
6 results
Modeling weed seedling emergence for time-specific weed management: a systematic review
-
- Journal:
- Weed Science , FirstView
- Published online by Cambridge University Press:
- 17 April 2024, pp. 1-17
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
Understanding the timing of weed emergence is crucial to effective management. Management practices implemented too early may fail to completely control late-emerging seedlings, whereas management practices implemented too late will suffer from low efficacy. Weed emergence times reflect biological factors, such as seed dormancy and germination requirements, as well as environmental conditions. We conducted a systematic review of studies that developed models to predict weed emergence temporal patterns. We screened 1,854 studies, 98 of which were included in the final dataset. Most of the studies included were conducted in North America (51%) or Europe (30%). A wide variety of weed species (102) and families (21) were included, and many studies modeled several weeds. Grass weeds (Poaceae) were modeled most frequently (83 instances). Most weeds (40%) had base temperature
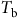 ${T_{\rm{b}}}$ values between 0 and 5 C, and 38% had base water potential
${T_{\rm{b}}}$ values between 0 and 5 C, and 38% had base water potential 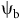 ${{\rm{\psi }}_{\rm{b}}}$ ranging from −1.0 to −0.5 MPa. Most studies used empirical parametric models, such as Weibull (40%) or Gompertz (30%) models. Nonparametric and mechanistic models were also represented. Models varied in their biological and environmental data requirements. In general, empirical parametric models based on hydrothermal time (i.e., time above base temperature and water potential thresholds) represented a good balance between ease of use and prediction accuracy. Soft computing approaches such as artificial neural networks demonstrated substantial potential in situations with complex emergence patterns and limited data availability, although they (soft computing approaches) can be susceptible to overfitting. Our study also demonstrated variability in model performance and limited generalizability across species and regions. This finding underscores the need for context-specific and well-validated weed emergence models to inform management, especially in the context of climate change.
${{\rm{\psi }}_{\rm{b}}}$ ranging from −1.0 to −0.5 MPa. Most studies used empirical parametric models, such as Weibull (40%) or Gompertz (30%) models. Nonparametric and mechanistic models were also represented. Models varied in their biological and environmental data requirements. In general, empirical parametric models based on hydrothermal time (i.e., time above base temperature and water potential thresholds) represented a good balance between ease of use and prediction accuracy. Soft computing approaches such as artificial neural networks demonstrated substantial potential in situations with complex emergence patterns and limited data availability, although they (soft computing approaches) can be susceptible to overfitting. Our study also demonstrated variability in model performance and limited generalizability across species and regions. This finding underscores the need for context-specific and well-validated weed emergence models to inform management, especially in the context of climate change.
EPSPS gene amplification confers glyphosate resistance in Palmer amaranth in Connecticut
-
- Journal:
- Weed Technology / Volume 38 / 2024
- Published online by Cambridge University Press:
- 18 March 2024, e31
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Herbicide resistance in Bromus spp.: a global review
-
- Journal:
- Weed Science / Volume 71 / Issue 5 / September 2023
- Published online by Cambridge University Press:
- 31 July 2023, pp. 422-437
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Western United States and Canada perspective: are herbicide-resistant crops the solution to herbicide-resistant weeds?
-
- Journal:
- Weed Science / Volume 70 / Issue 3 / May 2022
- Published online by Cambridge University Press:
- 09 March 2022, pp. 272-286
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Season-long weed control with sequential herbicide programs in California tree nut crops
-
- Journal:
- Weed Technology / Volume 34 / Issue 6 / December 2020
- Published online by Cambridge University Press:
- 30 June 2020, pp. 834-842
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Multiple Herbicide–Resistant Italian Ryegrass [Lolium perenne L. spp. multiflorum (Lam.) Husnot] in California Perennial Crops: Characterization, Mechanism of Resistance, and Chemical Management
-
- Journal:
- Weed Science / Volume 66 / Issue 6 / November 2018
- Published online by Cambridge University Press:
- 26 September 2018, pp. 696-701
-
- Article
- Export citation
