


Ramadan fasting (RF) is a unique model of prolonged intermittent fasting and re-feeding in which Muslims all over the world abstain from food and water from dawn to sunset for 29 or 30 d each yearReference Malhotra, Scott, Scott, Gee and Wharton1, Reference Nomani, Hallack, Nomani and Siddiqui2, Reference Cheah, Ch'ng, Husain and Duncan3. Eating and drinking is exclusively nocturnal during Ramadan. Fasting in general has been used in medicine for medical reasons including weight management. There are many adverse effects of total fasting as well as of crash diets. However, in RF there is no malnutrition or inadequate energy intake involved.
Kidneys play an essential role in the maintenance of total body fluid volume, its composition and pH by selective re-absorption. The major work of the kidney is Na+ re-absorption. The transport of other solutes, e.g. sugars, amino acids and inorganic phosphate (Pi) depends on Na+ re-absorptionReference Dousa and Kempson4. Na serves both as a driving force, in terms of gradient (Naextracellular>Naintracellular; Nao>Nai) for translocation of solutes across brush border membrane (BBM) and as a factor modulating the interaction of Pi with Na-Pi co-transporter in the BBMReference Dousa and Kempson4, Reference Murer and Biber5. The re-absorption functions of the kidney depend on the integrity of BBM and availability of energy in terms of ATP, provided by various metabolic pathways including glycolysis and oxidative metabolismReference Murer and Biber5, Reference Kurokawa, Nagami, Yamaguchi and Kinne6.
As millions of Muslims (young and old) have for centuries abstained from food and water in the daytime during the Islamic month of Ramadan, it seems important to examine the influence of this fasting schedule on human health, especially with regard to renal metabolism and transport functions. It has been shown that RF in man increases total body fat oxidation whereas carbohydrate oxidation slows downReference El Ati, Beji and Danguir7. However, we have recently shown that Ramadan-type fasting (RTF) caused a significant increase in the activities of various enzymes involved in carbohydrate metabolism both in the intestine and liver in the ratReference Farooq, Priyamvada, Arivarasu, Salim, Khan and Yusufi8. We now hypothesize that RTF would result in specific adaptive changes in renal metabolism and transport of Pi to maintain a positive balance of various metabolites and Pi for energy production.
To address this hypothesis, RTF was mimicked experimentally in rats used as a model for human RF. The rats were fasted (12 h) and then re-fed (12 h) daily for 30 d. The influence of RTF was determined on enzymes of carbohydrate metabolism, BBM and on BBM uptake of 32Pi. Since the kidney is a heterogeneous structure consisting of several tissue zones, e.g. cortex and medulla, the effect of RTF was also examined in the homogenates and BBM prepared from different regions of the kidney. The results of the present study demonstrate that RTF caused variable alterations in metabolic activities but enhanced BBM transport capacity of 32Pi.
Materials and methods
Materials
Wistar rats were purchased from All India Institute of Medical Sciences (New Delhi, India). 32Pi was purchased from Bhabha Atomic Research Center, (Mumbai, India). Scintillation fluid (Cocktail-T) was purchased from Sisco Research Laboratory (Mumbai, India). All other chemicals used were of analytical grade and were purchased from Sigma Chemical Co. (St Louis, MO, USA) unless otherwise stated.
Experimental design
All animals were kept under conditions that prevented them from experiencing unnecessary pain and discomfort according to the guidelines approved by the Institution Ethical Committee. Adult male Wistar rats weighing 150–200 g, fed with a standard pellet diet (Amrut Laboratories, Pune, India) and water ad libitum, were conditioned for 1 week before the start of the experiment. Since rats are nocturnal feeders, the effect of RTF was initially determined on various renal enzymes and biochemical parameters in rats that were fasted for 12 h followed by 12 h re-feeding with either daytime or night-time feeding schedules for 30 d to rule out variations due to day or night food intake and a rhythmic pattern of certain enzymes as previously reported for intestinal studiesReference Farooq, Priyamvada, Arivarasu, Salim, Khan and Yusufi8. The control rats received diet and water ad libitum day and night, whereas food/water was available ad libitum to the fasted rats during re-feeding periods. It was noted that fasted rats rushed to eat and drink immediately when food/water was restored during the re-feeding period. The daily intake of food/water was carefully monitored and was found to be approximately the same for control and day/night-time fasted rats. The urine samples from control and experimental rats were collected for 4 h on the last day of fasting. The animals were killed following the last 12 h period of re-feeding. Blood was withdrawn, kidneys were extracted and cortex and medulla were separated and homogenates and BBM vesicles (BBMV) were prepared according to the method described later.
The results summarized in Tables 1 and 2 show that day/night RTF resulted in similar alterations of various parameters studied. As there was no significant difference between the respective controls, the values were pooled together to make one control value (see Results). These observations suggest that rats, whether fasted in the daytime or night-time, show similar adaptations as observed earlierReference Farooq, Priyamvada, Arivarasu, Salim, Khan and Yusufi8, Reference Saito, Murakami, Nishida, Fujisawa and Suda9. Therefore, a comprehensive effect of RTF on BBM enzymes and Pi transport was determined as described later by daytime fasting (12 h) followed by a night-time re-feeding (12 h) schedule. After 30 d of the daytime fasting schedule, the rats were killed under light ether anaesthesia, kidneys were extracted and put in ice cold buffered saline for preparing homogenates and BBMV. The body weights (before and after the experiment) and kidney/cortex weights were recorded and are summarized in Table 3.
Table 1 Effect of daytime and night-time Ramadan-type fasting‡ (Values are means with their standard errors for eight samples in each group) (A) Serum parameters
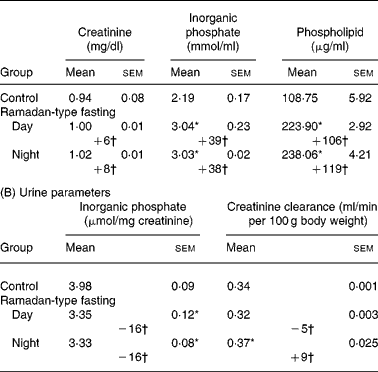
* Mean values were significantly different from control: P < 0·05.
† Values represent percentage change from control values.
‡ For details of procedures, see Materials and methods.
Table 2 Effect of a Ramadan-type fasting day on the specific activities of lactate dehydrogenase (LDH), malate dehydrogenase (MDH), glucose-6-phosphatase (G6Pase), fructose-1,6- bisphosphatase (FBPase), glucose-6-phosphate dehydrogenase (G6PDH) and malic enzyme (ME) in (A) cortical homogenate (CH) and (B) medullary homogenate (MH)‡ (Values (specific activities: μmol/mg protein per h) are expressed as means with their standard errors for three different experiments)

* Mean values were significantly different from control: P < 0·05.
† Values represent percentage change from control values.
‡ For details of procedures, see Materials and methods.
Table 3 Effect of Ramadan type fasting on body weight, kidney weight and cortex weight of rats† (Values are means with their standard errors for three different experiments with four rats in each group for each experiment)

* Values represent percentage change from control values.
† For details of procedures, see Materials and methods.
Preparation of homogenates to determine enzymes of carbohydrate metabolism
After completion of the fasting schedule, kidneys were removed, decapsulated and kept in ice cold buffered saline (154 mmol/l NaCl, 5 mmol/l Tris-HCl, pH 7·4). The cortex was carefully separated from medulla and a 15 % (w/v) homogenate was prepared in ice cold 100 mmol/l Tris-HCl, pH 7·4, using Potter-Elvehejem homogenizer (Remi motors, Mumbai, India) by passing five pulses Reference Khundmiri, Asghar, Banday, Khan, Salim, Levi and Yusufi10. The homogenate was centrifuged at 2000 g at 4°C for 10 min to remove cell debris and the supernatant fraction thus obtained was saved in aliquots and kept at − 20°C for enzyme analysis.
Preparation of brush border membrane
BBMV fractions were prepared at 4°C from homogenates of whole cortex, superficial cortex (BBMV-SC) and juxtamedullary cortex (BBMV-JMC) from the pool of the same kidneys simultaneously using MgCl2 precipitation techniques exactly as previously described Reference Khundmiri, Asghar, Khan, Salim and Yusufi11. Briefly, freshly minced tissues were homogenized in 50 mmol/l mannitol, 5 mmol/l Tris-HEPES buffer, pH 7·0, in a glass Teflon homogenizer (Thomas, PA, USA) with five complete passes. The homogenate was then subjected to a high speed Ultra-Turrex Kunkel homogenizer (Type T-25; Janke & Kunkel GMBH & Co. KG, Staufen, Germany) for three passes of 15 s each with an interval of 15 s between each pass. MgCl2 was added to the homogenate to a final concentration of 10 mmol/l and the mixture stirred for 20 min on ice. The homogenate was centrifuged at 2000 g in a Beckman centrifuge (J2 MI; Beckman Instruments Inc., Palo Alto, CA, USA) using a JA-17 rotor. The supernatant fraction was then centrifuged at 35 000 g for 30 min. The pellet was re-suspended in 300 mmol/l mannitol, 5 mmol/l Tris-HEPES buffer, pH 7·4, with four passes by a loose fitting Dounce homogenizer (Wheaton, IL, USA) in a 15 ml corex tube and centrifuged at 35 000 g for 20 min. The outer white fluffy pellet of BBM was re-suspended in buffered 300 mmol/l mannitol. Aliquots of homogenates (after high-speed homogenization) and BBMV thus prepared were saved and stored at − 20°C until further analysis for BBM enzymes. Freshly prepared BBMV were used for Pi transport. In each experiment, tissues from three to five rats were pooled to obtain a sufficient amount of starting material.
Enzyme assays
The activities of marker enzymes in the homogenate and BBM fraction were determined by standard methods as described in a previous studyReference Khundmiri, Asghar, Khan, Salim and Yusufi11. K m and V max were determined by assaying these enzymes at various substrate concentrations, 0·6–5·0 mmol/l for alkaline phosphatase (AlkPase), 0·1–0·6 mmol/l for γ-glutamyl transpeptidase (GGTase) and analysing the data by Lineweaver-Burk plot. Protein concentrations in BBMV preparations and homogenates were determined by the method of Lowry et al. Reference Lowry, Rosebrough, Farr and Randall12 as modified by Yusufi et al. Reference Yusufi, Murayama, Gapstur, Szczepanska-Konkel and Dousa13.
The activities of lactate dehydrogenase, malate dehydrogenase, glucose-6-phosphate dehydrogenase, malic enzyme involving oxidation of NADH or reduction of NADP+, were determined by measuring extinction changes at 340 nm in a spectrophotometer (Cintra 5; GBC Scientific Equipment Pty, Dandenong, Australia) as previously describedReference Farooq, Priyamvada, Arivarasu, Salim, Khan and Yusufi8, Reference Khundmiri, Asghar, Banday, Khan, Salim, Levi and Yusufi10.
Transport of 32inorganic phosphate
Measurement of 32Pi (Bhabha Atomic Research Center) uptake in BBMV was carried out at 25°C by rapid filtration technique as described by Yusufi et al. Reference Yusufi, Murayama, Gapstur, Szczepanska-Konkel and Dousa13, Reference Khundmiri, Asghar, Banday, Khan, Salim and Yusufi14 either in the presence or absence of Na-gradient. Uptake was initiated by addition of incubation medium (30 μl) containing 100 mmol/l mannitol, NaCl/KCl 100 mmol/l, 5 mmol/l Tris-HEPES, pH 7·5, 0·1 mmol/l K2H32PO4 to 15 μl BBM suspension (50–100 μg protein) and incubated for the desired time intervals (see Results). The uptake was stopped by the addition of 3 ml ice cold stop solution (containing 135 mmol/l NaCl, 5 mmol/l Tris-HEPES and 10 mmol/l sodium arsenate, pH 7·5) and filtered immediately through 0·45 μm DAWP Millipore filter, (Millipore Corporation, Billerica, MA, USA) and washed three times with the stop solution using a Cornwall type syringe (Wheaton, IL, USA). Correction for non-specific binding to filters was made by subtracting from all data the value of corresponding blank obtained by filtration of the incubation buffer without vesicles. The radioactivity of dried filters was measured by liquid scintillation counting (Reckbeta, LKB, Wallac, Sweden) with 10 ml scintillation fluid (Cocktail T; Sisco Research Laboratory).
Analysis of serum and urine parameters
Serum and urine parameters were analysed by standard methods as previously describedReference Khundmiri, Asghar, Khan, Salim and Yusufi11.
Statistical analysis
Results are expressed as means with their standard errors for at least three separate experiments. Each sample of BBM and homogenate was prepared by pooling tissues from four to five rats. The data are representative of twelve to fifteen rats per group per experiment. The data were analysed for statistical significance using Student's t test for group and/or by one way ANOVA using SPSS 7·5 software (SPSS Inc., Chicago, IL, USA).
Results
Effect of day and night-time Ramadan-type fasting on serum and urinary parameters
As reported previouslyReference Farooq, Priyamvada, Arivarasu, Salim, Khan and Yusufi8, serum glucose, cholesterol and lactate dehydrogenase activity declined similarly by day or night-time RTF (data not shown). Serum creatinine and blood urea N, two important parameters generally used to assess kidney functions, along with creatinine clearance were not affected by RTF. However, serum Pi and phospholipids both significantly increased (Table 1(A)) whereas Pi clearance was significantly lowered in RTF rats compared with control rats (Table 1(B)).
Effect of day and night-time Ramadan-type fasting on enzymes of carbohydrate metabolism in renal cortex and medulla
The effect of day/night-time RTF was determined on the activities of various enzymes involved in glucose degradation and/or its production in the homogenates of renal whole cortex and medulla. The activities of lactate dehydrogenase and malate dehydrogenase, representative enzymes of glycolysis and TCA cycle, respectively, significantly declined in cortical and medullary homogenates. The activities of glucose-6-phosphatase and fructose- 1,6-bisphosphatase, enzymes of gluconeogenesis, were also decreased similarly in RTF compared with control rats (Table 2). The activity of glucose-6-phosphate dehydrogenase (hexose monophosphate shunt) and malic enzyme, which play an important role in reducing anabolic pathways by producing NADPH, however, were differentially affected. The activity of glucose-6-phosphate dehydrogenase profoundly increased, whereas the activity of malic enzyme markedly decreased by RTF. RTF-induced changes in all the enzymes were always greater in cortical than in medullary homogenates. In addition, RTF caused a similar decrease in the activities of AlkPase and GGTase in BBMV isolated from whole cortex by the day/night fasting schedule (data not shown). The afore-mentioned results demonstrate that rats are suitable to study RF in man.
Effect of daytime Ramadan-type fasting on the structure and transport functions of the kidney
A major portion of various solutes, ions and water, are re-absorbed in the kidney by its proximal tubule across its BBMReference Murer, Werner, Reshkin, Waurin and Biber15. The BBM faces the lumen and contains a number of hydrolytic enzymes and transport systemsReference Kenny and Booth16, Reference Murer and Biber17. The activities of these BBM enzymes and transport, in particular of 32Pi, have been shown to be affected by dietary P status, drugs and hormonesReference Murer and Biber5. In the present studies the effect of RTF was examined on the activities of BBM enzymes, e.g. AlkPase and GGTase, and on the transport of 32Pi in BBMV prepared from different cortical regions.
Effect of Ramadan-type fasting on body/kidney weight and brush border membrane marker enzymes
The body weight of rats in the RTF group was slightly lowered as compared with that of the control rats, a similar insignificant decrease in kidney and cortex weight was observed in the RTF group (Table 3). As shown in Table 4, the activities of AlkPase and GGTase in homogenates and BBMV isolated from whole cortex, BBMV-SC and BBMV-JMC declined by RTF. However, there was a greater decrease in BBMV than in the respective homogenates. Further analysis showed that AlkPase activity was profoundly lowered in BBMV-SC ( − 37 %), whereas the activity of GGTase preferentially decreased in BBMV-JMC ( − 44 %) after 30 d RTF. The kinetic parameters (K m, V max) were also determined by assaying the enzymes in BBM preparations at different substrate concentrations. The results summarized in Table 5 show that RTF caused a decrease in V max for AlkPase, whereas both V max and apparent K m values decreased for GGTase by RTF. While the reduction in V max for AlkPase was largely observed in BBMV-SC, the decrease of both V max and K m values for GGTase was maximally observed in BBMV-JMC.
Table 4 Effect of Ramadan-type fasting on the specific activities of (A) alkaline phosphatase (AlkPase) and (B) γ-glutamyl transpeptidase (GGTase) in cortical homogenate (CH) and brush border membrane vesicles (BBMV) from whole cortex (WC), superficial cortex (SC) and juxtamedullary cortex (JMC)‡ (Values (specific activities: μmol/mg protein per h) are expressed as means with their standard errors for three different experiments)
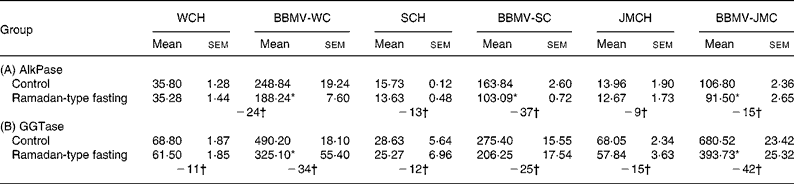
* Mean values were significantly different from control: P < 0·05.
† Values represent percentage change from control values.
‡ For details of procedures, see Materials and methods.
Table 5 Effect of Ramadan-type fasting on kinetic parameters of alkaline phosphatase (AlkPase) and γ-glutamyl transpeptidase (GGTase) in brush border membrane vesicles (BBMV) from whole cortex (WC), superficial cortex (SC) and juxtamedullary cortex (JMC)‡
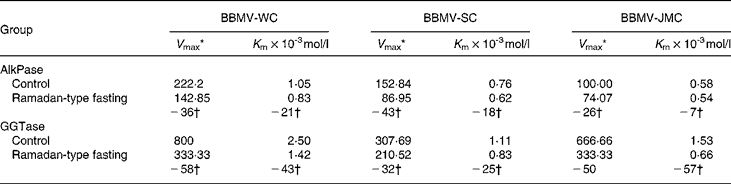
* μmol/mg protein per h.
† Values represent percent change from control values.
‡ For details of procedures, see Materials and methods.
Effect of daytime Ramadan-type fasting on Na-gradient dependent transport of 32inorganic phosphate in brush border membrane vesicles isolated from whole, superficial and juxtamedullary cortices
The bulk of filtered Pi in the kidney is re-absorbed by its proximal tubule. In this nephron segment, the initial step in lumen to interstitial Pi transport involves secondary active Na gradient-dependent (Nao>Nai) uptake of Pi across the BBMReference Dousa and Kempson4. The BBMV-SC and BBMV-JMC of rat kidney differ in their structure and transport propertiesReference Yusufi, Murayama, Gapstur, Szczepanska-Konkel and Dousa13. The effect of RTF on uptake of 32Pi was determined in the presence and absence of Na-gradient in the initial uphill phase (10 s, 30 s) and at equilibrium after 120 min in BBM preparations. The rate of concentrative uphill (10 s, 30 s) uptake of 32Pi in the presence of Na-gradient (Nao>Nai) was markedly increased in RTF compared with control values (Table 6). The concentrative uphill uptake (30 s) in relation to the equilibrium uptake (120 min), expressed as percentage overshoot (Δ%), was also increased by 30 d RTF. However, the uptake of 32Pi at the ‘equilibrium’ phase, at 120 min when Nao = Nai was not significantly different between the two groups. Also Na-independent uptake (in the absence of a Na-gradient, NaCl in the medium replaced by KCl, where Ko>Ki) of 32Pi at 30 s and 120 min were also not affected by RTF, indicating specific alterations only when Na-gradient was present.
Table 6 Effect of Ramadan-type fasting on 32inorganic phosphate uptake in brush border membrane vesicles (BBMV) from whole cortex (WC), superficial cortex (SC) and juxtamedullary cortex (JMC)‡ (Values are means with their standard errors for three different experiments)

* Mean values were significantly different from control: P < 0·05.
† Values in parentheses represent percent change from control values.
‡ For details of procedures, see Materials and methods.
Δ % overshoot at 30 s determined as a percentage change in uptake from 120 min.
To localize the effect of RTF, the uptake of Pi was also determined in BBMV-SC and BBMV-JMC. As shown in Table 6, RTF caused a preferential increase of Na-gradient dependent uptake of 32Pi in BBMV-SC compared with BBMV-JMC. The rate of Pi uptake was also determined with a wide range of Pi concentrations (0·05–1·0 mmol/l). The kinetic parameters were determined graphically from double reciprocal plots of (V max, K m) from the initial (10 s) 32Pi uptake in the presence of Na-gradient. RTF caused a marked increase of both apparent V max and K m values in various BBM preparations. However, RTF caused an increase of both V max and K m to a much greater extent in BBMV-SC than in BBMV-JMC (Table 7).
Table 7 Effect of Ramadan-type fasting on kinetic parameters of Na-dependent 32inorganic phosphate (Pi) uptake as a function of an external Pi concentration by brush border membrane vesicles (BBMV) from whole cortex (WC), superficial cortex (SC) and juxtamedullary cortex (JMC)‡

* pmol/mg protein per 10 s.
† Values represent percentage change from control values.
‡ For details of procedures, see Materials and methods.
Discussion
RF is a unique model of fasting in which Muslims the world over refrain from eating and drinking from dawn to sunset for 30 d each year and involves an unusual eating schedule. It has been established that an unusual eating schedule could induce different metabolic effectsReference Nelson, Cadotte and Halberg18. Previous human studies have reported that carbohydrate metabolism was slowed down by RF whereas fat oxidation significantly increasedReference El Ati, Beji and Danguir7. However, we recently observed that RTF in the rat model resulted in a profound increase in the activities of various enzymes involved in glycolysis, TCA cycle, gluconeogenesis and the hexose monophosphate shunt pathway in the intestine and liverReference Farooq, Priyamvada, Arivarasu, Salim, Khan and Yusufi8. Prolonged intermittent abstinence from water in addition to food for 12 h daily for 30 d may especially stress the kidney and alter its metabolic and transport functions.
We now hypothesize that RTF causes specific adaptive alterations in rat kidney to maintain a positive balance of various metabolites and Pi involved in energy production needed for various renal functionsReference Kurokawa, Nagami, Yamaguchi and Kinne6, Reference Kempson and Dousa19. As reported earlierReference Farooq, Priyamvada, Arivarasu, Salim, Khan and Yusufi8, a 30 d 12 h fasting/12 h feeding schedule in rats caused lowering of blood cholesterol and glucose with a slight decrease in body weight as observed after RF in human subjectsReference Malhotra, Scott, Scott, Gee and Wharton1, Reference Nomani, Hallack, Nomani and Siddiqui2. Serum creatinine and blood urea N as well as creatinine clearance were not unaltered by 30 d RTF, suggesting that the body adapts well and that normal kidney functions remained intact. Although various changes in urine volume, osmolarity, solutes, ions (Na+, K+) and urea were observed after RF in human subjects, there were no apparent adverse effects on kidney functionsReference Cheah, Ch'ng, Husain and Duncan3.
Prolonged intermittent RTF caused a significant decrease in lactate dehydrogenase and malate dehydrogenase activities in renal cortex and medulla. The activities of gluconeogenic enzymes, glucose-6-phosphatase and fructose-1,6-bisphosphatase were also decreased, suggesting that the enzymes involved in glucose degradation and its production were both decreased by a 30 d 12 h fasting/12 h feeding schedule in renal tissues. These observations in the kidney, however, were in contrast to the earlier observations on the influence of RTF on rat liver and intestinal carbohydrate metabolism, where RTF has resulted in increased activities of these enzymesReference Farooq, Priyamvada, Arivarasu, Salim, Khan and Yusufi8. The differential metabolic changes in different tissues can be attributed to differential adaptive mechanisms and/or to the availability of metabolites to different tissues. It might be possible that soon after feeding, the metabolites were first utilized by the intestine and liver and fell short for renal metabolic activities. The renal levels of metabolites are considered to be in equilibrium with their levels in the blood, as blood is passed through the kidney multiple times during circulation, where various metabolites are re-absorbed. This can be supported by the fact that levels of blood glucose lactate and pyruvate fell during RF in human subjectsReference Malhotra, Scott, Scott, Gee and Wharton1, Reference Nomani, Hallack, Nomani and Siddiqui2 resulting in the slowing down of the metabolismReference El Ati, Beji and Danguir7.
In addition to metabolic enzymes, the activities of AlkPase and GGTase also declined significantly in BBM preparations by RTF. AlkPase activity sharply fell in BBMV-SC whereas the activity of GGTase preferentially decreased in BBMV-JMC. The decrease in the activities of AlkPase and GGTase in the kidney was accompanied by the decrease of both V max and K m values, whereas in the intestine and liver the alterations were characterized by changes only in the V max without any effect on K m valuesReference Farooq, Priyamvada, Arivarasu, Salim, Khan and Yusufi8. It has been established that AlkPase and GGTase are differentially located and organized in the thickness of BBMReference Guder and Ross20; while AlkPase is considered to be a marker enzyme for BBMV-SC, GGTase is considered to be that of BBMV-JMCReference Yusufi, Murayama, Gapstur, Szczepanska-Konkel and Dousa13. These enzymes appeared to be differentially altered by RTF due to their locations and organization in the BBM as was demonstrated by thyroid hormone and by low PiReference Yusufi, Murayama, Keller and Dousa21, Reference Yusufi, Murayama, Szczepanska-Konkel, Hoppe and Dousa22.
Renal handling of Na+ and Pi is a critically important step in energy generation needed for many renal functionsReference Kempson and Dousa19. The bulk of filtered Na+ and Pi are re-absorbed in the renal proximal tubule by active transport across its BBMReference Dousa and Kempson4. The Na+-gradient (Nao>Nai) dependent uptake of Pi by renal proximal tubular BBM is a rate limiting step and regulated by dietary P status, hormones and drugsReference Kempson and Dousa19, Reference Yusufi, Murayama, Keller and Dousa21, Reference Yusufi, Murayama, Szczepanska-Konkel, Hoppe and Dousa22. The long-term phosphate deprivation in response to feeding a low-phosphate diet and the administration of thyroid hormones are the two experimental manoeuvres known to elicit the most pronounced long-term adaptive increases in the capacity of renal BBM for Na-gradient-energized Na-Pi co-transportReference Yusufi, Murayama, Gapstur, Szczepanska-Konkel and Dousa13, Reference Yusufi, Murayama, Keller and Dousa21. It has also been shown that BBMV-SC and BBMV-JMC differ in structure and in transport propertiesReference Yusufi, Murayama, Gapstur, Szczepanska-Konkel and Dousa13, Reference Yusufi, Murayama, Keller and Dousa21.
RTF caused a significant increase in the uptake of 32Pi by BBMV subpopulations. The increase in BBM uptake of 32Pi was specifically observed in the presence of a Na-gradient (Nao>Nai). RTF-induced adaptive increase in 32Pi uptake was characterized by increase of both the maximal rate of the transport (V max) and apparent K m. The increase in V max of 32Pi uptake can be considered as an increase in the number of newly synthesized transporters as demonstrated earlierReference Yusufi, Murayama, Keller and Dousa21, Reference Levi23, Reference Loghman-Adham24 and the changes in K m values may indicate changes in the intrinsic properties of the transporter. The increase in BBM uptake of Pi by RTF resulted in a parallel increase of serum Pi with a concomitant decrease of Pi excretion. The results are in partial agreement with the reported decrease of urinary excretion of both Na+ and Pi in human subjects after RFReference Cheah, Ch'ng, Husain and Duncan3, suggesting an increase of both Na+ and Pi re-absorption by the kidney. It has been shown that the decrease in Na+ excretion persisted even 1 month after RF in human subjectsReference Cheah, Ch'ng, Husain and Duncan3. It should be noted that decreased Na excretion could be the result of extra cellular volume contraction due to reduced water intake by an unknown mechanismReference Cheah, Ch'ng, Husain and Duncan3. The effect of RTF on 32Pi transport seems similar to the effect of a low-Pi diet, where both the Pi transport and serum Pi increased in parallelReference Yusufi, Murayama, Szczepanska-Konkel, Hoppe and Dousa22 and not like that of total fasting, where both Pi transport and serum Pi fell significantlyReference Kempson, Shah, Werness, Berndt, Lee, Smith, Knox and Dousa25.
In summary, the results of the present study suggest that rats can indeed be used as a model for RF in man to study renal metabolism and transport of Pi. Prolonged, intermittent 30 d RTF caused specific adaptive changes in the metabolic activities and transport functions in the rat kidney. While metabolism appears to be slowed down, Na-gradient dependent uptake of 32Pi was significantly enhanced, which maintains the positive balance of Pi needed for metabolic activities. Moreover, RTF seems to specifically affect the nephron subpopulations located in the superficial cortex. The results provide useful information of significant clinical importance on adaptations to unusual eating habits with restricted energy intake on renal metabolism and uptake of 32Pi.
Acknowledgements
This work was supported by a research grant (SO/SO/B-93/89) from DST to A.N.K.Y. The Council of Scientific and Industrial Research, New Delhi, India is acknowledged for the award of a Research Associate fellowship to N.F. S.J.K., M.A. and F.K. are recipients of a National Education Test fellowship from the University Grants Commission. Financial support to the department from University Grant Commission (UGC-DRF), Department of Science and Technology (DST-FIST) is also gratefully acknowledged.







