



Introduction
Use of functionally diverse cover crop mixtures is increasingly promoted in agricultural systems to balance tradeoffs between production and environmental goals (Blesh et al. Reference Blesh, VanDusen and Brainard2019; Finney et al. Reference Finney, Murrell, White, Baraibar, Barbercheck, Bradley, Cornelisse, Hunter, Kaye, Mortensen, Mullen and Schipanski2017). Employing functional diversity via cover cropping is most often associated with use of mixtures that include grass, legume, and brassica species. Establishment of diverse mixtures following corn grain harvest in the northeastern United States is limited by short growing season windows. Drill-interseeding cover crops into standing corn can overcome this limitation and provide opportunities for increasing functional diversity (Curran et al. Reference Curran, Hoover, Mirsky, Roth, Ryan, Ackroyd, Wallace, Dempsey and Pelzer2018). However, cover crop interseeding in corn production systems introduces additional management complexities. Designing herbicide-based weed control programs that negotiate tradeoffs between crop protection and cover cropping goals in interseeded systems remains a major challenge.
Research from multiple growing regions has identified grass, legume, and brassica cover crop species that establish and persist when interseeded into corn near the V3 to V5 corn growth stage (Alonso-Ayuso et al. Reference Alonso-Ayuso, Gabriel, Pancorbo and Quemada2020; Caswell et al. Reference Caswell, Wallace, Curran, Mirsky and Ryan2019; Rusch et al. Reference Rusch, Coulter, Grossman, Johnson, Porter and Garcia y Garcia2020). Commonly used residual herbicides that are applied preemergence (PRE) have the potential to reduce establishment of postharvest seeded cover crops but often pose minimal risk (Palhano et al. Reference Palhano, Norsworthy and Barber2018; Rector et al. Reference Rector, Pittman, Beam, Bamber, Cahoon, Frame and Flessner2020; Whalen et al. Reference Whalen, Bish, Young, Hager, Conley, Reynolds, Steckel, Norsworthy and Bradley2019). In comparison, interseeding at the V3 to V5 corn growth stage, or approximately 28 to 42 d after PRE herbicide applications, significantly increases the likelihood of reduced cover crop establishment due to persistence of soil-applied herbicides at phytotoxic levels. Several field studies have evaluated the potential of PRE herbicides to injure cover crops that are drill-interseeded into corn at the V3 to V6 stage, including annual ryegrass, medium red or crimson clover, and daikon radish (Brooker et al. Reference Brooker, Sprague and Renner2020; Stanton and Haramoto Reference Stanton and Haramoto2019; Wallace et al. Reference Wallace, Curran, Mirsky and Ryan2017).
Developing herbicide recommendations for interseeding cover crop mixtures with use of field-based experiments presents a unique problem given the array of PRE herbicide mixtures containing multiple sites of action that are available in corn production systems. Field trials have shown considerable variation in response of interseeded cover crops to PRE herbicides due in part to interactions between soil and environmental factors that influence herbicide degradation rates at local to regional scales (Brooker et al. Reference Brooker, Sprague and Renner2020; Stanton and Haramoto Reference Stanton and Haramoto2019; Wallace et al. Reference Wallace, Curran, Mirsky and Ryan2017). Growers also approach herbicide decision-making from different viewpoints in the context of cover cropping. Some growers may seek to adjust cover crop species selection based on their herbicide program, while others may seek to adjust herbicide programs based on their desired cover cropping strategy and species selection.
The objective of this study was to use greenhouse-based dose-response assays to describe the relative sensitivity of 12 cover crop species that differ in traits to chloroacetamide (acetochlor, dimethenamid, S-metolachlor) and pyrazole (pyroxasulfone) herbicides (Group 15 herbicides as categorized by the Weed Science Society of America), which are frequently used in premix or tank-mix PRE applications in corn. These dose-response assays complement previous field-based research that described the likelihood of injury to interseeded annual ryegrass and medium red clover following PRE applications of these herbicides across multiple locations and years in the Mid-Atlantic United States (Wallace et al. Reference Wallace, Curran, Mirsky and Ryan2017). Greater understanding of the relative difference in herbicide sensitivity among cover crop species that differ in functional traits, such as taxonomic group and seed mass, can improve herbicide management recommendations for interseeding or postharvest seeding diverse cover crop mixtures when coupled with field-based observations of herbicide persistence and carryover injury to commonly used cover crop species.
Materials and Methods
Experimental Design
A greenhouse study consisting of a series of dose-response assays was completed over a 4-mo period (March to June) in 2018. Dose-response assays were conducted for chloroacetamide and pyrazole herbicides that provide the foundation for PRE control of annual grass and small-seeded broadleaf weed species in corn production systems (Table 1). For each herbicide, treatments were a factorial combination of herbicide rate and cover crop species. Each factorial combination was replicated five times in two experimental runs, yielding 10 total experimental units per treatment. Experimental runs were staggered 7 to 10 d apart due to space and labor constraints.
Table 1. Labeled rate and biochemical properties of herbicides used in dose-response assays.
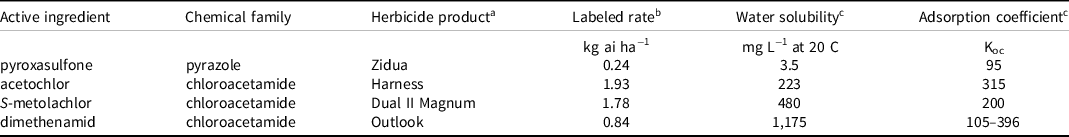
a Herbicide product suppliers: Zidua and Outlook, BASF Corporation, Florham Park, NJ; Harness, Bayer CropScience LB, St. Louis, MO; Dual II Magnum, Syngenta International AG, Basel, Switzerland.
b The labeled rate is 1×.
c For water solubility and adsorption coefficient, see Shaner (Reference Shaner2014).
Herbicide treatments were based on standard labeled rates (1×) for medium-textured soils (Table 1) and included seven treatment levels: 1×, 1/2×, 1/4×, 1/8×, 1/16×, 1/32×, and an untreated control (0×; UTC). Herbicide treatments rates were selected to simulate concentrations across five soil half-lives (Shaner Reference Shaner2014). Cover crop treatments included 12 commonly used species, representing four plant families and variation in seed mass within plant families where possible (Table 2).
Table 2. Cover crop species used in dose-response assays. a
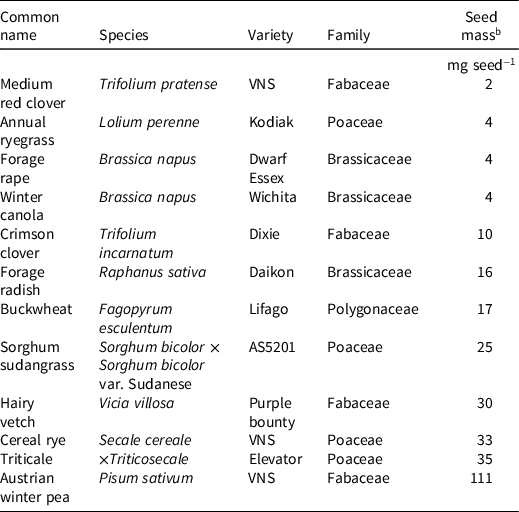
a Abbreviation: VNS, variety not stated.
b Seed mass estimated from 1,000-weight subsamples (n = 4) in each seed lot.
Experimental units were round plastic pots (10-cm diameter, 10-cm deep) filled with a 2:1 sand to potting mix (80% sphagnum peat moss, 15% vermiculite; SunGro®, Agawam, MA) ratio. The homogenized soil mix was subsampled and subjected to basic soil fertility analyses to assess attributes known to influence herbicide bioavailability. Soil pH was 6.8 and organic matter was 0.3%. Five seeds of each cover crop species were sown into assigned experimental units at a depth of 0.63 cm, and soil furrows were covered with soil and firmed to provide seed-to-soil contact prior to imposing herbicide treatments. Stock dilutions (1/2× to 1/32×) of each herbicide active ingredient were prepared from a solution containing the standard label rate (1×). Treatments were applied from low to high rates, including a water-only UTC, using a spray chamber equipped with an 80015 flat-fan nozzle (TeeJet Technologies, Glendale Heights, IL) attached to a pneumatic track positioned 45 cm above pots. Treatments were applied using a 185 L ha−1 spray volume. To avoid contamination, ammonia-water was used to flush the system between herbicide active ingredients. Experimental units were transferred to greenhouse benches immediately after application, arranged in a randomized complete block design with five replicates, and then watered (40 mL per pot) to activate herbicides. Pots were hand watered approximately every 4 d throughout the duration of the assay to maintain pots near field capacity while preventing soil water loss through drainage holes at the bottom of each pot.
Data Collection
Cover crop density and visual evaluation of herbicide symptomology was recorded 14 d after treatment (DAT). Each experimental run was terminated 28 DAT. Cover crop density was recorded and aboveground biomass was harvested, dried at 65 C for 5 d, and weighed. For each herbicide by cover crop combination, final dry matter biomass (Y; milligrams per pot; 28 DAT) was expressed as a percentage of the UTC, where UTC is the mean biomass of the 0× treatment and x is the biomass of each experimental unit for a given cover crop (i) by herbicide (j) combination within each experimental run. Data from both experimental runs were combined for model-fitting following standardization.
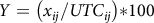 $Y = \left( {{x_{ij}}/UT{C_{ij}}} \right){\rm{*}}100{\rm{\;}}$
$Y = \left( {{x_{ij}}/UT{C_{ij}}} \right){\rm{*}}100{\rm{\;}}$
Statistical Analysis
The primary objective of dose response assays was to compare the relative sensitivity of cover crop species to each herbicide active ingredient. Consequently, nonlinear regression models were fitted by herbicide using the drc package in R software (Ritz et al. Reference Ritz, Baty, Strebig and Gerhard2015). For each herbicide, final dry matter biomass [Eq. 1] was plotted as a function of herbicide application rate and cover crop species. Three sigmoidal models were considered, including a log-logistic (Eq. 2), Weibull I (Eq. 3) and Weibull II (Eq. 4) function to allow for fitting of both symmetrical (log-logistic) and asymmetrical (Weibull I and II) relationships relative to the inflection point. In each sigmoidal model, Y is the dry matter biomass expressed as a percentage of the control (0% to 100%), d is the upper limit asymptote, x is the rate of herbicide applied as a proportion of the label rate (0 to 1), e is the value of x at the inflection point of the curve, and b is the slope of the curve at e.
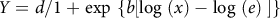 $Y = d/1 + {\rm{exp}}\ \left\{ {b\left[ {{\rm{log}}\ \left( x \right) - {\rm{log}}\ \left( e \right){\rm{\;}}} \right]} \right\}{\rm{\;}}$
$Y = d/1 + {\rm{exp}}\ \left\{ {b\left[ {{\rm{log}}\ \left( x \right) - {\rm{log}}\ \left( e \right){\rm{\;}}} \right]} \right\}{\rm{\;}}$
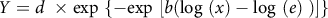 $Y = d{\rm{\;}} \times {\rm{exp}}\ \left\{ { - {\rm{exp}}\ \left[ {b\left( {{\rm{log}}\ \left( x \right) - {\rm{log}}\ \left( e \right){\rm{\;}}} \right)} \right]} \right\}$
$Y = d{\rm{\;}} \times {\rm{exp}}\ \left\{ { - {\rm{exp}}\ \left[ {b\left( {{\rm{log}}\ \left( x \right) - {\rm{log}}\ \left( e \right){\rm{\;}}} \right)} \right]} \right\}$
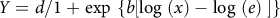 $Y = d/1 + {\rm{exp}}\ \left\{ {b\left[ {{\rm{log}}\ \left( x \right) - {\rm{log}}\ \left( e \right){\rm{\;}}} \right]} \right\}{\rm{\;}}$
$Y = d/1 + {\rm{exp}}\ \left\{ {b\left[ {{\rm{log}}\ \left( x \right) - {\rm{log}}\ \left( e \right){\rm{\;}}} \right]} \right\}{\rm{\;}}$
The upper asymptote was constrained to 100 given standardizations relative to the control resulted in a mean value of 100 at the y-intercept for each curve. Lower asymptotes were constrained to zero to reflect biological thresholds, resulting in estimation of two parameters (b, e) for each function. Inspection of model fit by cover crop species and herbicide revealed several cases where a species response was not well described by nonlinear curve fitting techniques, and parameters could not be estimated because models failed to converge. In these cases, the herbicide by cover crop species combination was excluded from nonlinear model fitting and mean biomass production (±95% confidence intervals; CI) for the labeled rate (1×) was expressed as a proportion of the untreated control.
The fit of alternative models [Equations 2 to 4] was evaluated using the mselect function, and selection of best-fit models for each herbicide was based on Akaike information criterion. For each best-fit model (Supplemental Table 1), full and reduced models were compared to evaluate the effect of cover crop species. Full models included cover crop species and herbicide dose as predictor variables, and reduced models were fit with data pooled across cover crop species. An F-test was used to assess the significance of cover species within the full model. Cover crop sensitivity to each herbicide was evaluated by calculating absolute ED50 values from best-fit models using the ED function. The biological interpretation of absolute ED50 in our models is the estimated dose that causes a 50% dry matter biomass reduction relative to the UTC. Use of absolute ED values avoid biological misinterpretation that can occur if estimates are based on inflection points (i.e., e parameter) when lower limits do not equal zero and permit relative comparisons between both symmetric and asymmetric fitted models (Keshtkar et al. Reference Keshtkar, Kudsk and Mesgaran2021). Selection of mid-point estimates (ED50) rather than tail-estimates (i.e., ED10 and ED90) was based on two considerations: 1) minimal levels of biomass reduction (i.e., 10%) are more acceptable for cover crops than cash crops, and 2) exclusion of rates above 1× inflated prediction intervals for ED90 estimates for species that showed a moderate level of tolerance to labeled rates.
The delta interval function in the drc package was used to calculate 95% confidence intervals for absolute ED50 point estimates. Within a given herbicide, point estimates with 95% confidence intervals that did not overlap were interpreted as evidence of differences in sensitivity among cover crop species to the test herbicide (Ritz and Streibig Reference Ritz and Streibig2005). Additionally, methods for evaluating relative potency between two herbicides (Ritz et al. Reference Ritz, Cedergreen, Jensen and Streibig2006) were adopted to quantify the relative sensitivity of cover crop species to each herbicide using annual ryegrass as the reference species. Estimated relativity sensitivity ratios (RA) were expressed as the herbicide dose that produces a 50% biomass reduction (ED50) in a cover crop species (x B) in proportion to the herbicide dose that produces a 50% biomass reduction in the reference species (x A). Annual ryegrass was chosen as the reference species because it is the mostly widely used and tested species in drill-interseeded systems. When RA > 1, the alternative cover crop species can be considered more tolerant to the herbicide than annual ryegrass because a higher dose is needed to produce the same level of biomass reduction. When RA < 1, the alternative cover crop species can be considered more sensitive to the herbicide than annual ryegrass. Significance testing of RA estimates were conducted using the EDcomp function.
Results and Discussion
Acetochlor
The relationship between dry matter biomass production and herbicide dose was described using a Weibull II model. High levels of biomass reduction (>50%) were observed for each cover crop species at the 1× rate. Each species was included in the full model, and the inclusion of cover crop species (i.e., full model) improved fit relative to the reduced model (F = 21.1, P < 0.001; Fig 1A).
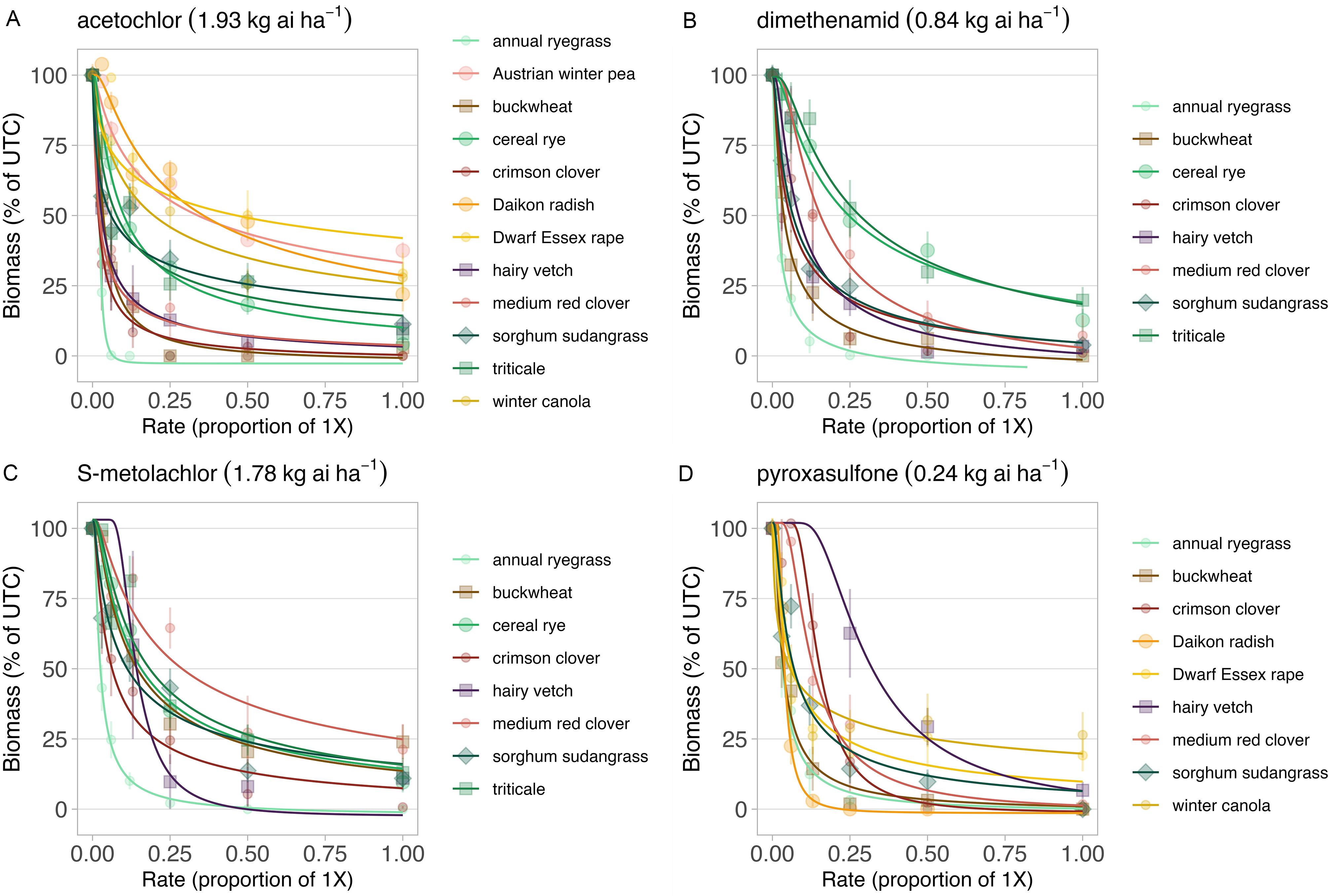
Figure 1. Best-fit models (Weibull II) describing the relationship between biomass of cover crop species and the dose of four herbicides, including (A) acetochlor (1×; 1.93 kg ai ha−1), (B) dimethenamid (1×; 0.84 kg ai ha−1), (C) S-metolachlor (1×; 1.78 kg ai ha−1), and (D) pyroxasulfone (1×; 0.24 kg ai ha−1). Cover crop biomass is expressed as a percentage of the untreated control (UTC; 0×) 28 d after treatment, and herbicide dose is expressed as the proportion of the standard labeled rate (1×) for each herbicide.
Estimated absolute ED50 values ranged from 0.03 to 0.46, respectively, with greater uncertainty (95% CI) at higher estimated values (Fig. 2A). Observed patterns indicate that brassica species are less sensitive to acetochlor compared to other taxonomic groups, with ED50 point estimates occurring between 1 and 3 simulated half-lives. Due to variability in ED50 point estimates, it is uncertain whether sensitivity differs among brassica species. Brooker et al. (Reference Brooker, Sprague and Renner2020) reported that oilseed radish was less sensitive to acetochlor than annual ryegrass and crimson clover in a greenhouse study using 1/4×, 1/2×, and 1× label rates.
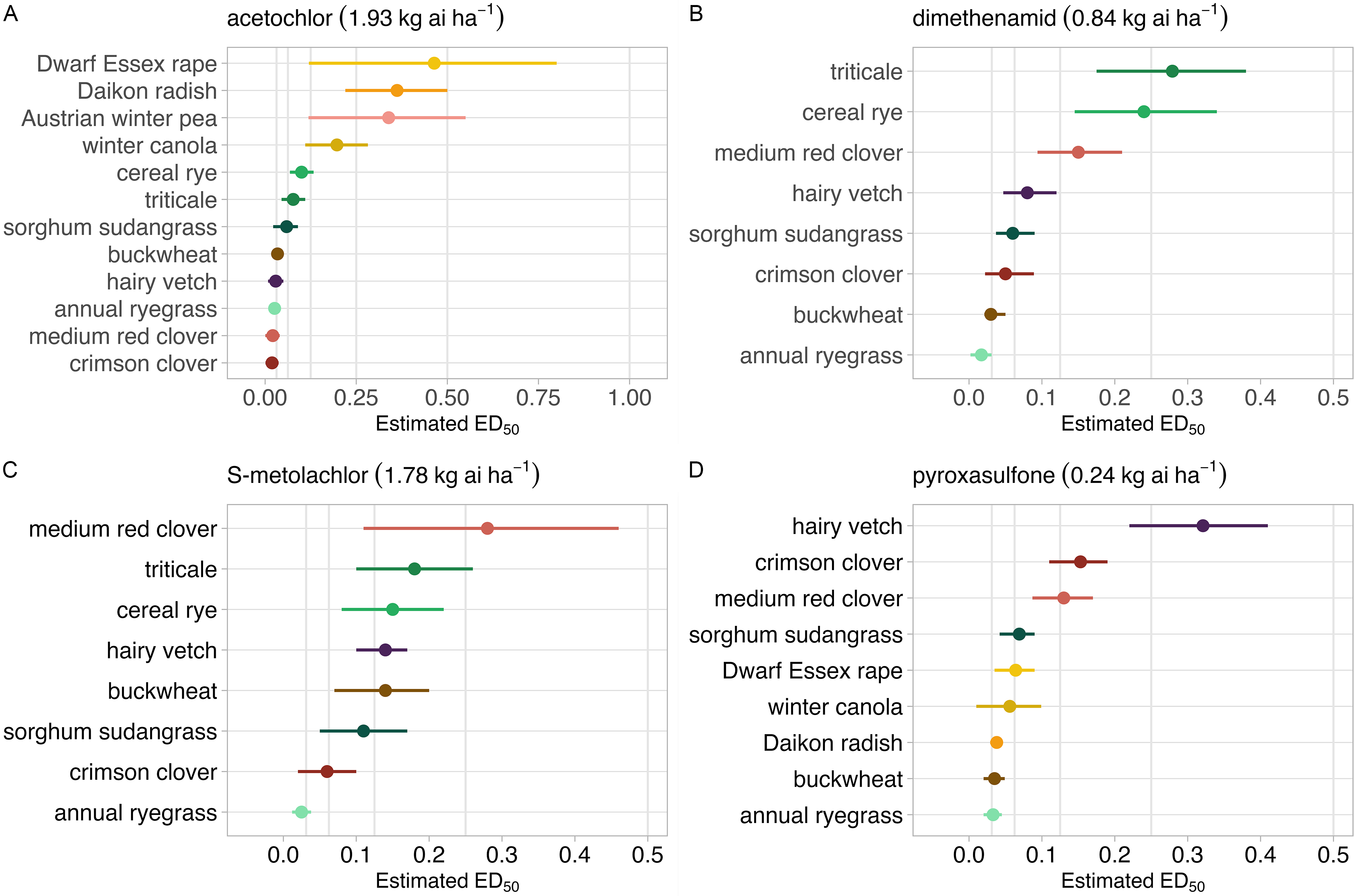
Figure 2. Model estimates of absolute ED50 values (± 95% confidence interval) for cover crop species fitted to best-fit models of dry matter biomass response to herbicide rates applied in proportion to each herbicide’s standard label rate (1×), including (A) acetochlor, (B) dimethenamid, (C) S-metolachlor, and (D) pyroxasulfone. The biological interpretation of ED50 is the estimated herbicide dose, expressed as a proportion of the 1× rate, that is needed to reduce cover crop biomass production by 50% relative to the untreated control. Cover crop species are excluded within herbicide categories when species tolerance to labeled rates precluded model fitting.
Differences in relative sensitivity were observed among legume species, where Austrian winter pea was less sensitive to acetochlor than medium red clover, crimson clover, and hairy vetch, which did not differ in comparison. Differences in relative sensitivity were also observed among grass species. Cereal rye, triticale, and sorghum sudangrass [Sorghum bicolor (× Sorghum bicolor var. Sudanese)] had a higher ED50 than annual ryegrass but did not differ in comparison. However, ED50 point estimates for all grass species were below 0.1, indicating sensitivity at low doses. The mean ED50 for buckwheat (Fagopyrum esculentum L.) did not differ compared to small-seeded species such as annual ryegrass and medium red clover. Palhano et al. (Reference Palhano, Norsworthy and Barber2018) did not observe a significant reduction in biomass across a range of cover crop species, including small- and large-seeded legumes, winter cereals, and buckwheat, following acetochlor applications at a 1/16× label rate in a field-based cover crop sensitivity study within silt-loam soil.
Field studies of acetochlor carryover injury to postharvest seeded cover crop species are highly context dependent. For example, Cornelius and Bradley (Reference Cornelius and Bradley2017) reported crimson clover and Austrian winter pea injury on silt loam soils in 2 of 3 yr when acetochlor was applied in its microencapsulated form (Warrant®; 1.26 kg ai ha−1) as a postemergence (POST) application (V2 to V3; July 2 to 28) and cover crops were seeded after soybean was harvested for forage, approximately 48 to 70 DAT. Other studies investigating postharvest seeding scenarios have found negligible effects on cover crops and few taxonomic or trait-based differences in response to microencapsulated acetochlor applied POST on silt loam to silty clay loam soils in the Midwest (102 to 167 DAT; Whalen et al. Reference Whalen, Bish, Young, Hager, Conley, Reynolds, Steckel, Norsworthy and Bradley2019), silt loam soils in the southern Mid-Atlantic (70 DAT; Rector et al. Reference Rector, Pittman, Beam, Bamber, Cahoon, Frame and Flessner2020), and to acetochlor (Harness®) applied PRE on silt loam soils in Arkansas (171 to 183 DAT; Palhano et al. Reference Palhano, Norsworthy and Barber2018).
Dimethenamid
The biomass response of brassica species and Austrian winter pea to increasing dimethenamid rates could not be fit with nonlinear models due to moderate reductions (<50%) in biomass at the highest (1×) applied rates. These species were excluded from full models. Inclusion of remaining species in full models (Weibull II) significantly improved fit compared to the reduced model (F = 18.2; P < 0.001; Fig 1B).
Estimated absolute ED50 values ranged from 0.02 to 0.28, respectively, with greater uncertainty (95% CI) at higher estimated values (Fig. 2B). Observed patterns indicate that brassica species are less sensitive to dimethenamid compared to other taxonomic groups, as evidenced by moderate biomass reduction observed at the labeled 1× rate. Among legumes, Austrian winter pea was the least sensitive species. Model results suggest that medium red clover is less sensitive to dimethenamid than crimson clover, with hairy vetch sensitivity being intermediate to clover species. Estimated ED50 values for winter cereals, cereal rye, and triticale were higher than sorghum sudangrass and annual ryegrass, with winter cereal ED50 point estimates between 1 and 2 simulated half-lives. The mean ED50 for buckwheat was 0.04.
These results indicate that brassica species and the large-seeded legume, Austrian winter pea, are significantly more tolerant to dimethenamid compared to grass species, small-seeded legumes, and buckwheat. Results also suggest that winter cereal cover crops are less sensitive to dimethenamid than annual ryegrass, and that medium red clover is less sensitive to dimethenamid than crimson clover. Similar taxonomic trends in relative sensitivity to dimethenamid were reported in previous greenhouse trials of oilseed radish, crimson clover, and annual ryegrass (Brooker et al. Reference Brooker, Sprague and Renner2020).
S-metolachlor
The biomass response of brassica species and Austrian winter pea to increasing S-metolachlor rates could not be described with nonlinear models due to moderate reductions in biomass (<50%) at the highest (1×) applied rates. These species were excluded from full models. Inclusion of remaining species in full models (Weibull II) significantly improved fit compared to the reduced model (F = 8.2; P < 0.001; Fig. 1C).
Estimated absolute ED50 values ranged from 0.03 to 0.29, respectively, with greater uncertainty (95% CI) at higher estimated values (Fig. 2C). Cereal rye, triticale, and sorghum sudangrass ED50 values did not differ, but each species was less sensitive to S-metolachlor than annual ryegrass. High levels of variability in medium red clover response to S-metolachlor rates limit the strength of inference, but trends suggest it is less sensitive to S-metolachlor than crimson clover and similar to hairy vetch. The mean ED50 for buckwheat was 0.14.
Brooker et al. (Reference Brooker, Sprague and Renner2020) reported greater S-metolachlor rates needed than evaluated in this study to reduce annual ryegrass biomass production by 50% in greenhouse trials but found similar results for oilseed radish and crimson clover. Tharp and Kells (Reference Tharp and Kells2000) reported greater reductions in annual ryegrass biomass (>95%) following S-metolachlor applications at a 1× rate compared to medium red clover (75%) and crimson clover (45%) in a greenhouse trial. Under field conditions, Palhano et al. (Reference Palhano, Norsworthy and Barber2018) did not observe a significant biomass reduction among grass and legume cover crop species following applications of S-metolachlor at a 1/16× label rate (0.05 kg ai ha−1) in silt-loam soil. In comparison, estimated ED25 and ED50 values from the best-fit model were 0.06 and 0.15 kg ai ha−1. Cornelius and Bradley (Reference Cornelius and Bradley2017) reported biomass reductions in crimson clover, hairy vetch, Austrian winter pea, and annual ryegrass in at least one experimental year following POST (V2 to V3; July 2 to 28) applications of S-metolachlor on silt loam soils when seeding cover crops (48 to 70 DAT) following soybean forage harvest. Field studies of postharvest cover crop seeding in the Midwest United States (102 to 167 DAT; Whalen et al. Reference Whalen, Bish, Young, Hager, Conley, Reynolds, Steckel, Norsworthy and Bradley2019), the southern Mid-Atlantic (70 DAT; Rector et al. Reference Rector, Pittman, Beam, Bamber, Cahoon, Frame and Flessner2020), and Arkansas (171 to 183 DAT; Palhano et al. Reference Palhano, Norsworthy and Barber2018) found minimal S-metolachlor carryover injury and no species-level interactions.
Pyroxasulfone
Austrian winter pea, triticale, and cereal rye response to pyroxasulfone rates could not be described with nonlinear models due to only moderate reductions in biomass (<50%) at the highest (1×) applied rates. These species were excluded from full models. Inclusion of remaining species in full models (Weibull II) significantly improved fit compared to the reduced model (F = 12.6; P< 0.001; Fig. 1D).
Estimated absolute ED50 values ranged from 0.03 to 0.32, respectively, with greater uncertainty (95% CI) at higher estimated values (Fig. 2D). Observed patterns indicate that winter cereals, cereal rye and triticale, and Austrian winter pea are less sensitive to pyroxasulfone than other species. Model results indicate that legume species, including hairy vetch and clovers, are less sensitive to pyroxasulfone compared to brassica species, though both taxonomic groups showed levels of sensitivity that are likely to be biologically relevant. Among legume species, estimated ED50 values for hairy vetch were higher than crimson clover and medium red clover, which did not differ in comparison. Mean ED50 values were similar among brassica species. The mean ED50 for buckwheat was 0.03.
These results indicate that the response of brassica species to pyroxasulfone is markedly different compared to chloroacetamide herbicides, with each brassica species exhibiting a high level of sensitivity (GR50; <1/16× rate). Brooker et al. (Reference Brooker, Sprague and Renner2020) reported greater GR50 values, and thus less sensitivity, of oilseed radish to pyroxasulfone when exposed to 1/4×, 1/2×, and 1× label rates in greenhouse trials. Multiple studies have reported negligible pyroxasulfone injury to postharvest seeded brassica cover crop species, including oilseed radish (48 and 70 DAT; Cornelius and Bradley Reference Cornelius and Bradley2017), daikon radish (102 to 167 DAT; Whalen et al. Reference Whalen, Bish, Young, Hager, Conley, Reynolds, Steckel, Norsworthy and Bradley2019), and rapeseed (70 DAT; Rector et al. Reference Rector, Pittman, Beam, Bamber, Cahoon, Frame and Flessner2020), which suggests that pyroxasulfone dissipation during a corn production season would typically permit postharvest seeding cover crops but early drill-interseeding of brassica species may significantly increase the likelihood of injury.
Cereal rye and triticale were comparatively less sensitive to pyroxasulfone than chloroacetamides based on ED50 point estimates from dose-response assays. This result is perhaps expected given that pyroxasulfone is labeled for PRE control of small-seeded grass weeds such as Italian ryegrass [Lolium perenne L. spp. multiflorum (Lam.) Husnot] in certain winter wheat (Triticum aestivum L.) production regions where crop tolerance has been demonstrated (Hulting et al. Reference Hulting, Dauer, Hinds-Cook, Curtis, Koepke-Hill and Mallory-Smith2012; Jones et al. Reference Jones, Taylor and Everman2021). Field-based studies of pyroxasulfone injury to winter cereals (winter wheat, triticale, and cereal rye) have produced variable results, ranging from significant reductions in biomass when seeded 48 to 70 DAT (Cornelius and Bradley Reference Cornelius and Bradley2017) to negligible effects when seeded after a full-season grain crop (Rector et al. Reference Rector, Pittman, Beam, Bamber, Cahoon, Frame and Flessner2020; Whalen et al. Reference Whalen, Bish, Young, Hager, Conley, Reynolds, Steckel, Norsworthy and Bradley2019).
Annual Ryegrass as a Reference Species
Growers are often encouraged to adapt cover crop interseeding to their corn production system by starting simply, with use of consistent performing species and best management practices before increasing cover crop species diversity to target additional ecosystem services. An important site-specific adaption will be to determine the value of chloroacetamide and pyrazole herbicides in PRE programs and the likelihood of injury to consistently performing interseeded species over a range of field conditions before introducing additional species.
Relativity sensitivity ratios (RA) were calculated using annual ryegrass as a reference species to guide cover crop species selection (Table 3). Annual ryegrass is a useful reference species for such adaptive management because it is 1) sensitive to chloroacetamide and pyrazole herbicides at low doses; 2) consistently establishes when interseeding into corn (Curran et al. Reference Curran, Hoover, Mirsky, Roth, Ryan, Ackroyd, Wallace, Dempsey and Pelzer2018; Moore and Mirsky Reference Moore and Mirsky2020; Rusch et al. Reference Rusch, Coulter, Grossman, Johnson, Porter and Garcia y Garcia2020); and 3) has been included in several field-based evaluations of PRE herbicide effects on cover crop stand establishment when interseeding in a range of U.S. crop production regions (Brooker et al. Reference Brooker, Sprague and Renner2020; Stanton and Haramoto Reference Stanton and Haramoto2019; Wallace et al. Reference Wallace, Curran, Mirsky and Ryan2017).
Table 3. Mean relative sensitivity ratio and standard errors for cover crop species by herbicide using ED50 values and annual ryegrass as the reference species. a – d
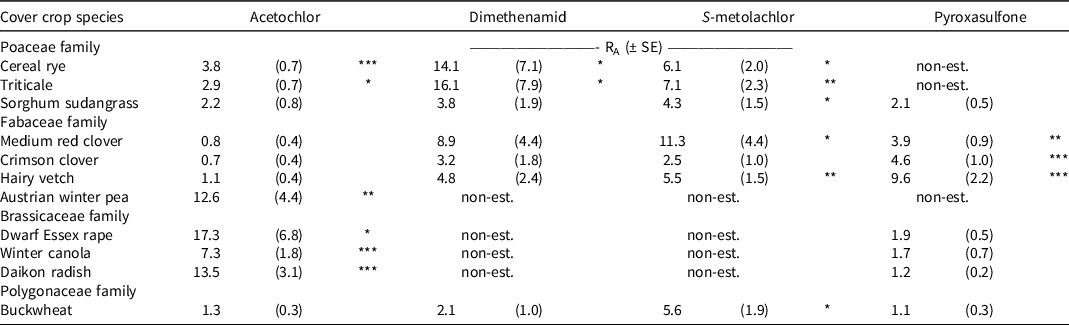
a Abbreviations: SE, standard error; RA, relative sensitivity ratio.
b An RA value >1 indicates greater tolerance to a test herbicide relative to annual ryegrass, meaning a higher dose is needed to achieve the same level (50%) of biomass production. An RA <1 indicates greater sensitivity to a test herbicide relative to annual ryegrass.
c Significance (t-test) of estimated ratio, indicating whether the value is statistically different than null (RA = 1): *P < 0.05, **P < 0.01, ***P < 0.001.
d Non-est indicates the relative sensitivity ratio (RA) is not estimable because species was excluded from fitting of full models due to tolerance at labeled rates; thus greater tolerance relative to annual ryegrass is inferred.
Results indicate that cereal rye and triticale are more tolerant than annual ryegrass to chloroacetamide and pyrazole herbicides, but sorghum sudangrass was more tolerant than annual ryegrass to only S-metolachlor. Among legumes, Austrian winter pea was more tolerant than annual ryegrass to each herbicide. Medium red clover, crimson clover, and hairy vetch sensitivity did not differ in comparison to annual ryegrass in acetochlor and dimethenamid models, but each of these legume species was more tolerant to pyroxasulfone, and medium red clover and hairy vetch were more tolerant to S-metolachlor. Brassica species were more tolerant than annual ryegrass to acetochlor, dimethenamid, and S-metolachlor but did not differ in sensitivity to pyroxasulfone. Finally, buckwheat was more tolerant to S-metolachlor than annual ryegrass, but did not differ in sensitivity to acetochlor, dimethenamid, or pyroxasulfone.
Herbicide Management Implications
Relative differences in sensitivity of cover crop species are reported by herbicide active ingredient to focus inferences on the potential for diversifying cover crop mixtures given the use of each herbicide. Caution should be used if drawing inferences about differences in sensitivity of a given cover crop species among these herbicides because of known differences in their soil persistence, and thus the likelihood of carryover injury, under field conditions. A field study that measured dissipation (days to 50% dissipation; DT50) in fine loamy soils suggests that acetochlor and dimethenamid dissipate at a faster rate (DT50; 4 to 9) than S-metolachlor (DT50 = 9 to 27) and pyroxasulfone (DT50; 8 to >71; Mueller and Steckel Reference Mueller and Steckel2011). However, published soil half-lives (DT50) from field-based trials should not be used in combination with predicted ED50 values (expressed as the number of half-lives) from this study to estimate the number of days needed to safely seed cover crop species because, as noted by Mueller and Steckel (Reference Mueller and Steckel2011), there is often poor correlation between species-level responses and herbicide concentrations that are a composite of multiple depth increments.
Poor correlation between plant responses and herbicide concentration in field studies can result from differences in herbicide mobility and sorption-desorption kinetics, which is influenced by residence time in soil (Lavy et al. Reference Lavy, Mattice, Massey, Skulman, Senseman, Gbur and Rarrett1996; Pignatello et al. Reference Pignatello, Ferrandino and Huang1993) and ultimately determines herbicide bioavailability. Across a broad range of soil types, Westra et al. (Reference Westra, Shaner, Bararick and Khosla2014a) reported that soil binding of pyroxasulfone, S-metolachlor, and dimethenamid was most positively correlated to soil organic matter among soil properties. However, the authors also found that despite comparatively lower water solubility, pyroxasulfone is more available for plant uptake in soil solution than dimethenamid and S-metolachlor due to lower sorption capacity (Kd; 1.725 mg mL−1), which likely contributes to its observed herbicidal activity at low use rates. Westra et al. (Reference Westra, Shaner, Westra and Chapman2014b) reported that the majority (>90%) of pyroxasulfone and S-metolachlor remains in the upper soil profile (0 to 7.5 cm) for several months but can move below the biologically active zone (0 to 7.5 cm) following significant rainfall events in lighter-textured (sandy loam) soils. Therefore, soil temperature and moisture content are primary drivers of microbial degradation among all chloroacetamides (Zimdahl and Clark Reference Zimdahl and Clark1982) and pyroxasulfone, whereas it is likely that precipitation events that influence downward movement below the biologically active zone and soil properties that influence sorption-desorption dynamics are key factors that determine relative differences among chloroacetamides and pyroxasulfone persistence within any given location and year.
Poor correlation between plant responses and herbicide concentration also results from intrinsic differences in the biological response of plant species to herbicides across concentration gradients. Despite its potential utility, there has been no systematic effort to describe relative sensitivity trends among cover crop taxonomic groups and traits for herbicides with similar properties and use patterns. This study begins to address that knowledge gap.
Understanding cover crop relative sensitivity patterns among chloroacetamides and pyroxasulfone will likely have the greatest impact on cover crop interseeding management decisions. For example, few cover crop species-level interactions have been reported in field trials investigating PRE use of Group 15 herbicides and postharvest seeding in grain production systems (Palhano et al. Reference Palhano, Norsworthy and Barber2018; Rector et al. Reference Rector, Pittman, Beam, Bamber, Cahoon, Frame and Flessner2020; Whalen et al. Reference Whalen, Bish, Young, Hager, Conley, Reynolds, Steckel, Norsworthy and Bradley2019). In comparison, field trials investigating either cover crop interseeding at early corn growth stages (Brooker et al. Reference Brooker, Sprague and Renner2020; Wallace et al. Reference Wallace, Curran, Mirsky and Ryan2017) or cover crop seeding following POST applications in a forage crop (48 to 70 DAT; Cornelius and Bradley Reference Cornelius and Bradley2017) have reported species-level interactions and differences in injury potential among herbicides. Mid-Atlantic field trials that evaluated the response of annual ryegrass interseeded 35 to 42 DAT observed higher levels of biomass reduction following S-metolachlor and pyroxasulfone treatments (>40%) compared to acetochlor and dimethenamid (<20%; Wallace et al. Reference Wallace, Curran, Mirsky and Ryan2017). In comparison, an Upper Midwest field study reported high levels of annual ryegrass stand reduction across acetochlor, dimethenamid, pyroxasulfone and S-metolachlor treatments (67% to 86%) when interseeding from the V3 to V6 corn growth stage (Brooker et al. Reference Brooker, Sprague and Renner2020). These field trials confirm that annual ryegrass is a sensitive indicator species (Table 3) that can inform cover crop species selection when designing mixtures.
Two additional management constraints are worth noting in the context of designing cover crop mixtures for drill-interseeding in corn. First, PRE herbicide programs in corn often include herbicides that inhibit 4-hydroxyphenylpyruvate dioxygenase (HPPD) and photosystem II (PS II) to broaden the weed control spectrum, which can increase the likelihood of injury to interseeded brassica and legume cover crops species (Brooker et al. Reference Brooker, Sprague and Renner2020; Wallace et al. Reference Wallace, Curran, Mirsky and Ryan2017). Consequently, it is likely that multiple site-of-action PRE herbicide programs will preclude interseeding diverse cover crop mixtures and opportunities for reducing soil-applied herbicide inputs with use of two-pass programs that include nonresidual POST herbicides will need to be explored. Identifying differences among cover crops species in relative sensitivity to HPPD- and PS II-inhibiting herbicides would further refine cover crop species selection in interseeded systems.
In addition, several cover crop species that demonstrated some level of tolerance to chloroacetamide and pyrazole herbicides in this study may be less suitable for interseeding in corn compared to annual ryegrass when using standard 76-cm row spacing. For example, cereal rye has been shown to be a viable option for interseeding in the Upper Midwest (Noland et al. Reference Noland, Wells, Shaeffer, Baker, Martinson and Coulter2018; Rusch et al. Reference Rusch, Coulter, Grossman, Johnson, Porter and Garcia y Garcia2020) but less suitable in the southern Mid-Atlantic (Moore and Mirsky Reference Moore and Mirsky2020). Emerging interest in using wider-row (152-cm) spacing to facilitate interseeding cover crop mixtures likely increases the range of cover crop species that can be used, including summer annual species such as sorghum sudangrass or buckwheat. Comprehensive herbicide programs that include chloroacetamides or pyrazoles will likely be needed to maintain crop protection goals in wide-row corn systems.
Negotiating tradeoffs between weed control and cover cropping goals is a significant management challenge when using interseeded cover crop mixtures to target multiple ecosystem services in corn production systems. Results of this study suggest that relative sensitivity of chloroacetamide and pyrazole herbicides differ 1) among cover crop taxonomic groups, and 2) among cover crop species differing in seed mass within taxonomic groups. Brassica species, large-seeded legumes such as Austrian winter pea, and winter cereals are significantly more tolerant to dimethenamid and acetochlor in comparison to annual ryegrass and may be considered either as mixture components or alternatives to annual ryegrass when using interseeding tactics. It will be imperative, however, to use these findings in conjunction with field-based observations of cover crop injury potential in interseeded systems.
Acknowledgments
Thanks to Elizabeth Maloney for her assistance and dedication in completing this project. No conflicts of interest are declared. This research did not receive grant support.






 Open access
Open access