
71 results
Behaviour of golden hamsters (Mesocricetus auratus) kept in four different cage sizes
-
- Journal:
- Animal Welfare / Volume 16 / Issue 1 / February 2007
- Published online by Cambridge University Press:
- 11 January 2023, pp. 85-93
-
- Article
- Export citation
Effects of aviary and box sizes on body mass and behaviour of domesticated budgerigars (Melopsittacus undulatus)
-
- Journal:
- Animal Welfare / Volume 15 / Issue 4 / November 2006
- Published online by Cambridge University Press:
- 11 January 2023, pp. 353-358
-
- Article
- Export citation
A scoring system to evaluate physical condition and quality of life in geriatric zoo mammals
-
- Journal:
- Animal Welfare / Volume 16 / Issue 3 / August 2007
- Published online by Cambridge University Press:
- 11 January 2023, pp. 309-318
-
- Article
- Export citation
Altered frontolimbic activity during virtual reality-based contextual fear learning in patients with posttraumatic stress disorder
-
- Journal:
- Psychological Medicine / Volume 53 / Issue 13 / October 2023
- Published online by Cambridge University Press:
- 05 January 2023, pp. 6345-6355
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Bacterial infections and antibiotic utilization varies by coronavirus disease 19 (COVID-19) severity in hospitalized cancer patients: Analysis from the first phase of the pandemic
- Part of
-
- Journal:
- Infection Control & Hospital Epidemiology / Volume 44 / Issue 3 / March 2023
- Published online by Cambridge University Press:
- 26 May 2022, pp. 413-419
- Print publication:
- March 2023
-
- Article
- Export citation
Assessing the infection risk of a vertical garden in a hospital setting
-
- Journal:
- Infection Control & Hospital Epidemiology / Volume 43 / Issue 2 / February 2022
- Published online by Cambridge University Press:
- 18 February 2021, pp. 273-275
- Print publication:
- February 2022
-
- Article
-
- You have access
- HTML
- Export citation
Chapter Thirteen - Artificial Dignity: The Humanizing and Dehumanizing Implications of Polanyi versus Turing’s Ontology
-
-
- Book:
- The Inherence of Human Dignity
- Published by:
- Anthem Press
- Published online:
- 25 February 2022
- Print publication:
- 15 February 2021, pp 217-232
-
- Chapter
- Export citation
The cocked hat: formal statements and proofs of the theorems
-
- Journal:
- The Journal of Navigation / Volume 74 / Issue 3 / May 2021
- Published online by Cambridge University Press:
- 15 February 2021, pp. 713-722
- Print publication:
- May 2021
-
- Article
- Export citation
-
Navigators have been taught for centuries to estimate the location of their craft on a map from three lines of position, for redundancy. The three lines typically form a triangle, called a cocked hat. How is the location of the craft related to the triangle? For more than 80 years navigators have also been taught that, if each line of position is equally likely to pass to the right and to the left of the true location, then the likelihood that the craft is in the triangle is exactly 1/4. This is stated in numerous reputable sources, but was never stated or proved in a mathematically formal and rigorous fashion. In this paper we prove that the likelihood is indeed 1/4 if we assume that the lines of position always intersect pairwise. We also show that the result does not hold under weaker (and more reasonable) assumptions, and we prove a generalisation to
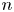 $n$ lines.
$n$ lines.
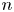
FC10-02 - Ghrelin affects sleep, secretion of cortisol and growth hormone and psychopathology in patients with major depression
-
- Journal:
- European Psychiatry / Volume 26 / Issue S2 / March 2011
- Published online by Cambridge University Press:
- 16 April 2020, p. 1865
-
- Article
-
- You have access
- Export citation
Duloxetine increases stage 3 sleep and suppresses rapid eye movement (REM) sleep in patients with major depression
-
- Journal:
- European Psychiatry / Volume 22 / Issue S1 / March 2007
- Published online by Cambridge University Press:
- 16 April 2020, p. S234
-
- Article
-
- You have access
- Export citation
Sleep and neuroendocrinology
-
- Journal:
- European Psychiatry / Volume 17 / Issue S1 / May 2002
- Published online by Cambridge University Press:
- 16 April 2020, p. 72s
-
- Article
-
- You have access
- Export citation
Pindolol in panic disorder
-
- Journal:
- European Psychiatry / Volume 17 / Issue S1 / May 2002
- Published online by Cambridge University Press:
- 16 April 2020, p. 163s
-
- Article
-
- You have access
- Export citation
A meta-analysis of gene (5-HTT) × environment interactions in eating pathology using secondary data analyses
-
- Journal:
- European Psychiatry / Volume 33 / Issue S1 / March 2016
- Published online by Cambridge University Press:
- 23 March 2020, p. S83
-
- Article
-
- You have access
- HTML
- Export citation
Serum Proteins Alter the Ability of Neutrophils to Phagocytose Malaria Related Hemozoin Crystals
-
- Journal:
- Microscopy and Microanalysis / Volume 26 / Issue S1 / March 2020
- Published online by Cambridge University Press:
- 28 April 2020, pp. 153-154
- Print publication:
- March 2020
-
- Article
-
- You have access
- Export citation
Die Zisterzienser im Mittelalter. Georg Mölich, Norbert Nussbaum, and Harald Wolter-von dem Knesebeck, eds. Cologne: Böhlau Verlag, 2017. 394 pp. €52.
-
- Journal:
- Renaissance Quarterly / Volume 71 / Issue 4 / Winter 2018
- Published online by Cambridge University Press:
- 14 January 2019, pp. 1523-1524
- Print publication:
- Winter 2018
-
- Article
- Export citation
14 - Biogeomorphic Ecosystems within Stressful and Disturbed Environments: A Focus on Termites and Pioneer Plants
- from Part IV - Coupling Fluvial and Aeolian Geomorphology, Hydrology/Hydraulics, and Ecosystems
-
-
- Book:
- A Biogeoscience Approach to Ecosystems
- Published online:
- 27 October 2016
- Print publication:
- 13 October 2016, pp 456-474
-
- Chapter
- Export citation
12 - Enforcing international humanitarian law through human rights bodies
- from Part III - International organizations as actors for ensuring compliance
-
-
- Book:
- Inducing Compliance with International Humanitarian Law
- Published online:
- 05 September 2015
- Print publication:
- 18 September 2015, pp 263-299
-
- Chapter
- Export citation
38 - REM sleep in patients with depression
- from Section V - Functional significance
-
-
- Book:
- Rapid Eye Movement Sleep
- Published online:
- 07 September 2011
- Print publication:
- 14 July 2011, pp 383-394
-
- Chapter
- Export citation
Multiscale Modeling of a Quantum Dot Heterostructure
-
- Journal:
- MRS Online Proceedings Library Archive / Volume 1370 / 2011
- Published online by Cambridge University Press:
- 07 July 2011, mrss11-1370-yy10-02
- Print publication:
- 2011
-
- Article
- Export citation
DIVISION II: SUN and HELIOSPHERE
-
- Journal:
- Proceedings of the International Astronomical Union / Volume 6 / Issue T27B / December 2010
- Published online by Cambridge University Press:
- 14 May 2010, pp. 146-157
- Print publication:
- December 2010
-
- Article
-
- You have access
- Export citation
