
21 results
Voluntary Animal Welfare Assessment of Mass-Produced Farm Animal Housing Equipment Using a Standardised Procedure
-
- Journal:
- Animal Welfare / Volume 12 / Issue 1 / February 2003
- Published online by Cambridge University Press:
- 11 January 2023, pp. 75-84
-
- Article
- Export citation
YRP.03 Genotyping Reelin in multiple affected families with schizophrenia: a suitable marker for the vulnerability to develop schizophrenia?
-
- Journal:
- European Psychiatry / Volume 17 / Issue S1 / May 2002
- Published online by Cambridge University Press:
- 16 April 2020, p. 220s
-
- Article
-
- You have access
- Export citation
Review: Comparative methane production in mammalian herbivores
-
- Article
-
- You have access
- HTML
- Export citation
ALMA spectrum of the extreme OH/IR star OH 26.5+0.6
-
- Journal:
- Proceedings of the International Astronomical Union / Volume 14 / Issue S343 / August 2018
- Published online by Cambridge University Press:
- 30 December 2019, pp. 436-437
- Print publication:
- August 2018
-
- Article
-
- You have access
- Export citation
Synthesis and structure of magnesium hydroxide fluoride, Mg(OH)F: a topological intermediate between brucite- and rutile-type structures
-
- Journal:
- Mineralogical Magazine / Volume 76 / Issue 1 / February 2012
- Published online by Cambridge University Press:
- 05 July 2018, pp. 25-36
-
- Article
- Export citation
Nomenclature of the micas
-
- Journal:
- Mineralogical Magazine / Volume 63 / Issue 2 / April 1999
- Published online by Cambridge University Press:
- 05 July 2018, pp. 267-279
-
- Article
- Export citation
Evaluation of the ‘Jumping to conclusions’ bias in different subgroups of the at-risk mental state: from cognitive basic symptoms to UHR criteria
-
- Journal:
- Psychological Medicine / Volume 46 / Issue 10 / July 2016
- Published online by Cambridge University Press:
- 20 April 2016, pp. 2071-2081
-
- Article
- Export citation
Dynamic magnetic resonance imaging and spectroscopie of experimental brain injury
-
- Journal:
- Acta Neuropsychiatrica / Volume 8 / Issue 4 / December 1996
- Published online by Cambridge University Press:
- 18 September 2015, pp. 76-86
-
- Article
- Export citation
A Nanocrystalline Hilbert-Phase Plate for Phase-Contrast Transmission Electron Microscopy of Amorphous Objects
-
- Journal:
- Microscopy and Microanalysis / Volume 18 / Issue S2 / July 2012
- Published online by Cambridge University Press:
- 23 November 2012, pp. 496-497
- Print publication:
- July 2012
-
- Article
- Export citation
Comprehensive Comparison of Various Techniques for the Analysis of Elemental Distributions in Thin Films
-
- Journal:
- Microscopy and Microanalysis / Volume 17 / Issue 5 / October 2011
- Published online by Cambridge University Press:
- 12 September 2011, pp. 728-751
- Print publication:
- October 2011
-
- Article
- Export citation
Contributors
-
-
- Book:
- Hormones, Cognition and Dementia
- Published online:
- 06 July 2010
- Print publication:
- 24 September 2009, pp vii-x
-
- Chapter
- Export citation
The Dosage compensation of Drosophila and mammals as showing the accuracy of the normal type*
-
- Journal:
- Genetical Research / Volume 8 / Issue 1 / August 1966
- Published online by Cambridge University Press:
- 14 April 2009, pp. 41-59
-
- Article
-
- You have access
- Export citation
Early results from the SAGE-SMC Spitzer legacy
-
- Journal:
- Proceedings of the International Astronomical Union / Volume 4 / Issue S256 / July 2008
- Published online by Cambridge University Press:
- 01 July 2008, pp. 184-188
- Print publication:
- July 2008
-
- Article
-
- You have access
- Export citation
Vagus nerve stimulation for depression: efficacy and safety in a European study
-
- Journal:
- Psychological Medicine / Volume 38 / Issue 5 / May 2008
- Published online by Cambridge University Press:
- 04 January 2008, pp. 651-661
-
- Article
- Export citation
Defects in SiC substrates and epitaxial layers affecting semiconductor device performance
-
- Journal:
- The European Physical Journal - Applied Physics / Volume 27 / Issue 1-3 / July 2004
- Published online by Cambridge University Press:
- 15 July 2004, pp. 29-35
- Print publication:
- July 2004
-
- Article
- Export citation
Distribution and properties of oxide precipitates in annealed nitrogen doped 300 mm Si wafers
-
- Journal:
- The European Physical Journal - Applied Physics / Volume 27 / Issue 1-3 / July 2004
- Published online by Cambridge University Press:
- 15 July 2004, pp. 159-161
- Print publication:
- July 2004
-
- Article
- Export citation
-
Spatial distribution and properties of oxide were examined in 300 mm nitrogen (N) doped CZ-Si. Experimentally grown materials with N ranging from ~ 1013 cm−3 to 1015 cm−3 were studied by infrared light scattering tomography, scanning infrared microscopy, transmission electron microscopy and electron beam induced current. It was established that an increasing N content improves the uniformity of the radial distribution of precipitates in the bulk of the wafer, the density of precipitates reaching a level of ~ 109 cm−3. The width of the denuded zone varies in the range from $15\;\mu$
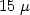 m to $70\;\mu$
m to $70\;\mu$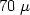 m depending on radial position and
N doping level. Electron microscopy revealed lower oxide precipitate
densities of about 105 to 108 cm−3. The results are
interpreted in terms of existence of agglomerates of nanometer size
precipitate nuclei and/or by the defect-induced strain relaxation
around the precipitates.
m depending on radial position and
N doping level. Electron microscopy revealed lower oxide precipitate
densities of about 105 to 108 cm−3. The results are
interpreted in terms of existence of agglomerates of nanometer size
precipitate nuclei and/or by the defect-induced strain relaxation
around the precipitates.
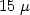
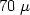
TEM investigations of Ga(Sb,As) quantum dots grown on a seed layer of (In,Ga)As quantum dots
-
- Journal:
- Microscopy and Microanalysis / Volume 9 / Issue S03 / September 2003
- Published online by Cambridge University Press:
- 05 September 2003, pp. 212-213
- Print publication:
- September 2003
-
- Article
- Export citation
¿Son los síntomas afectivos un factor de riesgo para la discinesia tardía en la esquizofrenia?
-
- Journal:
- European Psychiatry (Ed.Española) / Volume 8 / Issue 4 / May 2001
- Published online by Cambridge University Press:
- 12 May 2020, pp. 255-258
- Print publication:
- May 2001
-
- Article
- Export citation
Characterization of the Complex Matrix of the Mytilus Edulis Shell and the Implications for Biomimetic Ceramics
-
- Journal:
- MRS Online Proceedings Library Archive / Volume 255 / 1991
- Published online by Cambridge University Press:
- 21 February 2011, 3
- Print publication:
- 1991
-
- Article
- Export citation
Commission 19: Rotation of the Earth (Rotation De La Terre)
-
- Journal:
- Transactions of the International Astronomical Union / Volume 20 / Issue 1 / 1988
- Published online by Cambridge University Press:
- 25 April 2016, pp. 179-194
- Print publication:
- 1988
-
- Article
-
- You have access
- Export citation
