
4 results
A new frontier in laboratory physics: magnetized electron–positron plasmas
-
- Journal:
- Journal of Plasma Physics / Volume 86 / Issue 6 / December 2020
- Published online by Cambridge University Press:
- 18 November 2020, 155860601
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Debye length and plasma skin depth: two length scales of interest in the creation and diagnosis of laboratory pair plasmas
- Part of
-
- Journal:
- Journal of Plasma Physics / Volume 83 / Issue 1 / February 2017
- Published online by Cambridge University Press:
- 06 February 2017, 595830106
-
- Article
-
- You have access
- HTML
- Export citation
-
In traditional electron/ion laboratory plasmas, the system size
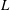 $L$ is much larger than both the plasma skin depth
$L$ is much larger than both the plasma skin depth 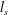 $l_{s}$ and the Debye length
$l_{s}$ and the Debye length 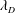 $\unicode[STIX]{x1D706}_{D}$. In current and planned efforts to create electron/positron plasmas in the laboratory, this is not necessarily the case. A low-temperature, low-density system may have
$\unicode[STIX]{x1D706}_{D}$. In current and planned efforts to create electron/positron plasmas in the laboratory, this is not necessarily the case. A low-temperature, low-density system may have  $\unicode[STIX]{x1D706}_{D}<L<l_{s}$; a high-density, thermally relativistic system may have
$\unicode[STIX]{x1D706}_{D}<L<l_{s}$; a high-density, thermally relativistic system may have 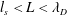 $l_{s}<L<\unicode[STIX]{x1D706}_{D}$. Here we consider the question of what plasma physics phenomena are accessible (and/or diagnostically exploitable) in these different regimes and how this depends on magnetization. While particularly relevant to ongoing pair plasma creation experiments, the transition from single-particle behaviour to collective, ‘plasma’ effects – and how the criterion for that threshold is different for different phenomena – is an important but often neglected topic in electron/ion systems as well.
$l_{s}<L<\unicode[STIX]{x1D706}_{D}$. Here we consider the question of what plasma physics phenomena are accessible (and/or diagnostically exploitable) in these different regimes and how this depends on magnetization. While particularly relevant to ongoing pair plasma creation experiments, the transition from single-particle behaviour to collective, ‘plasma’ effects – and how the criterion for that threshold is different for different phenomena – is an important but often neglected topic in electron/ion systems as well.
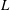
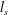
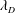

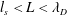
12 - The peopling of Madagascar
-
-
- Book:
- Molecular Biology and Human Diversity
- Published online:
- 19 September 2009
- Print publication:
- 12 September 1996, pp 156-170
-
- Chapter
- Export citation
Mitochondrial DNA variation in eastern highlanders of Papua New Guinea
-
-
- Book:
- Genetic Variation and its Maintenance
- Published online:
- 05 March 2012
- Print publication:
- 30 October 1986, pp 87-100
-
- Chapter
- Export citation
