experiments for wild species in general and, perhaps, alpine species in particular. We introduced a coefficient, denoted as OESL (%), which facilitates effective screening, enabling the selection of the most appropriate germination criterion for potential seedling emergence post-storage. Such considerations should enhance the overall conservation efficiency. This might be particularly important for valuable collections of rare and endangered species, especially those with short-lived seeds. Whilst R may continue to be the preferred criterion in germination tests for seed storage experiments for practical reasons, it may be necessary to use a more stringent ‘viability’ standard (the parameter that germination is estimating), than that typically used (e.g. 75%; ISTA, 2018), to ensure that the genetic potential of the desired ≥75% of individuals is realized.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0960258524000175.
Funding statement
This research was funded through the People Programme (Marie Curie Actions) of the European Union's Seventh Framework Programme FP7/2007-2013/ under REA grant agreement no607785 (NASSTEC).










 Open access
Open access