



Introduction
Faunal remains relate to numerous spheres of practice: diet, technology, environment, landscape exploitation, mobility patterns, social agency, and many more (e.g. Allsen Reference Allsen2006; Bar-Oz Reference Bar-Oz2004; Cartmill Reference Cartmill1993; Munro et al. Reference Munro, Kennerty, Meier, Samei, Al-Nahar and Olszewski2016; Stiner Reference Stiner2002; Stiner & Kuhn Reference Stiner, Kuhn, Hublin and Richards2009; Stiner et al. Reference Stiner, Barkai and Gopher2009). Leafing through hundreds of archaeozoological reports from the southern Levant, it is evident that herbivore mammals comprise the major component of faunal assemblages, with gazelle and fallow deer dominating the prehistoric archaeozoological record and sheep, goats and cattle later periods (Bar-Oz & Weissbrod Reference Bar-Oz, Weissbrod, Bar-Yosef and Enzel2017; Tsahar et al. Reference Tsahar, Izhaki, Lev-Yadun and Bar-Oz2009). From time to time, however, the odd bone of a lion, a bear, or a leopard is reported (e.g. Dayan et al. Reference Dayan, Bar-Yosef, Tchernov and Yom-Tov1986; Kaplan & Ritter-Kaplan Reference Kaplan, Ritter-Kaplan and Stern1993; Wapnish & Hesse Reference Wapnish, Hesse, Finkelstein, Ussishkin and Halpern2000). Until now, such rare carnivoran finds were treated as mere curiosities, because of their sparse representation when viewed at the scale of a single archaeological stratum, site, or even cultural period; single finds are often perceived as unable to support a grounded discussion of their significance. We would like to challenge this view, and argue that if these rare traces seem insignificant, it is because they are observed in too narrow a temporal scale. We demonstrate that, when observed from a broad temporal perspective, clear patterns emerge with significant ideological and symbolic implications.
Our investigation consists of three parts. First, we hypothesize about the anthropological mechanisms that may have been responsible for the occurrence of large carnivoran bones in archaeological sites. Next, based on the review of more than 500 archaeozoological assemblages, we trace the temporal distribution of large carnivoran remains. Finally, we examine our hypotheses about the anthropological mechanisms at work against the temporal distributions of carnivore remains, seeking to produce an integrated account of the processes and mechanisms responsible for the observed patterns.
Setting the stage
Our discussion focuses on lions, leopards and bears. Our choice of these three large carnivores is due to their charismatic presence and visibility in ancient Near Eastern iconography (e.g. Allsen Reference Allsen2006; Beard Reference Beard2020; Russell Reference Russell2012; Russell & Meece Reference Russell, Meece and Hodder2006; Strawn Reference Strawn2005; Türkcan Reference Türkcan2007). Other large, south Levantine carnivores are not considered here due to technical and substantive reasons: wolves are often difficult to distinguish from dogs, hyenas are strongly and negatively associated with scavenger niches (Orbach & Yeshurun Reference Orbach and Yeshurun2019) and cheetahs (Acinonyx jubatus) are too few to warrant an analysis (e.g. Richardson Reference Richardson, Gebel, Kafafi and Rollefson1997).
Anchored in the southern Levant, our study unfolds in the longue durée. It spans the Last Glacial Maximum and the Iron Age, incorporating four major socio-cultural chapters of human history (Table 1). The first chapter encompasses the early and middle Epipaleolithic (c. 25,000–15,000 bp), when mobile hunter-gatherers occupied open-air camps for short durations (Goring-Morris Reference Goring-Morris, Belfer-Cohen, Enzel and Bar-Yosef2017; Maher et al. Reference Maher, Richter and Stock2012). Hunted ungulates and smaller animals were the staples of their animal economies (e.g. Bar-Oz Reference Bar-Oz2004). The second chapter in our sequence encompasses the late Epipaleolithic Natufian culture (Stage 2a: 15,000–11,500 bp), attributed to complex hunter gatherers (Belfer-Cohen & Goring-Morris Reference Belfer-Cohen, Goring-Morris, Bar-Yosef and Valla2013; Maher et al. Reference Maher, Richter and Stock2012) and the Pre-Pottery Neolithic period (Stage 2b: 11,500–8300 bp). Broadly speaking, it embodies the Neolithic Revolution, entailing settling down in larger sedentary communities, plant and animal domestication and a transition from egalitarian to trans-egalitarian modes of social organization (Hayden Reference Hayden and Delage2004; Zeder Reference Zeder2012).
Table 1. The periodical timeframe examined.
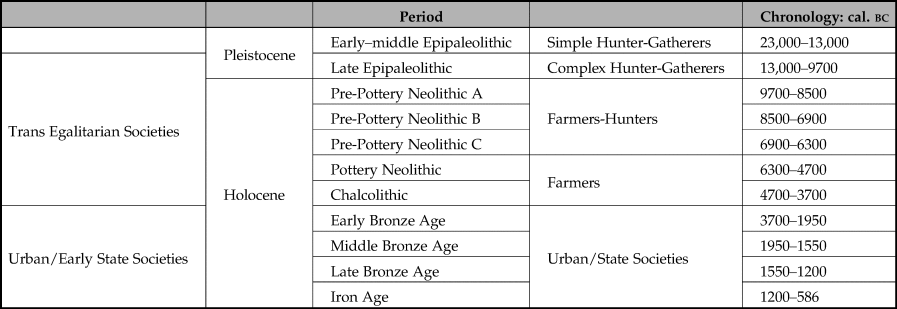
The third chapter spans the subsequent Pottery Neolithic and Chalcolithic periods, embodying the ultimate institutionalization of the agricultural way of life. Among its many features, the institutionalization of agriculture includes complete reliance on crop farming and herd management (Gopher Reference Gopher and Gopher2012), the disappearance of hunting as an economically significant practice (Tsahar et al. Reference Tsahar, Izhaki, Lev-Yadun and Bar-Oz2009), the intensification of long-distance commerce and specialized industries (e.g. metal, stone, ceramic), the proliferation of prestige artefacts, and crystallization of religious institutions (Rowan & Golden Reference Rowan and Golden2009). Although it is widely agreed these developments have considerable implications for the distribution of wealth and power, no definitive evidence was found to date to support claims for the institutionalization of social inequality, comparable to those observed in north Mesopotamia (Marcus Reference Marcus2008). The fourth and last chapter in our sequence spans the Bronze and Iron Ages (3700–586 bc), and represents the formation of urban societies. Temples, palaces, public buildings and writing are among the material manifestations that constituted cities and states. Broadly stated, it was a time preoccupied with the development of managerial and administrative apparatuses (Gilboa Reference Gilboa, Steiner and Killebrew2014; Greenberg Reference Greenberg2019; Joffe Reference Joffe2002).
Given the economic insignificance of carnivores, we suggest three non-exclusive hypotheses to explain the occurrence of the discussed carnivoran finds in archaeological sites throughout the four chapters: contingencies, costly signalling and symbolism.
Contingencies
Our first hypothesis for the occurrence of leopards, bears and lions in the archaeological record states that it is principally the consequence of incidental encounters. Large predators occupy the same or neighbouring ecological niches as humans do. Consequently, humans and large predators are bound to meet; it is a simple matter of probabilities. In this respect, carnivore bones become incorporated in archaeological sites when a chance encounter ends in the animal's death and the collection of its carcass.
Exploring the implications of this mechanism further, we ought to consider two main lines of variation: (1) ecological variability and diversity across space and time and (2) changing nature of human–carnivore relations. Beginning with ecological variability, it is notable that, despite the extensive temporal framework considered, the Levant proved to be remarkably stable, and most species survived well into the Holocene (Tsahar et al. Reference Tsahar, Izhaki, Lev-Yadun and Bar-Oz2009). Nonetheless, encompassing several climatic zones, the southern Levant witnessed shifts of ecological niche boundaries—e.g. between the Mediterranean and arid zones (Palmisano et al. Reference Palmisano, Woodbridge and Roberts2019). However, for our concerns and within our long-term temporal purview, these are but small-scale variations with limited significance for the broader picture. Nonetheless, it urges us to verify that patterns do not coincide with specifiable geographic or climatic zones (i.e. between a Saharo-Arabian desert and Mediterranean mountain terrain), rather compiled into a wide and inclusive frame.
Concerning the second point of human–carnivore encounters, two types of circumstances are notable, pertaining to hunter-gatherers and farmers, respectively. While hunter-gatherers rarely seek confrontation with predators (e.g. Marshall Thomas Reference Marshall Thomas2003, 74, 78; Naveh & Bird-David Reference Naveh and Bird-David2014), such events are expected to occur under competitive circumstances over the same prey. Under these conditions, hunters may end up killing a competing predator while seeking to extract their game (O'Connell et al. Reference O'Connell, Hawkes and Jones1988a,Reference O'Connell, Hawkes and Jonesb), although such events were probably not very common. Among farmers, on the other hand, the confrontation of predators may be induced by the latter's choice to feed on domesticated livestock (Stein et al. Reference Stein, Fuller, Damery, Sievert and Marker2010; Thorn et al. Reference Thorn, Green, Dalerum, Bateman and Scott2012). Here, we may expect more frequent human–carnivore encounters as human settlements expand and economic enterprises reach into larger territories for agriculture and herding. While this holds for all three species of concern here, it is notable that, compared to lions and leopards, bears are less likely to attack herds.
Although bears and lions avoid arid climate zones, these species are considered generalists with respect to physical habitat. Therefore, within our scale of analysis and given herding and hunting exploration zones, spatial variations within the southern Levant are too closely clustered to make a quantitatively significant mark in their distribution. Accordingly, if contingencies were the principal mechanism responsible for the occurrence of large carnivores in the archaeozoological assemblages, we would expect to find a random pattern of taxa throughout the sequence (with a possible decline in bears) and an overall increase in the frequency of carnivore remains, which would reflect a higher rate of encounters correlated with a trend of human population growth.
Costly signalling
Costly signalling speaks for the hunt of large carnivores as a deliberate move intended to acquire social capital. Among hunter-gatherers, hunting and the subsequent distribution of meat can form means to acquire social capital (Hawkes Reference Hawkes1991). The logic behind this social mechanism is often explained using the evolutionary theory of ‘costly signalling’ (e.g. Bliege Bird & Smith Reference Bliege Bird and Smith2005; Hawkes & Biiege Bird Reference Hawkes and Bliege Bird2002; Smith & Bliege Bird Reference Smith and Bliege Bird2000; Stibbard-Hawkes Reference Stibbard-Hawkes2019). Ethnographic and historical research has demonstrated that the same general principles of prestige through hunting also apply to more complex societies (Allsen Reference Allsen2006; Cartmill Reference Cartmill1993; Darimont et al. Reference Darimont, Codding and Hawkes2017; Mihalik et al. Reference Mihalik, Bateman and Darimont2019). The tradeoff is more or less as follows: by putting oneself in harm's way, confronting a fierce and dangerous opponent, the hunter gains social status, influence and power. Lions, bears and leopards—perhaps the most fearsome predators in the region—undoubtedly constitute high-profile targets, bolstering the hunter's image in terms of bravery, strength and skill. Moreover, when these animals are regarded as a threat to the community or its livestock, the successful hunter also emerges as a saviour.
If costly signalling were a major contributing factor to large carnivores’ incorporation in archaeological deposits, we would expect their frequency to increase with social complexity. This is because the ferocity of competition over power, resources and prestige is likely to be correlated with the intensity of ostentatious demonstrations (e.g. Owens & Hayden Reference Owens and Hayden1997; Hayden & Villeneuve Reference Hayden and Villeneuve2011). Following the logic of costly signalling, therefore, we expect to find (1) that substantial engagement with these large predators began with the emergence of complex hunter-gatherer societies, (2) that it increased steadily throughout our temporal sequence, and (3) that it reached its peak as power was being institutionalized and contended during the Bronze and Iron Ages.
Symbolism
Animals figure widely in oral traditions as symbols and embodiments of abstract ideas, signifying anything from gods to human traits (e.g. fox for cunning, chicken for cowardice and peacock for vanity) (Werness Reference Werness2006). In this vein, the third hypothesis suggests that the occurrence of leopards, bears and lions in faunal assemblages be viewed as the archaeologically visible component of an otherwise abstract and symbolic preoccupation with animals. One should bear in mind, however, that an animal's symbolic importance can manifest itself in practice as a taboo over their hunting, as suggested by former research of leopard in Neolithic Anatolia, where their iconography is rich but their skeletal remains are almost absent (Hodder Reference Hodder2006, 261). Thus, if symbolically driven, we may expect that the occurrence of large carnivore remains will correlate—either positively or negatively—to the temporal trajectories charted by other symbolic media, namely the iconographic record. In other words, a positive correlation will be due to a positive view of their hunt (i.e. increase), whereas a negative correlation will imply that a taboo on the hunt was in place. Either way, a pattern should be observed.
Generally speaking, a pattern of this sort is expected to emphasize a shift between leopards and lions. The leopard constituted an important symbol during the Near Eastern Neolithic, evoked in stone, clay, plaster, paint and other media. Some of the most striking examples were recorded in sites like Tell ‘Abr 3 in Syria (Yartah Reference Yartah2005), Göbekli Tepe in southeast Turkey (Peters & Schmidt Reference Peters and Schmidt2004) and Çatalhöyük in central Turkey (Hodder Reference Hodder2006; Mellaart Reference Mellaart1967). In the southern Levant, the site of ‘Uvda 6 is a notable case, consisting of stone constructions in the form of leopards (Avner Reference Avner2002). Lions, on the other hand, are widely evoked during the Bronze and Iron Ages, represented in a range of media that span monumental orthostats and small cups or seals (e.g. Strawn Reference Strawn2005; Zuckerman Reference Zuckerman2008); lion-hunt scenes become a conspicuous iconographic theme that repeatedly occurs in palatial and imperial contexts (Betzig Reference Betzig2008).
As for bears, according to this line of reasoning, we would expect them to be inconspicuous throughout the sequence. While iconographic representations of bears have been documented during the Neolithic and the Bronze and Iron Ages, they are too few and far between (Beard Reference Beard2020; Türkcan Reference Türkcan2007) to indicate the operations of a substantive symbol.
Searching patterns in the archaeozoological data
A total of 516 faunal assemblages were consulted, each assigned to a specific site and archaeological period or culture (database modified from Bar-Oz & Weissbrod Reference Bar-Oz, Weissbrod, Bar-Yosef and Enzel2017; Tsahar et al. Reference Tsahar, Izhaki, Lev-Yadun and Bar-Oz2009). Of these, only 47 contained large predator remains, consisting of 18 leopards, 19 bears and 19 lions, each represented by isolated bones. The temporal and spatial distribution of these species is presented in Figure 1 and Tables 2–5.

Figure 1. Maps of sites bearing lion, bear and leopard remains: (a) late Epipaleolithic and Pre-Pottery Neolithic; (b) Pottery Neolithic and Chalcolithic; (c): Bronze and Iron Ages. Pie diagrams represent percentage of sites bearing bones of lions, bears and leopards.
Table 2. Stage 1, Early–Middle Epipaleolithic representation of leopards (Panthera pardus), bears (Ursus arctos syriacus) and lions (Panthera leo) in the southern Levant.

Table 3. Stage 2, Late Epipaleolithic to Pre-Pottery Neolithic, representation of leopards (Panthera pardus), bears (Ursus arctos syriacus) and lions (Panthera leo) in the southern Levant.

Table 4. Stage 3, Pottery Neolithic to Chalcolithic, representation of leopards (Panthera pardus), bears (Ursus arctos syriacus) and lions (Panthera leo) in the southern Levant.
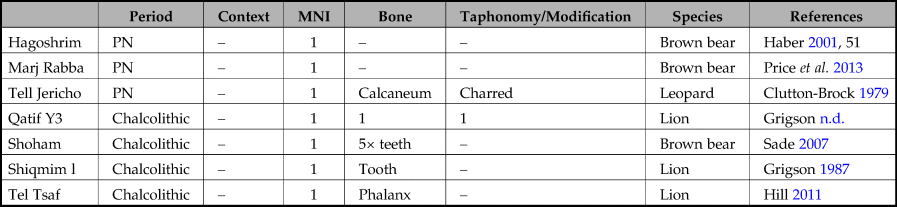
Table 5. Stage 4, Bronze and Iron age representations of leopards (Panthera pardus), bears (Ursus arctos syriacus) and lions (Panthera leo) in the southern Levant.
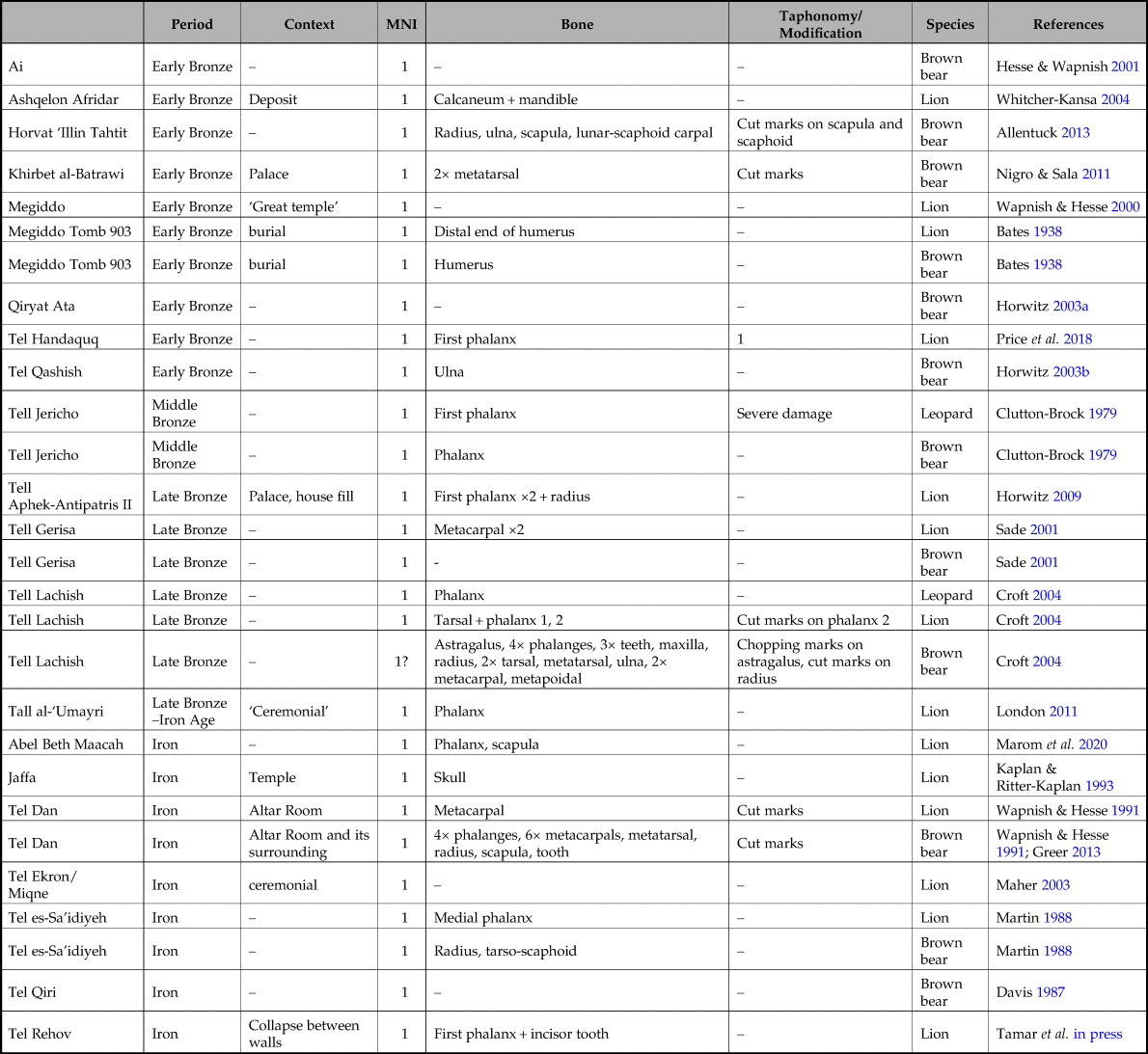
The first noteworthy observation is that large carnivores constitute a tiny fraction of the rich archaeozoological assemblages of the Levant. Importantly, having consulted hundreds of assemblages that derive from a small but intensively researched region, it is difficult to attribute the small numbers to inadequate sampling or inappropriate field practices. Instead, the low frequency of large felids and bears in archaeological deposits is better considered a feature of the phenomenon explored. As such, it underscores and qualifies the exaggerated references to these animals in other media (e.g. Russell Reference Russell2016). First-millennium Mesopotamian literature and iconography are particularly notable in this regard. They portray Assyrian and Babylonian kings killing hundreds of lions, sometimes on foot and sometimes from a chariot (Betzig Reference Betzig2008 and references therein). While scholars have already argued that the numbers cited in such documents ought to be considered symbolically rather than concretely (e.g. Strawn Reference Strawn2005, 162–4), our results suggest that practices of this sort may have been few and far between.
Notwithstanding the small size of our sample, we posit that it is large enough for purposes of statistical analysis and validation. This is in line with a series of methodical assertions on the matter (Motulsky Reference Motulsky2018, 258): most statisticians do not determine a formal lower limit for sample size. When such lower limits are set, they tend to vary. Kraemer and Blasey (Reference Kraemer and Blasey2015), for instance, argued that, for statistical validity, each group ought to comprise at least 10 counts. On the other hand, setting a standard in pharmacology, Curtis and colleagues (Reference Curtis, Bond and Spina2015) determined that a sample consisting of at least five counts per group is sufficient for publication of a statistical analysis. An even lower minimum was set by de Winter (Reference de Winter2013), who used simulations to argue that a t-test can be successfully applied with n = 3 per group. Given these claims, we feel confident that although our samples are small, they are sufficiently large—they exceed 15 counts per taxonomic group—to support statistically valid quantitative analyses (see Lazagabaster et al. Reference Lazagabaster, Ullman, Porat, Halevi, Porat, Davidovich and Marom2021 for a similar argument on archaeological carnivore frequencies).
Carnivore frequencies are clearly associated with the socio-cultural chapters outlined above, and clearly differentiate the mobile hunter-gatherer episode from those that follow. During the 10,000 years of our first chapter, spanning the early and middle Epipaleolithic, only four instances of large carnivores were recorded (Table 2). On the other hand, over the nearly 12,500 years of the rest of our sequence, 52 instances were recorded, manifesting a substantial increase in frequency (Tables 3–6).
Table 6. Contingency table of the occurrence of taxa across stages 2–4. Data are presence/absence.
Two points are of further interest here. The first is that the turning point coincides with the emergence of the complex Natufian hunter-gatherer societies in the late Epipaleolithic (Belfer-Cohen & Goring-Morris Reference Belfer-Cohen, Goring-Morris, Bar-Yosef and Valla2013; Hayden Reference Hayden and Delage2004). The second point is that the rise in large carnivore occurrences transpires while the frequency of hunted animals declines (Fig. 2), best reflected also in that the majority of the cases in stage 2 are from the Pre-Pottery Neolithic B (Stage 2b). This is a very important observation, because it counters the argument that more excavations and sites in the later part of the sequence could result in an increase in the number of large carnivores through more intensive sampling. This rising carnivore trend peaked in the third and fourth stages, in which the basis to urban communities were laid. At this point, hunting was of marginal economic significance, but, within the declining component of wild game in these assemblages, large carnivores constituted an increasingly substantial portion.
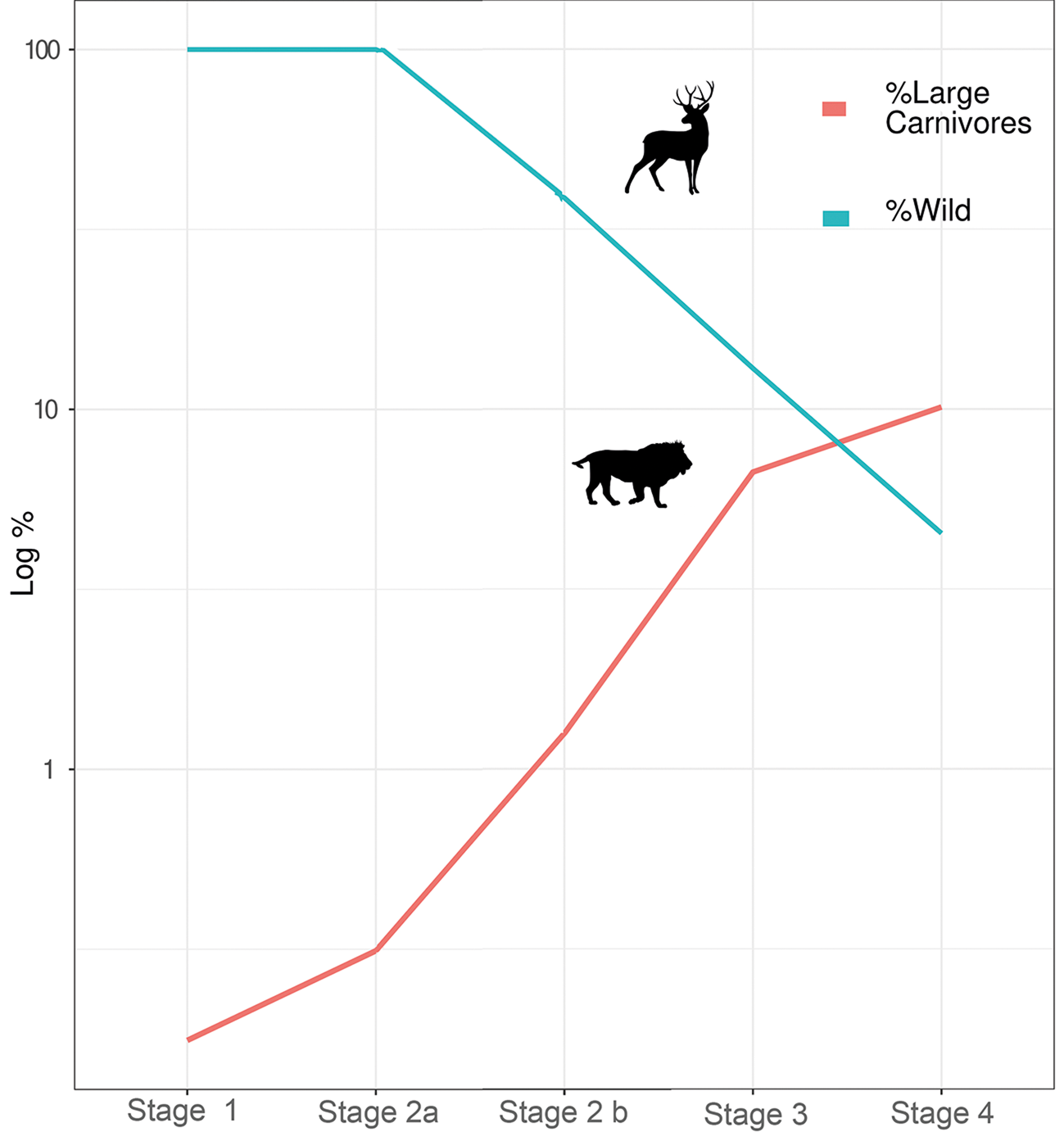
Figure 2. Changes in the relative frequency of large carnivores out of the total number of large game, and of the ratio of hunted (cervids, gazelles, lions, bears and leopards) versus domestic species across the four stages (data retrieved from the reports listed in Tables 2–5). Stage 2 is divided into Natufian (2a) and Pre-Pottery Neolithic (2b). Y-axis is in log(10) scale.
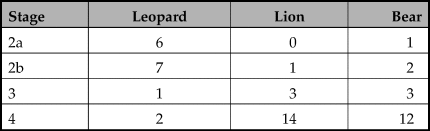
Tracing the relative proportions of the three carnivore species throughout the stages provides the following observations (Fig. 3). Leopards are unequivocally dominant in the second socio-economic stage, accounting for 76.5 per cent (n = 13) of the large carnivores, followed far behind by bears (17.6 per cent, n = 3) and lions (5.9 per cent, n = 1). In the third stage, the pattern is different. Leopard frequencies decline, constituting 14.3 per cent (n = 1) of the large carnivores, while bears and lions come to occupy centre stage with 42.9 per cent (n = 3) each). Finally, the fourth stage seems to continue this trend: the frequency of leopards continues to decline (7.1 per cent, n = 2), while lions and bears become dominant, constituting 50.0 per cent (n = 14) and 42.9 per cent (n = 12) of the carnivoran remains, respectively.

Figure 3. The relative frequency in percentages of lions, leopards and bears throughout the sequence (for details, see Tables 2–5). Stage 2 is divided into Natufian (2a) and Pre-Pottery Neolithic (2b).
Evidently, our second and fourth socio-cultural chapters present us with the most distinctive patterns, suggesting the association of large carnivore species with key episodes of human history. Thus, we find that the emergence of trans-egalitarian societies and the Neolithic Revolution was linked with the leopard, whereas the crystallization of cities and early state societies was associated with lions and bears. Indeed, drawing on contingency Table 6, we found this difference to be statistically significant (Fisher's exact test, p-value = 0.0006), although the sample sizes are below the recommended five counts for each cell of the contingency table. Moreover, plotting chi-square residuals as a measure for a species departure from the null hypothesis produced striking and definitive trends for all three species (Fig. 4): a sharp and consistent decline of leopards, concomitant with a steady increase of lion and bear frequencies. Notwithstanding the small sample size, the inverse correlations between leopard and lion (r = –0.99, P = 0.0008) and leopard and bear (r = –0.99, P = 0.002) are striking.

Figure 4. Change in chi-squared adjusted residuals showing the steady increase of bear and lion numbers and the decrease of leopards in relation to a hypothesis of no association between period and taxonomic frequency.
Having discussed the temporal trends, let us complete our review of the emerging patterns by exploring regional variations in the distribution of carnivore remains. A glance is enough to note some regional patterns: in the arid zones south of Be'er Sheba, one finds only leopards, while the Shephela region (between the southern coastal plain and the mountain terrain) is notable for the high frequency of lions (Fig. 1). This indicates that there is some regional variation in accord with taxa-dependent habitat preferences. However, these patterns are attributable to specific episodes of our sequence—the second and fourth, respectively—and do not seem to suggest a broader trend that transcends our periodic division. In other words, the absence of lions in the Shephela, during our second episode, and the absence of leopards in the arid zones, during the third and fourth episodes, cannot be attributed to ecological factors as lions and leopards survived in these regions into historical (and even modern) times. Therefore, we argue they reflect cultural choices. A complementary perspective can be provided by the Jordan Valley and the northern part of the studied region, where leopards are characteristic of the early stage 2, whereas lions and bears are notable in the later stage 4.
Further indications of purposeful cultural choices emerge from the body parts represented (Tables 2–5; Fig. 5). While leopards are represented by an assortment of postcranial bones, lions and bears show a definitive leaning towards feet and head bones. Given that feet and head bones are characteristic of pelts, the observed difference may indicate that bear and lion skins were valuable, those of leopards less so.

Figure 5. Skeletal element representation by anatomical region and taxon (a). Recalculated to show the percentage of heads and feet only (b) (for details, see Tables 2–5).
The contexts of recovery are also telling. There seems to have been a greater tendency to deposit lion remains in special locations (e.g. caches, temples and palaces: Horwitz Reference Horwitz, Gadot and Yadin2009; Kaplan & Ritter-Kaplan Reference Kaplan, Ritter-Kaplan and Stern1993; Wapnish & Hesse Reference Wapnish and Hesse1991; Whitcher Kansa Reference Whitcher Kansa2004). Seven of the 18 lions recorded in stages 2–4 were retrieved from special deposits of this sort, while only three of the 18 bears and one of 16 leopards derive from similar contexts.
Hypotheses revisited
Above, three tentative hypotheses were offered to explain leopard, bear and lion remains in archaeological assemblages: one spoke for contingencies, another for costly signalling and a third for symbolic discourse. Now that the data and empirical patterns have been outlined, we may revisit them one at a time to estimate their suitability.
Contingencies
According to the contingencies hypothesis, the first expectation is that frequency of the discussed carnivore bones ought to increase steadily with time as people claim more land and resources for themselves and their herds, in the process creating more opportunities for human–carnivore conflicts (Marshall Thomas Reference Marshall Thomas2003; Stein et al. Reference Stein, Fuller, Damery, Sievert and Marker2010; Thorn et al. Reference Thorn, Green, Dalerum, Bateman and Scott2012). This is clearly the case at hand, as reflected in Figure 2.
Nevertheless, the temporally patterned differentiation between leopards, on the one hand, and bears and lions, on the other, speaks against the contingencies hypothesis. It expects to find a more-or-less consistent ratio of these large carnivores throughout the sequence. Perhaps it could tolerate subtle changes due to small-scale environmental shifts, but nothing as dramatic as the observed wholesale shift from one species to another. Also, the possibility that bear relative frequency will decline, following the expectation felids are more likely to attack the growing herds, was not found to be valid. Accordingly, contingencies alone cannot account for the distinct temporal distributions of the species. Some other mechanisms had to intervene between the physical engagement of large predators and the incorporation of physical remains of these animals in archaeological assemblages.
Costly signalling
The hypothesis of costly signalling, as outlined above, suggests the engagement with leopards, bears and lions was a mode of strategic conduct by social actors seeking to boost their standing and influence (e.g. Hayden Reference Hayden1998). Several observations speak in favour of this hypothesis in the context of south Levantine archaeological carnivore assemblages. The first signs of earnest engagement with these large carnivores coincide with the emergence of the complex hunter-gatherers of the Natufian culture (Belfer-Cohen & Goring-Morris Reference Belfer-Cohen, Goring-Morris, Bar-Yosef and Valla2013; Hayden Reference Hayden and Delage2004). It is a time when social mechanisms designed to maintain an egalitarian ethos and discourage boastful behaviour (Bird-David Reference Bird-David, Widlok and Tadesse2005; Kent Reference Kent1993; Lee Reference Lee1979; Wiessner Reference Wiessner1983; Woodburn Reference Woodburn1982) began to relax, mainly due to the shift to sedentism and attendant implications. Consequently, a way opened for hitherto discouraged practices, including a strategic search for new media to boost status (e.g. Bliege Bird & Smith Reference Bliege Bird and Smith2005; Hawkes & Blege Bird Reference Hawkes and Bliege Bird2002; Smith & Bliege Bird Reference Smith and Bliege Bird2000). Without these levelling mechanisms, successful hunters find themselves in an advantageous position to accumulate prestige and power (Hayden & Villeneuve Reference Hayden and Villeneuve2011; Woodburn Reference Woodburn1982). Within this social climate, it is not far-fetched to consider the pursuit of large carnivores as a form of purposefully orchestrated social extravaganza
Moreover, the need for such an extravaganza is likely to have increased during the Neolithic period, as domesticates acquired an increasingly significant part in the animal economy (e.g. Hayden & Villeneuve Reference Hayden and Villeneuve2011; Russell Reference Russell2012; Twiss Reference Twiss2008). As this process unfolded, the hunt and the hunter were economically marginalized, and the large-sized game no longer carried the social benefits and gratitude it used to. Under these circumstances, a search for mightier or more exotic targets constitutes a logical response (Mihalik et al. Reference Mihalik, Bateman and Darimont2019). Large predators are likely to have been well suited for the purpose, and their pursuit would have heralded the transformation of the hunt and the hunter from subsistence providers to combatants, a trend that reached its fruition in the Bronze and Iron Ages.
This leads us to the apparent correlation between the observed preference for lions during the Bronze and Iron Ages and the growing popularity of lion-hunt scenes with their explicit political and imperial agendas. The latter explicitly use the lion and its defeat to advance and legitimize claims for power (Betzig Reference Betzig2008; Strawn Reference Strawn2005), and it seems likely that the increasing frequency of lion remains at this time would be channelled in the same way. Moreover, the semblance of regulated management and the tendency to deposit them in unique locations reinforces this institutionalized power-driven connection.
Nevertheless, costly signalling seems to fall short in explaining the shift from leopards to bears and lions. In principle, the three predators are interchangeable and make little difference for the social mechanics of self-aggrandisement. While the superiority of hunting lions in this frame might be a concern, there is no reason to relinquish the other two if the goal is to demonstrate one's power. Thus, costly signalling is likely to form part of the answer, but certainly not all of it.
Symbolism
If the costly signalling hypothesis has an instrumental tenor, the appeal to symbolic discourse is concerned with conceptual infrastructures. It suggests that leopard, bear and lion remains in archaeological assemblages be understood as the visible tips of a broader preoccupation with these animals as symbols and metaphors. That such a concern did, in fact, exist in the region at the time is demonstrated by the proclivity to evoke these animals in other media: clay, stone, paint, etc. (Betzig Reference Betzig2008; Hodder Reference Hodder2006; Mellaart Reference Mellaart1967; Peters and Schmidt Reference Peters and Schmidt2004; Russell Reference Russell2012; Russell & Meece Reference Russell, Meece and Hodder2006; Strawn Reference Strawn2005).
There is little in the empirical patterns observed that speaks directly against this hypothesis. However, at least one feature speaks strongly in its favour: the definitive distinction between an early leopard-oriented period and the later lion- and bear-oriented period. This distinction seems to accord with trends observed in other modes of symbolic expression throughout the Near East and implies that these animals’ symbolic significance was not taboo-oriented. It also suggests that the recorded succession be viewed as embodiments of the symbols’ careers, including their emergence, negotiation and displacement. In this capacity, the shift from leopard to lion/bear underscores a profound turn in cultural reasoning, priorities and temperament.
In this respect, the case of the bear is puzzling. While the frequency of bears becomes significant in the fourth phase of our sequence, alongside the lions, they figure poorly in other media of symbolic expression (Beard 2000; Türkcan Reference Türkcan2007). Lions’ and bears’ different contexts of recovery is another point of note. While many of the lions were found in special ritual and palatial contexts, this can only be said for a fraction of the bears. Also, the body parts found may attest to the differences, with lions showing the most striking case of feet and heads that correlate to the use of pelts, thus prolonging the hunt into an ongoing social display.
Discussion: towards an integrated account
Having reviewed the three hypotheses, it is evident that none offers a satisfactory explanation on its own. The hypothesis of contingencies underscores the necessary precondition but is insufficient to explain the shifting patterns between the species; costly signalling puts the finger on a robust social mechanism that may have driven the process forward but is unable to account for the change in media; lastly, symbolism provides a prism through which different meanings might have been attached to the leopard, the bear and the lion but remains agnostic concerning all other matters. Fortunately, because these hypotheses occupy different territories—environmental circumstance, politics and metaphors, respectively—one does not exclude the others. In fact, they are probably complementary. Accordingly, in this section, we will try to weave them together. However, for the sake of simplicity, we will set the contingencies hypothesis aside. We do so not because we consider it irrelevant but because it operates at a level that enables the observed phenomena, yet does not seem to account for their particularities.
Hence, we begin with the intertwinement of symbolism and costly signalling. On the one hand, for costly signalling embedded in large predator hunting to be socially effective, it must rely on an appropriate pre-existing symbolic infrastructure. Thus, if hunting a lion is to be useful to boost one's social status, it must rest on an agreed understanding of what a lion means. Only if the large predator is conceived as an objectification of strength, confidence and power can its hunt be mobilized to assert the strength and power of the hunter. On the other hand, abstract symbols and ideas exist and persist only insofar as they are put to work (Bourdieu Reference Bourdieu1990; Giddens Reference Giddens1984). In other words, a symbol needs to be evoked if it is not to disappear. Accordingly, while costly signalling wields symbols in the service of short-term goals, it also reinforces them. In this manner, acts that draw on the leopard or the lion for political purposes also actively support and fuel these animals’ symbolic efficacy. Moreover, in doing so, such practices and applications also negotiate and transform the implicated meanings, applying them under varying circumstances and with shifting emphases.
Thus, our two hypothesized mechanisms—costly signalling and symbolic discourse—are intertwined. They carry each other along a path of their own making. In this vein, we may consider the species’ changing frequencies and, especially, the shift from the leopard to the lion and bear as an expression of the furrow that the two mechanisms ploughed together, making up a play in three acts. The first act corresponds to our second episode, spanning the late Epipaleolithic and Pre-Pottery Neolithic periods. This episode witnessed a substantial increase in the frequencies of large carnivoran bones in archaeological assemblages, mainly attributed to leopards. The second act spans the Pottery Neolithic and Chalcolithic periods and corresponds to our third episode; we observe a general increase in the frequency of large carnivores along with a definitive displacement of leopards by bears and lions. The final act, attributed to the fourth episode in our sequence, spans the Bronze and Iron Ages and consists of further increase in frequencies of carnivore bones while maintaining the dominance of lions and bears. Equipped with a cursory understanding of the relationship between the two hypotheses and a threefold temporal structure for their mutual course, we may now try to follow them with greater detail. We will do so by setting them against the backdrop of the broad evolutionary trends that transpired at the time.
Our first act, marked by leopards, coincides with the Neolithic Revolution. It is a period in the course of which humans were busy negotiating and redefining their place in the world (Cauvin Reference Cauvin2000; Hodder Reference Hodder1990). They did so first by gravitating away from mobility and towards a sedentary way of life, and later by gradually displacing hunting and gathering with herding and farming (Abbo et al. Reference Abbo, Pinhasi van-Oss, Gopher, Saranga, Ofner and Peleg2014; Gopher Reference Gopher and Gopher2012; Zeder Reference Zeder2012). Three features seem particularly pertinent to our discussion. The first is that, in the process of these negotiations, a line was drawn, distinguishing between humans and the rest of the world. This line that was initially but a tentative boundary of the permanent camp—a place to step out of in order to return to it—was persistently reified as more swathes of daily life were domesticated and brought under human control and management. As this human domain continued to evolve and distinguish itself through work and planning, so the world beyond it was alienated, culminating with Nature that is opposed and external to Culture (Hodder Reference Hodder1990).
The second feature concerns the erosion and subsequent undermining of mechanisms that maintain egalitarian relations among individuals. These mechanisms were part and parcel of a mobile hunter-gatherer way of life (Bird-David Reference Bird-David, Widlok and Tadesse2005; Kent Reference Kent1993; Lee Reference Lee1979; Woodburn Reference Woodburn1982). As people moved towards sedentism and food production, these mechanisms began to falter, incrementally making way for differential distributions of influence and power (Hayden Reference Hayden1998). As suggested above, the hunter was probably one of the early beneficiaries of this condition, allowing a successful hunt to be translated better into prestige and social capital (Bliege Bird & Smith Reference Bliege Bird and Smith2005; Hawkes & Bliege Bird Reference Hawkes and Bliege Bird2002; Smith & Bliege Bird Reference Smith and Bliege Bird2000; Stibbard-Hawkes Reference Stibbard-Hawkes2019). This brings us to our third feature of concern: the incremental but consistent marginalization of the hunter. As the subsistence economy drew more heavily towards agriculture and the management of domesticated livestock (Abbo et al. Reference Abbo, Pinhasi van-Oss, Gopher, Saranga, Ofner and Peleg2014; Gopher Reference Gopher and Gopher2012; Zeder Reference Zeder2012), the hunter's economic significance waned and, with it, his/her social esteem.
How does the leopard fit in here? We would like to suggest, in correlation to former research (Hodder Reference Hodder1990), that it crystallized into a symbol of an increasingly mysterious and dangerous world located beyond the confines of the human circle. As the natural environment becomes demarcated and pushed away, the various elements that constitute it inevitably become less familiar, strange and potentially dangerous, encroaching on the carefully structured world people have contrived for themselves. The leopard seems particularly well suited to embody these fears (Hodder Reference Hodder2006). It is a dangerous predator; unlike humans, it is solitary and roams the landscape alone; it is stealthy and covert, out of sight but lurking nearby. Accordingly, the unplanned face-to-face encounter of leopards is likely to have been extremely rare (e.g. O'Connell et al. Reference O'Connell, Hawkes and Jones1988a,Reference O'Connell, Hawkes and Jonesb). Otherwise, a leopard's presence becomes known indirectly via tracks, scat and consumed animal carcasses, rendering its elusiveness a prime feature. In this capacity, it was a symbol drawn upon to articulate and negotiate the distinction between the human and non-human, embodying an ambivalence of fear and yearning, rejection and admiration, which permeated both sides of this still-crystallizing distinction. Thus, the capturing of a leopard, or leopard remains, and bringing it into the cultural realm may have functioned as a device of symbolic negotiation. It offered an opportunity and a channel to conceptualize, apprehend and process something—the non-human realm—that is becoming increasingly ambiguous and threatening. In this respect, the person who hunts the leopard or retrieves its remains is likely to have enjoyed the community's esteem and reverence. He or she brought from the precarious unknown that which the group needed to comprehend and manage their changing place in the world.
It is not improbable that for some, the priorities would have reversed and that the capture of a leopard or its remains acquired the qualities of strategic socio-political practice. Indeed, the hunt of large felids as a means of prestige accumulation is a familiar theme in anthropological and historical research (Buxton Reference Buxton1968; Evans-Pritchard Reference Evans-Pritchard1956; Hazzah et al. Reference Hazzah, Borgerhoff Mulder and Frank2009; Pickenpaugh Reference Pickenpaugh1997). However, a movement in this direction is likely to have intensified with the increasing investment in pastoralism and herd management (Arbuckle & Hammer Reference Arbuckle and Hammer2019). As these practices became widespread, leopards would become more conspicuous and economically menacing as they preyed on livestock (Stein et al. Reference Stein, Fuller, Damery, Sievert and Marker2010; Thorn et al. Reference Thorn, Green, Dalerum, Bateman and Scott2012). Under these circumstances, two crucial developments would have merged: (a) the diminishing social standing of the hunter and (b) a ‘demystification’ of the leopard. In this vein, hunters may have sought to curb the attrition of their status and assert their economic relevance by hunting leopards (e.g. Mihalik et al. Reference Mihalik, Bateman and Darimont2019; Owens & Hayden Reference Owens and Hayden1997). Hence, the increasing frequency of leopard bones during the Pre-Pottery Neolithic.
These trends would have ultimately culminated in the second act of our sequence. It spans the Pottery Neolithic and Chalcolithic periods that mark the closure of the Neolithic Revolution: hunting (and gathering) was now an occupation of negligible economic significance, and the agricultural village with its attendant reliance on domesticated species became the new orthodoxy (Gopher Reference Gopher and Gopher2012); the new economic structures continued to spin in novel ways as domesticated species divulged unexpected benefits in the form of secondary products (Sherratt Reference Sherratt1983); otherwise rare materials—metals, ivory, basalt—begin circulating in greater intensity and over larger distances; and religious institutions begin to crystallize (Ein Gedi, Ghassul, Gilat: Rowan & Golden Reference Rowan and Golden2009).
Under these circumstances, the symbolic boundary-policing that maintained and negotiated the line that separates the human/cultural realm from the non-human/natural one was no longer valued. This Nature/Culture divide was institutionalized, becoming an inseparable and self-perpetuating feature of the agricultural mode of existence (Descola Reference Descola2013; Hodder Reference Hodder1990). Accordingly, the leopard lost its symbolic significance and, by extension, its capacity to draw political weight. It has been reduced to an economic nuisance. While leopards are likely to have remained as much a hazard as they were before, calling for their hunt (see Avner et al. Reference Avner, Shalmon, Hadas, Porat, Horwitz, Brugal, Gardeisen and Zucker2011; Porat et al. Reference Porat, Avner, Holzer, Shemtov and Horwitz2013), there was no motivation to engage with them further. Hunting of large carnivores, however, did not decline and the endeavour shifted to the larger and more dangerous bears and lions.
The third act of our sequence marks the emergence of a new thesis. Letting go of cosmological questions about humans’ place in the world, the focus shifts to humans’ relations among themselves. Specifically, spanning the Bronze and Iron Ages, it is a time of the institutionalization of power, embodied in temples, palaces, armies, city walls, treasuries and other establishments of this sort (Jennings & Earle Reference Jennings and Earle2016). Among these, the lion is readily appropriated as a symbol of vigour, courage and determination, as numerous murals and other depictions readily demonstrate (Betzig Reference Betzig2008; Strawn Reference Strawn2005). The costly signalling associated with the lion hunt is practically fused with the animal's symbolic significance, converting the lion's physical strength into the king's political authority (Cornelius Reference Cornelius1989; Strawn Reference Strawn2005: 158–60). It is based on a state of mind that is effective to this day: that power begets power, a political variation of sympathetic, almost Frazerian, logic. The king appropriates the lion's power by vanquishing it, thus justifying his right to rule. In this respect, the striking increase in the frequencies of lion remains in the archaeological record of the Bronze and Iron Ages indicates a growing disposition towards the admiration of physical force and an intense effort to justify and consolidate political authority by means of costly signalling.
The place of the bear is ambiguous, however. While in some cases it might have been symbolically equivalent to the lions, as noted in several first-millennium bc texts, its literary and iconographic prominence was nowhere nearly as substantial as the lion's (Beard Reference Beard2020). In other words, while the shift from leopards to lions is also observable in the iconographic record, the case of the bear is different. Perhaps this is a matter of medium and bears figured more prominently in verbal discourse. Maybe also relevant here is the observation that lion remains are more closely associated with ritual and palatial contexts than bears. This possibly implies that, while both were mobilized by costly signalling, they operated in different social spheres and may have carried distinct symbolic and ideological charges.
This leads us to consider how the representation of the different body parts of the large carnivores can contribute to our understanding of the dynamics between their role in costly signalling and ideological manifestations. Costly signalling displayed by the hunt is a short-lived event, even if the carcass is brought back to the site. Pelts, on the other hand, constitute a medium to maintain a long-term effect. While leopard pelts are illustrated on the walls of Çatalhöyük (Mellaart Reference Mellaart1967), our results (Fig. 5) suggest that this practice seems to have been more pertinent for lions. In this respect, the use of pelts, which may have emerged as an effort to prolong the impact of the successful hunt, would have shifted away from costly signalling as the honest signal (e.g. Bliege Bird & Smith Reference Bliege Bird and Smith2005) of performing the hunt by themselves become obscured and possibility also irrelevant. By doing so, however, it entered more clearly into the symbolic and ideological discourse.
Thus, to summarize our integrated narrative, in the beginning, we find a concern with the world; in the end, we find politics. Whereas the ‘Neolithic leopard’ was symbolically cosmological, the ‘stately lion’ was inherently authoritarian. Whereas political benefits drawn from the leopard were necessarily circuitous and probably meagre, the lion was a symbol designed to rake in political advantages.
Supplementary material
Supplementary material may be found at https://doi.org/10.1017/S0959774322000221
Acknowledgements
Graphics were made by Sapir Haad. We wish to thank Isaak Gilead for data of Qatif Y and we also wish to thank the two reviewers for their constructive comments. This research was partially funded by the DEADSEA_ECO project (ERC-Stg #802752).













 Open access
Open access