


Are We a Privileged Generation?
For many millennia, humans have gazed up in wonder at the night-time sky. The full panoply of the Milky Way is an awesome sight. The scale of space is immense. Is there life out there somewhere? If so, where, and what form does it take? In the space of a couple of sentences, we’ve already gone from generalized wonder to specific questions. The next step is from questions to hypotheses, or, in other words, proposed answers. Here are two such hypotheses that I’ll flesh out as the book progresses: first, life exists on trillions of planets in the universe; second, it usually follows evolutionary pathways that are broadly similar to – though different in detail from – those taken on Earth.
Going from cosmic wonder to questions about alien life and on to hypothetical answers has been possible for a long time. But the final steps – from hypothetical answers to testing the hypotheses to arriving at an understanding of reality – have so far proved beyond humanity’s grasp. That may be about to change. We may be on the threshold of being able to answer the age-old question ‘are we alone in the universe?’ with a resounding ‘no’. But what is the basis of this claim? The short answer is advancing technology, and in particular advancing design of telescopes. We’ll get to that topic soon. But first, let me reassure you that I’m not alone in my optimism about the imminent discovery of evidence for alien life.
A 2021 paper in the leading scientific journal Nature, by the American physicist James Green and his colleagues, began with the following sentence: ‘Our generation could realistically be the one to discover evidence of life beyond Earth.’ Its authors clearly share my optimism in this respect. Of course, there’s a question-mark hanging over the meaning of ‘our generation’, as the six authors are varied in age, and their readers even more so. But that’s a small uncertainty in the grand scheme of things. The key point is that many scientists suspect that the first evidence of life beyond Earth will be forthcoming on a timescale of years to decades, not centuries. Even if it takes five decades to acquire the first conclusive evidence, a reasonable proportion of those alive today (regrettably not including me!) will still be here to see it.
In the previous paragraph, I used the phrase ‘many scientists’. But scientists of what type? Science is a broad endeavour, and some parts of it are more relevant to the search for life than others. However, that said, many scientific disciplines have a role to play. They include cosmology, astronomy, astrophysics, planetary science, atmospheric science, geology, ecology, evolutionary biology, genetics, molecular biology, and organic chemistry – and that is far from being an exhaustive list. The principal discipline is astrobiology, which is a mix of some or all of the above, with the exact mix depending on the astrobiologist concerned. Astrobiologists of my generation weren’t trained as such, as there were no courses in the subject, so we’ve migrated towards it from various starting points – in my case evolutionary biology. But there’s a younger cohort of astrobiologists who have indeed been trained in this discipline.
Importantly, the ‘many scientists’ who suspect that the first evidence of alien life will be found on the years-to-decades timescale include specialists in all of the disciplines mentioned above, and indeed others too. So this optimism isn’t a passing fad in one particular narrow branch of science. Rather, it’s a reasoned assessment of the stage we have reached in the process of searching for life, and where we’re likely to get to with the new generation of space telescopes that are currently at various stages of planning, construction, or early operation, including the James Webb Space Telescope (alias JWST or simply ‘Webb’), more on which later.
The paper in Nature that I mentioned above is focused on formulating a policy for reporting possible evidence of alien life. The authors highlight the complementary problems of false positives and false negatives. The former involve concluding that some observation is evidence for life when in fact it’s not; the latter involve failing to detect life on a planet when it’s actually there. They particularly stress their concern about the likely sensationalization in the popular media of results that turn out to be false positives, and I think they are right to do so. Their approach to avoiding – or at least reducing – such problems is to develop a scale of the quality of possible evidence of alien life, and to make clear where on this scale any particular finding falls. This way, a hint of possible life shouldn’t be mistaken for a certainty. In Chapter 5, we’ll apply their approach to claims of extraterrestrial life in the solar system.
In a way, I see this book as being complementary to the paper by James Green and colleagues. Those authors were concerned with understanding the nature of possible evidence for alien life in a rational framework, and preventing distortion of that evidence. I’m concerned with understanding the multidisciplinary basis of the search for life, and preventing flights of fancy that are too narrowly based. For example, a biologist might get excited about the possibility of evolution taking place on a particular planet, when astronomical considerations suggest that’s unlikely – for example because of a short lifespan of its host star. Equally, an astronomer might get excited about evolution taking place very rapidly on some planets via large-effect mutations, when the biological argument against this happening isn’t restricted to evolution on Earth. And some scientists (or, more likely, some non-scientists) might get excited about the possibility of life based on silicon rather than carbon, without being aware of the unlikelihood or impossibility (it’s hard to say which) of having a silicon-based molecule that can rival DNA in terms of informational capacity.
A Brief History of the Search
To look at the history of our search for extraterrestrial life, and its philosophical foundations, ancient Greece is a good place to start. Most of the impressive philosophers there in the period from about 600 to 150 BCE (Before the Common Era, alias BC for Before Christ) included astronomical matters in their thoughts. They wondered very deeply about the things they saw in the night-time sky. Anaximander (c. 610 to 546 BCE) argued for a plurality of worlds rather than a single one. However, his approach was constrained by the limited knowledge available at the time on the nature of what we now know to be stars and planets. His ‘many worlds’ were abstract philosophical entities, not solid rocky bodies orbiting stars. Anaxagoras (c. 500 to 428 BCE) is thought to have been the first person to propose that stars are distant suns – or conversely that our Sun is a close-up star. And Aristarchus of Samos (c. 310 to 230 BCE) may have been the first person to propose a heliocentric solar system, with Earth and the other planets known at the time orbiting the Sun.
Prior to the realizations that (1) stars are suns and (2) planets orbit suns, hypotheses about extraterrestrial life couldn’t be formulated in the way that’s possible today. However, as early as Aristarchus’s time, such hypotheses would have been possible. We don’t know of any hypotheses of this kind from way back then. One of the reasons for the lack of them was the unfortunate failure of Aristarchus to persuade most of his contemporaries of the truth of his heliocentric view. It wasn’t until the sixteenth century that Copernicus (1473–1543) would succeed where Aristarchus had failed, and a heliocentric view became commonplace among astronomers.
Although future historical research may turn up ideas about life being found on planets other than Earth between the times of Aristarchus and Copernicus, the earliest known hypotheses of this kind at the time of writing were those of the post-Copernican Italian priest–astronomer Giordano Bruno (1548–1600). Bruno’s view was that stars were distant suns (following Anaxagoras), that each of these had planets orbiting it in the same way as does our local Sun (a generalization of the views of Aristarchus and Copernicus), and that many of these planets were homes to life. Bruno was a man before his time; the first planets orbiting other stars than the Sun – collectively termed exoplanets – weren’t discovered until the late twentieth century. Unfortunately for humanity, and even more so for him personally, his free-thinking approach to both astronomy and theology fell foul of the Catholic Church’s Roman Inquisition. Bruno was burned at the stake – an unimaginably cruel form of homicide – in 1600, in the Campo de’ Fiori in central Rome, where a statue of him can now be found.
The first recognized telescope was made in the Netherlands in 1608, though there may have been previous prototypes. Galileo famously used a telescope to observe the moons of Jupiter orbiting the giant planet in 1610, setting him off on a course that would also fall foul of the Church, though in his case terminating in house arrest rather than execution. As far as we know, Galileo himself didn’t develop hypotheses about alien life, but as telescopes evolved, other users did. The Italian astronomer Giovanni Schiaparelli (1835–1910) observed linear features on Mars that he called canali – which translates into English either as ‘canals’ (which implies human construction) or ‘channels’ (which doesn’t). Schiaparelli wrote about a possible technological civilization on Mars, as did the American astronomer Percival Lowell (1855–1916), who was much struck by the supposed canals. Lowell published a 1906 book entitled Mars and Its Canals, with a 1908 follow-up, Mars as the Abode of Life. In the end, this whole line of enquiry came to nothing, when it was shown that the ‘canals’ were optical illusions.
The modern era of the search for life can be dated to around 1960, with the start of the endeavour called SETI (Search for Extraterrestrial Intelligence). A pioneering paper on the possibility of communication with intelligent life on other planets was published in 1959 by the astronomers Giuseppe Cocconi and Philip Morrison. Frank Drake and others began practical SETI work with Project Ozma in 1960. This was a search for radio signals from alien civilizations, conducted using the National Radio Astronomy Observatory, now the Green Bank Observatory, in West Virginia. Of course, the search for alien intelligence is only one part of the search for life. Nevertheless, from the 1960s onwards, both astrobiology in general and SETI in particular have evolved side by side in a much more continuous way than did the search for life before the second half of the twentieth century.
The Importance of Telescopes
It’s a long way from Galileo’s tiny telescope of 1610 to today’s large ground-based instruments such as Chile’s VLT (Very Large Telescope), and their space-based counterparts such as Hubble and Webb. I’ll now look at a few selected milestones along this evolutionary journey. If you’d like a bit more detail than I can give in just a few pages on the functioning and/or history of telescopes, I can recommend Geoff Reference CottrellCottrell’s 2016 book Telescopes: A Very Short Introduction.
Before examining milestones in the evolution of telescopes, it’s worth reminding ourselves of why, from an astrobiological perspective, they are collectively important. Telescopes of Galileo’s day were great for astronomy – especially when compared with the naked eye – but they weren’t much use for astrobiology. In contrast, today’s telescopes have the capability of addressing hypotheses about alien life. For example, we can now use telescopes to examine the atmospheres of distant exoplanets. We can search for signs of oxygen, which is regarded as a biosignature. In particular, it may be a signature of photosynthesis by microbes and plants. It’s true that oxygen may come to exist in planetary atmospheres by abiotic means. However, as we’ll see in Chapter 6 (penultimate section), it may be possible to distinguish the two types of origin of oxygen. This is a hugely exciting prospect.
So, what changes in telescope design have contributed to our current ability to look for biosignatures? Here are the main ones. First, increasing size. Telescopes are first and foremost light-gathering devices. The amount of light they can collect depends on their size – and in particular the diameter of their primary lens or mirror. The earliest telescopes were very small. Galileo’s had a diameter of 3.7 centimetres, Newton’s (in 1668) 2.5 centimetres. They’re not directly comparable because Galileo’s was a refractor (a telescope that uses lenses) while Newton’s was a reflector (a telescope that uses mirrors), but they were both tiny by current standards. Today’s large telescopes have mirrors with diameters measured in metres rather than centimetres. For example, the Gran Telescopio Canarias, situated on the Spanish island of La Palma, has an aperture of 10.4 metres.
The increase in telescope size has been continuous from the 1600s to the 2000s, though it has speeded up significantly since 1900. The largest telescope in the world at the end of the nineteenth century was the Leviathan of Parsonstown in County Offaly, Ireland, which had a diameter of 1.83 metres. This size was exceeded in 1917 with the construction of the Hooker Telescope on Mount Wilson in southern California, with a diameter of 2.5 metres. So the last 100 years have seen the largest increase – more than 8 metres between the early twentieth and early twenty-first centuries.
However, these measurements in metres don’t quite suffice to explain just how much more powerful modern telescopes are than their ancient forerunners. The light-collecting capability of a telescope scales not with the diameter of its primary mirror but with its surface area. So, in terms of functionality, a 10-metre mirror is not 10 times as good as a 1-metre mirror; rather it’s better by a factor of 100. Given that Galileo’s tiny telescope allowed him to see the moons of Jupiter, and discern individual stars in the ‘milk’ of the Milky Way, the capabilities of today’s telescopes are truly amazing.
But, for astrobiology, size is not enough. There are several other developments that we need to understand. First, so obvious that it hardly needs to be said, the advent of photography in the 1800s, and its later coming together with telescopes in the form of astrophotography. Second, Earth’s atmosphere is a major cause of distortion in the business of seeing clearly into space. As is well known, stars don’t twinkle – they only seem to do so because of atmospheric effects. The main exoplanet-detecting telescopes have been placed above the atmosphere so that this distortion is avoided. These are the Kepler and TESS space telescopes (more on which in Chapter 6). The first is named after the German astronomer Johannes Kepler (1571–1630), who formulated the laws of planetary motion; the second is an acronym of Transiting Exoplanets Survey Satellite, with ‘transiting’ referring to the movement of a planet across the face of its host star, as seen from our perspective.
As well as making a telescope bigger, enabling it to take images, and in some cases launching it into space, another development we need to consider is the range of wavelengths it can see. ‘Light’ is a slippery term. It’s sometimes used for the part of the electromagnetic spectrum we can see, though this is better described as ‘visible light’. Alternatively, it can be used more broadly to refer to the whole of the spectrum – as in the distinction between the speeds of light waves and sound waves. Early telescopes were designed – understandably – to see visible light. Today’s telescopes are much more varied in this respect. Some can see in the ultraviolet range, some in the infrared range, some can see radio waves, and some can see across boundaries between our quasi-arbitrary sections of the spectrum.
One example of ‘seeing across boundaries’ is the James Webb Space Telescope, launched in December 2021. Another will be a future space telescope, which, at the concept stage, was called LUVOIR, an acronym of Large Ultra-Violet, Optical, and Infra-Red (telescope). This design survived NASA’s 2020 decadal survey, which produced its report in late 2021 due to the Covid pandemic. However, it was scaled down in size for reasons of cost. The primary mirror will be of a smaller diameter than was originally envisaged. When constructed, the diameter will be about 6 metres – still a considerable size for a space telescope. Its final name will probably be different. In the immediate aftermath of the decadal survey, some astronomers jokingly suggested simply losing the L (large) and calling it UVOIR, but a better name will almost certainly be found. The ability to see across the boundary between visible and infrared light that characterizes both James Webb and LUVOIR means that they will be ideally placed to see oxygen biosignatures.
This point brings us to the other important development in telescopes that we need to examine: the use of a technique called spectroscopy. Here’s the basic idea behind this technique, as applied in space telescopes for astrobiological purposes. You look at light from a distant source – say a star that has an orbiting planet. In fact, you look at it twice, once when the planet is behind the star or a long way off to one side, and once when it is transiting – i.e. in front of its star from our perspective. You compare graphs of amount of light received versus wavelength for the two occasions (Figure 1.1). Dips in the amount of light at particular wavelengths when the planet is in front of the star are interpreted as resulting from absorption of light by gases in the planet’s atmosphere. Since different gases have different characteristic patterns of absorption, you can identify them.
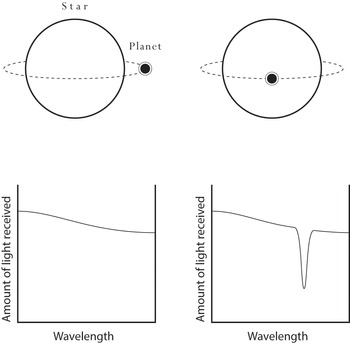
Figure 1.1 Detecting gases in exoplanet atmospheres. Left: amount of light received by a space telescope when the planet is ‘off to one side’ from our perspective. Right: amount of light received when the planet is in a direct line between its host star and the Earth. Note the dip in the amount of light received at a particular wavelength in the right-hand panel. If instead of looking at this graph you were looking at the light itself, after it had been split into its various colours by a prism, you would see a dark band at the wavelength (= colour) concerned. Such a dip (or band) is interpreted as the result of absorption by a particular gas in the planet’s atmosphere, and is known as an absorption band. Since different gases have different patterns of absorption centred on different wavelengths – in other words different ‘signatures’ – they can be identified. Some gases, notably oxygen, are regarded as biosignatures. As we will see later (in Chapter 6), the idea of a single absorption band is a simplification; each gas typically has several of them.
There’s a final twist to this story. The effects of a planetary atmosphere are tiny compared with the immense blast of light that hits our telescope from the fiery furnace of the planet’s host star. A better approach would seem to be this: look not at the star with the planet in front of it but rather at the planet itself. There’s a big problem here that at first seems insurmountable but is not. Even when the planet is a long way off to the side of the star, starlight dominates what you see, and in any event planets don’t produce light of their own. The solution to this problem is twofold. First, where possible, blot out the light from the star using some form of starshade. Second, observe in the infrared range because, while planets don’t emit their own visible light, they do emit in the infrared; in contrast, starlight peaks in the visible region and declines in the infrared. Looking directly at planets in this way is called direct imaging.
Seeing (or ‘listening’) in the radio range is also important – especially from a SETI perspective. As well as searching for biosignatures that could come from very simple forms of life such as bacteria, we also look for technosignatures – those that could only come from an advanced civilization. If such a civilization decides to broadcast its existence in the way that humans have done since the middle of the twentieth century, it will probably use radio signals to do so. Thus, radio telescopes come into their own in the search for intelligence in the universe. Well-known radio telescopes include Jodrell Bank in Cheshire, England, and the Arecibo telescope in Puerto Rico, the latter now lamentably defunct following damage sustained in 2020. It gave its name to the famous Arecibo message broadcast by it into space in 1974. There are also large arrays of radio telescopes – for example ASKAP (Australian Square Kilometre Array Pathfinder), located about 700 kilometres north of the city of Perth, which started observing the sky in 2012.
A Working Definition of Life
We are about to feel the multidisciplinary nature of the search for extraterrestrial life that I mentioned at the outset, because we are about to move from the astronomical realm to its biological counterpart. In particular, we are going to move from the telescopes used in the search for habitable planets – and ultimately the life that’s on some of them – to the search for a workable definition of life. How can we search for life if we can’t define it? And yet the task of defining life is not an easy one – even in the restricted context of planet Earth (are viruses alive?). It’s more difficult again to attempt a definition that will work in a cosmic context. But an imperfect definition is better than nothing – so here goes.
One way of looking at a definition is as an abstract generalization about a class of entities that’s based on prior knowledge of many concrete individual entities of the sort concerned. So, let’s start with a list of ‘concrete’ living entities – organisms – on our home planet. Not a complete list, of course, for that would include more than a million names. But we don’t need a complete list, nor even a particularly lengthy one. A short one will suffice, and indeed is preferable, in terms of not drowning in detail. Here it is: humans, snails, whales, trees, toadstools, and bacteria.
A definition of ‘X’ – in this case ‘life’ – only works if we also have entities that we know are ‘not X’, which in this case means ‘not life’, or alternatively ‘inert objects’. Here’s a short list of such objects, all of which are generally agreed not to be living things: rocks, clouds, cars, smoke, and water. Of course, some of these entities might contain life forms – for example a cloud might contain airborne microbes – but that’s not the same thing as a cloud actually being a living organism itself. And in one case – cars – they’re made by life forms, but again that’s something different.
So, the question is this: how do we word an abstract definition of life so that it includes all of the things that we think of as living and none of those that we think are not? There are different ways of doing this, and I have no intention of surveying them all. That would be both boring and pointless. Instead, I will focus on the definition that I think works best. I call it the RIM definition of life. These letters stand for the following three attributes of life forms: Reproduction, Inheritance, Metabolism. So, an entity that has all three RIM attributes is alive, while an entity with none of them is not. We’ll get to the annoying grey area in the middle shortly.
Now is the moment for a warning about the risk of infinite regress. To define life, it’s necessary to define reproduction, inheritance, and metabolism. This could lead to a need to define words that occur in these definitions, and so on indefinitely. You’ll be pleased to know that I’m absolutely not going to go down that road. However, I am going to briefly consider each of the three components of RIM. Since these three things are crucial in allowing us to distinguish life forms from inert entities – potentially throughout the universe – it simply won’t work if we’re fuzzy about their meanings.
This point was brought home to me when I was reading the Very Short Introduction to astrobiology written by the planetary scientist David C. Catling in 2013. With regard to metabolism, Catling says the following: ‘All life metabolizes but so does my car.’ I don’t agree with him, but I see what he means. The burning of a fuel such as petrol powers a car in much the same way as the burning of a fuel such as glucose powers a living organism. So Catling is right to see a parallel. But he’s wrong – in my view – to equate automotive and biological ‘metabolism’.
I have an instinctive feeling that the two are different in some fundamental way, and perhaps you do too. But what is it? Of course, the fuels are different, but that’s not the best approach to this issue, because we can’t guarantee that the fuel of alien life is glucose. Here’s a better approach. A life form metabolizes right from the start; a car doesn’t. An organism is not simply a collection of cells, membranes, vesicles, filaments, and so on, that is inert until some point in its life when it gets fired up with an infusion of glucose or some other fuel. In contrast, a car is an object of precisely this latter sort. It’s a collection of metal and plastic parts that gets built in stages and hasn’t a whiff of ‘metabolism’ until after it’s complete – or has reached its ‘adult’ stage, to stretch the use of that term – and someone turns on the ignition.
So, to be a life form from a metabolic perspective you need to be metabolizing right from the start. Even the very first cell of a new human or other animal – the zygote – metabolizes. An organism is an intrinsically metabolizing thing. Its default state is to metabolize. However, to call life forms perpetually metabolizing things would be a step too far. We’re all familiar with hibernation – in hedgehogs, for example. This is a state of reduced metabolic activity in cold weather. And there’s the related phenomenon of aestivation – reduced metabolic activity in hot dry weather, as is found in some Mediterranean snails. But these two states are eclipsed by a more extreme one called cryptobiosis (‘hidden life’), in which metabolism may be reduced to zero, or very nearly so.
The best example of animals exhibiting cryptobiosis are the tiny invertebrates called tardigrades, or water bears. These have played an interesting role in astrobiology, as we’ll see later. So it’s appropriate that they should feature in our discussion here of the definition of life. When environmental conditions become too extreme for a tardigrade to maintain normal, active, rapidly metabolizing life, it converts itself into a static, low-or-zero metabolizing structure called a tun. Both this state and the normal active state are shown in Figure 1.2. When conditions improve again, it can convert back from a tun into an active, mobile, metabolizing tardigrade. But this fact doesn’t really cause us difficulties of definition. As long as we stick with a view of organisms as ‘intrinsically metabolizing’ rather than ‘perpetually metabolizing’, then we’re ok.
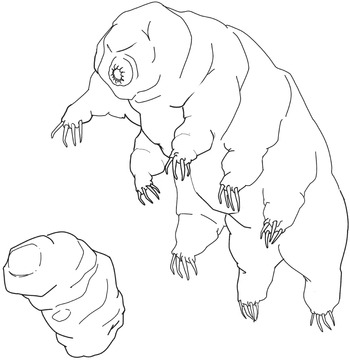
Figure 1.2 Two forms of the tiny invertebrate called a tardigrade or water bear. Upper right: how the animal looks under benign environmental conditions, when it is active. Lower left: a tardigrade tun, which is a resistant, non-metabolizing form that the animal converts itself into under extreme environmental conditions such as very low temperatures or lack of water. If conditions become less extreme, the tun can revert to the active form.
That was a quick look at ‘metabolism’. Now, what about reproduction? Cars don’t reproduce. Neither do rocks or clouds of smoke. Organisms do. Perhaps this is more straightforward, then, than the case of metabolism? Well, not entirely. Regrettably, not all adult humans can reproduce. But those who can’t are no less alive than those who can. In bees and ants, there are whole castes that can’t reproduce – workers, for example. These are no less alive than reproductive drones and queens. Also, cases are known where two species occasionally interbreed, producing offspring that are typically sterile – such as the mule, a hybrid between a horse and a donkey. So, we have to use reproduction as a characteristic of a species rather than of every individual, in our attempts to draw a line between life and non-life.
Now we come to the third attribute included in the RIM definition of life – inheritance. This applies to all life forms on Earth, though in different ways to some than to others. We humans are most familiar with the sort of inheritance that is associated with sexual reproduction, in which, as we can plainly see, offspring tend to resemble one or both parents more than they do other members of the overall population. Most other animal species have sexual reproduction, so the same phenomenon applies, even though, for example, we humans find it hard to distinguish degrees of similarity among different individual houseflies.
Inheritance still applies in the world of asexual reproduction, which is much more common in bacteria and other microbes than it is in animals. A population of a particular kind of bacterium that reproduces in this way can be thought of us a collection of clones. Each parent, on its own, produces progeny that are genetically identical to itself, so the within-clone variation is zero at the level of the gene (unless a mutation occurs), and may be quite small at the level of the organism. In contrast, the between-clone variation is greater. Since clones are to all intents and purposes families, the situation regarding inheritance is broadly the same. Finally, we must be equally careful in deciding what entity to apply inheritance to, as we were in using the term reproduction. Inheritance applies to the relationship between generations, not to individual organisms taken alone. Of the three RIM attributes, metabolism is the only one that can be applied to individuals considered in isolation from other members of their family, population, or species.
In summary, the RIM definition of life can be constructed something like this. An entity is alive if (1) it has intrinsic metabolism; (2) it belongs to, or is descended from, a group (species) that reproduces sexually, asexually, or through a mixture of the two; and (3) there is inheritance of organismic features from one generation to the next. If it has none of these features, it is not alive. Now we get to the grey area mentioned above. What happens if an entity has one or two of these attributes, rather than zero or three?
I’m quite confident that most entities on planet Earth can be classified as living or non-living on the basis of the RIM definition, as given above. However, there is an important exception: viruses. Are these entities tiny subcellular organisms, or very large and complex, but non-living, molecules? Most people think of them as being alive, but is this intuitive feeling justified? Viruses have no metabolism. At its simplest, a virus is a piece of hereditary material made of nucleic acid (sometimes DNA, sometimes RNA) wrapped in a protein coat. Its size is orders of magnitude less than that of a single cell. Viruses operate by entering host cells and hijacking their molecular machinery to make offspring viruses.
It’s clear from this brief description that viruses have the R and the I of RIM, but not the M. So they possess two out of the three attributes of life. However, this statement has to be qualified in an important way. Viruses can’t reproduce outside of a living cell of their host organism, be it an animal, a plant, a fungus, or a microbe. So their reproduction, and hence their inheritance, is not intrinsic. Perhaps we could conclude that they score half, half, and zero on the RIM scale, thus giving them an overall score of one rather than two. Whatever exact score we give them, the conclusion is inescapable: viruses inhabit a grey area between life and non-life.
A Working Definition of Intelligence
The searches for alien life in general, and intelligent alien life in particular, are very different endeavours. We’ve already seen that they involve different wavelengths on the electromagnetic spectrum, and consequently different designs of telescope. Searching for signatures of oxygen is centred on the visible and infrared parts of the spectrum. In contrast, searching for the technosignatures of intelligent civilizations is centred on longer wavelengths – those of the radio range. I’m going to use radio as a pragmatic way of defining intelligence from a SETI perspective. But let’s first look at the span of intelligences among animals on Earth from a different perspective.
Intelligence is a continuous variable, not a binary one. Life forms here, and probably elsewhere too, show a wide range of behaviours that can be interpreted as indicating a wide range of intelligence underlying them. The construction of a scale for the measurement of intelligence is notoriously difficult, even within the confines of a particular species – humans. How intelligent someone is cannot be completely captured on a single scale; intelligence is a multidimensional property. Paradoxically, though, in a multi-species context the job actually gets easier rather than harder. The reason for this is that the differences we’re dealing with are much greater.
A scale for intelligence in the context of life on Earth starts from zero and extends indefinitely upwards. Organisms that lack nervous systems are deemed to have no intelligence. These include all microbes and plants, as well as the most primitive of animals – sponges and a little-known group called placozoans (which translates as ‘flat animals’). Slightly more advanced animals such as jellyfish have diffuse nervous systems but lack brains, and don’t really exhibit any behaviour that merits the description intelligent. There are many animals that have small brains, including insects and arachnids. These typically have rather fixed, instinctive behaviour patterns, but some seem more intelligent than others. Jumping spiders – for example those of the genus Portia – have been put forward as the most intelligent arthropods, owing to their complex patterns of hunting behaviour.
The most intelligent invertebrates of all are octopuses; there’s a great book about octopus intelligence by Peter Godfrey-Smith, called Other Minds. Many studies on these wonderful creatures, both in the lab and in the field, have illustrated complex behaviours that are clearly learned rather than instinctive. Octopuses exceed many vertebrates in intelligence. However, some groups of birds seem to be particularly intelligent – notably crows and parrots. And many mammalian groups show high intelligence, especially cetaceans (dolphins and whales) and primates. Among the primates, the great apes stand out, and among these, of course, humans represent an apex.
Rather than pretend that it’s possible to create a cross-species numerical scale of intelligence, let’s simply acknowledge five broad-brush levels: zero (e.g. sponges), little if any (e.g. worms), some (e.g. jumping spiders), a lot (e.g. octopuses, crows, most mammals), and ‘high’ (humans). Counterparts of these may well eventually be found among the life forms of other planets, though there may also be a further category (‘very high’) on some of them, the nature of which we can only dimly conjecture.
Now let’s poke further into the ‘high’ category here on Earth. Human tool-making is thought to have originated about three million years ago – in the form of stones fashioned for hammering and chopping. That was a significant milestone. Wheels and writing – two further important milestones – were invented thousands rather than millions of years ago. Telescopes were invented a few hundred years ago – around 1600, as we’ve already seen. And radio transmission was achieved in the years around 1900, with the Italian scientist Guglielmo Marconi being a key figure.
Here’s an important caveat. The pattern of increasing sophistication of human inventions over time can’t be mapped directly to a pattern of increasing intelligence in time. Human brain size reached its present level more than 100,000 years ago. Brain size is a very blunt instrument, but even when more subtle aspects of the brain, such as its shape, are taken into account, the brains of members of our species – Homo sapiens – seem to have reached something resembling the present-day condition by about 50,000 years ago. Not only that, but the Neanderthals alive at that time had brains that were at least as big as ours.
So humans of 50,000 years ago probably had an intelligence level not dissimilar to that of an average human alive today. Marconi’s ability to send radio signals across the Atlantic in the early twentieth century was probably not so much a consequence of human evolution in the last 50,000 years as it was a consequence of being able to build on a wide range of discoveries and inventions that had taken place in previous history, and on the prediction of radio waves by the Scottish physicist James Clerk Maxwell in the 1860s.
It’s important to keep in mind this point that the mapping of the level of technology of a society to the level of intelligence of its members is complex. However, it’s also important to be pragmatic when trying to come up with a working definition. I’m going to use the ability to send and receive radio waves as indicating that a certain threshold of intelligence has been reached, because searches of a SETI type are largely conducted using radio, and the only beings we’ll detect by conducting these searches are those who can send out powerful signals (radio or possibly laser) into space. The relevant intelligence threshold may have been reached thousands of years earlier, but that’s not important from a practical perspective.
Two Key Questions
We’ve now considered what we’re looking with – telescopes plus various associated techniques including spectroscopy. And we’ve also thought carefully about what we’re looking for – both life in general, as defined by reproduction, inheritance and metabolism, and intelligent life with advanced technology in particular, as defined by the ability to send and receive radio signals. Our final job, before venturing out into the cosmos to take its measure, is to be clear about the key questions we’re asking in our endeavour to understand what has sometimes been called ‘the biological universe’. There are two of these questions, or perhaps more accurately two groups of questions, as follows.
First, a group that I think of as being about the geography of alien life. These are questions about distribution in space. For example: Is life only found on planets orbiting stars, or might it be found in other places too, such as rogue planets (which don’t orbit anything) or moons, which orbit planets? Where is the closest life – and the closest intelligent life – to us? How many inhabited bodies are there, at various levels of spatial scale from the solar system through the galaxy to the universe as a whole? Are there planetary systems in which more than one planet is inhabited?
Second, a group of questions that I think of as being about the biology of alien life. These are questions about the nature of life forms, wherever they are found. For example: Is life usually (or always) based on carbon? Is it usually dependent on water? Is life usually made of cells or cell-like units? Are large life forms common, and do they usually arise from the evolution of multicellularity rather than by some other means? Are there features of large multicellular life forms that are to be expected to occur widely across multiple planets, for example skeletons? How does intelligent life arise from unintelligent beginnings, and are there parallels in the evolution of intelligence from one planet to another?
Although these two groups of questions are reasonably distinct from each other, they are not without overlap and interaction, as I mentioned earlier. For example, by now we are fairly sure that, Earth aside, there are no multicellular life forms in the solar system, even though the possibility of microbial life having evolved on some local body – for example Jupiter’s moon Europa or Saturn’s moon Enceladus – remains a possibility, as we’ll see in Chapter 5. In contrast, it seems highly probable that there are plenty of rocky bodies scattered across our galaxy that have large life forms as well as microbes. Some authors have argued against this – notably the American scientists Peter Ward and Donald Brownlee in their 2000 book Rare Earth. These authors argued that Earth possessed a combination of unique features that was unlikely to be replicated elsewhere. However, that was before the discovery of Earth-like exoplanets, and their argument no longer seems tenable.
As we proceed, these two groups of questions – about the geography and biology of alien life – will always both be at least in the background. In particular chapters, one may be more to the fore than the other. For example, Chapter 2 will focus largely on the places where life might evolve, while Chapter 3 will concentrate on the nature and evolution of life on Earth, and possibly elsewhere. As the book progresses, I’ll try to tie the two groups of questions – and possible answers – together. Ideally, we’ll end up with a pretty good idea of the most likely abodes for alien life and the nature of that life, at least in general terms.


