
87 results
The Oxford handbook of early Christian archaeology. Edited by David K. Pettegrew, William R. Caraher and Thomas W. Davis. Pp. xvi + 707 incl. 189 figs and 2 tables. Oxford–New York: Oxford University Press, 2019. £125. 978 0 19 936904 1
-
- Journal:
- The Journal of Ecclesiastical History / Volume 74 / Issue 3 / July 2023
- Published online by Cambridge University Press:
- 05 July 2023, pp. 645-647
- Print publication:
- July 2023
-
- Article
-
- You have access
- HTML
- Export citation
On solving an isotope dilution model for the partition of phenylalanine and tyrosine uptake by the liver of lactating dairy cows
-
- Journal:
- The Journal of Agricultural Science / Volume 160 / Issue 5 / October 2022
- Published online by Cambridge University Press:
- 08 August 2022, pp. 360-370
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Preeclampsia-induced alterations in brain and liver gene expression and DNA methylation patterns in fetal mice
-
- Journal:
- Journal of Developmental Origins of Health and Disease / Volume 14 / Issue 1 / February 2023
- Published online by Cambridge University Press:
- 24 June 2022, pp. 146-151
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Further solutions to an isotope dilution model for partitioning phenylalanine and tyrosine between milk protein synthesis and other metabolic fates by the mammary gland of lactating dairy cows
-
- Journal:
- The Journal of Agricultural Science / Volume 160 / Issue 3-4 / June 2022
- Published online by Cambridge University Press:
- 11 May 2022, pp. 250-259
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
426 - FindMyApps: Protocol for a randomized controlled trial with community-dwelling people with dementia, evaluating the effectiveness and cost-effectiveness of an intervention to improve self-management and social participation
-
- Journal:
- International Psychogeriatrics / Volume 32 / Issue S1 / October 2020
- Published online by Cambridge University Press:
- 04 November 2020, p. 142
-
- Article
-
- You have access
- Export citation
On cohesive almost zero-dimensional spaces
- Part of
-
- Journal:
- Canadian Mathematical Bulletin / Volume 64 / Issue 2 / June 2021
- Published online by Cambridge University Press:
- 15 July 2020, pp. 429-441
- Print publication:
- June 2021
-
- Article
- Export citation
-
We investigate C-sets in almost zero-dimensional spaces, showing that closed
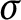 $\sigma $
C-sets are C-sets. As corollaries, we prove that every rim-
$\sigma $
C-sets are C-sets. As corollaries, we prove that every rim-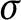 $\sigma $
-compact almost zero-dimensional space is zero-dimensional and that each cohesive almost zero-dimensional space is nowhere rational. To show that these results are sharp, we construct a rim-discrete connected set with an explosion point. We also show that every cohesive almost zero-dimensional subspace of
$\sigma $
-compact almost zero-dimensional space is zero-dimensional and that each cohesive almost zero-dimensional space is nowhere rational. To show that these results are sharp, we construct a rim-discrete connected set with an explosion point. We also show that every cohesive almost zero-dimensional subspace of 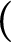 $($
Cantor set
$($
Cantor set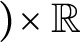 $)\!\times \mathbb R$
is nowhere dense.
$)\!\times \mathbb R$
is nowhere dense.
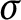
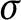
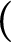
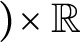
Review: Rumen sensors: data and interpretation for key rumen metabolic processes
-
- Article
-
- You have access
- HTML
- Export citation
Socio-economic differences in the change of fruit and vegetable intakes among Dutch adults between 2004 and 2011: the GLOBE study
-
- Journal:
- Public Health Nutrition / Volume 21 / Issue 9 / June 2018
- Published online by Cambridge University Press:
- 20 February 2018, pp. 1704-1716
-
- Article
-
- You have access
- HTML
- Export citation
Effects of roughage characteristics on enteric methane emission in dairy cows
-
- Journal:
- Advances in Animal Biosciences / Volume 7 / Issue 3 / November 2016
- Published online by Cambridge University Press:
- 28 October 2016, pp. 229-230
- Print publication:
- November 2016
-
- Article
- Export citation
The role of majority and minority language input in the early development of a bilingual vocabulary*
-
- Journal:
- Bilingualism: Language and Cognition / Volume 19 / Issue 1 / January 2016
- Published online by Cambridge University Press:
- 26 February 2015, pp. 191-205
-
- Article
- Export citation
Verbal fluency in Parkinson's disease: results of a 2-min fluency test
-
- Journal:
- Acta Neuropsychiatrica / Volume 18 / Issue 1 / February 2006
- Published online by Cambridge University Press:
- 24 June 2014, pp. 38-41
-
- Article
- Export citation
