



Introduction
Members of the species complex Contracaecum rudolphii parasitize mainly cormorants (family Phalacrocoracidae) worldwide, with 6 sibling species displaying characteristic geographic distribution and host preference. Particularly, C. rudolphii A has been described in the great cormorant Phalacrocorax carbo sinensis (Mattiucci et al., Reference Mattiucci, Turchetto, Brigantini and Nascetti2002, Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020; Amor et al., Reference Amor, Farjallah, Piras, Burreddu, Garippa and Merella2020; Carmeno et al., Reference Carmeno, Rusconi, Castelli, Prati, Bragoni, Santoro, Postiglione, Sassera and Olivieri2022; Cammilleri et al., Reference Cammilleri, D'Amelio, Ferrantelli, Costa, Buscemi, Castello, Bacchi, Goffredo, Mancini and Cavallero2023), in the European shag Ph. aristotelis aristotelis (Abollo et al., Reference Abollo, Gestal and Pascual2001) and Ph. aristotelis desmarestii (Roca-Geronès et al., Reference Roca-Geronès, Fisa, Montoliu, Casadevall, Tobella, Bas, Palomba and Mattiucci2023) from Europe; C. rudolphii B parasitizes Ph. carbo sinensis (Mattiucci et al., Reference Mattiucci, Turchetto, Brigantini and Nascetti2002, Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020; Amor et al., Reference Amor, Farjallah, Piras, Burreddu, Garippa and Merella2020; Carmeno et al., Reference Carmeno, Rusconi, Castelli, Prati, Bragoni, Santoro, Postiglione, Sassera and Olivieri2022; Cammilleri et al., Reference Cammilleri, D'Amelio, Ferrantelli, Costa, Buscemi, Castello, Bacchi, Goffredo, Mancini and Cavallero2023) from Europe; C. rudolphii C is reported in the double-crested cormorant Ph. auritus from the USA (D'Amelio et al., Reference D'Amelio, Barros, Ingrosso, Fauquier, Russo and Paggi2007), while C. rudolphii D and C. rudolphii E are reported in Ph. carbo and Ph. varius from Australia (Shamsi et al., Reference Shamsi, Norman, Gasser and Beveridge2009). Moreover, C. rudolphii F is reported in the brown pelican Pelecanus occidentalis (family Pelecanidae) from the Gulf of Mexico (D'Amelio et al., Reference D'Amelio, Cavallero, Dronen, Barros and Paggi2012).
The distribution of these sibling species is shaped by the feeding ecology and migratory movements of their definitive hosts. Cormorants are piscivorous birds, feeding on a wide variety of marine, brackish and freshwater fish. The continuous ingestion of fish paratenic hosts results, in many occasions, in massive infections in cormorants, which are sometimes associated with severe gastric lesions (Rokicki et al., Reference Rokicki, Sołtysiak, Dziekońska-Rynko and Borucińska2011). In European cormorants, the sibling species C. rudolphii A and B are reported, with C. rudolphii sp. A having a life cycle more adapted to brackish and marine ecosystems, while C. rudolphii B occurring mainly in freshwater ecosystems (Mattiucci et al., Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020; Roca-Geronès et al., Reference Roca-Geronès, Fisa, Montoliu, Casadevall, Tobella, Bas, Palomba and Mattiucci2023). In central and eastern Europe, both species are found, often in mixed infections, in Ph. carbo sinensis (Mattiucci et al., Reference Mattiucci, Turchetto, Brigantini and Nascetti2002), which are thought to feed in different environments during their migration (Frederiksen et al., Reference Frederiksen, Korner-Nievergelt, Marion and Bregnballe2018). The great cormorant Ph. carbo sinensis is a cosmopolitan species widely distributed in all continents (Battisti et al., Reference Battisti, Taffon and Giucca2008; Davidovich et al., Reference Davidovich, Yasur-Landau, Behar, Pretto and Scholz2023a) and its diet is essentially represented by fish.
In Italy, larval stages of C. rudolphii s.l. are reported from a number of fish species; particularly C. rudolphii A is found in fish from brackish and marine ecosystems, including European seabass Dicentrarchus labrax (Paggi et al., Reference Paggi, D'Amelio, Ortis, Mattiucci, Mariniello, Di Cave and Orecchia1998; Culurgioni et al., Reference Culurgioni, Sabatini, De Murtas, Mattiucci and Figus2014; Guardone et al., Reference Guardone, Susini, Castiglione, Ricci, Corradini, Guidi and Armani2020; Mattiucci et al., Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020), gilthead sea bream Sparus aurata (Guardone et al., Reference Guardone, Susini, Castiglione, Ricci, Corradini, Guidi and Armani2020), common sole Solea solea and, gobies Gobius niger and G. paganellus (Culurgioni et al., Reference Culurgioni, Sabatini, De Murtas, Mattiucci and Figus2014); several studies also document its occurrence in European eel Anguilla anguilla from brackish waters and coastal lagoons (Paggi et al., Reference Paggi, D'Amelio, Ortis, Mattiucci, Mariniello, Di Cave and Orecchia1998; Culurgioni et al., Reference Culurgioni, Sabatini, De Murtas, Mattiucci and Figus2014; Dezfuli et al., Reference Dezfuli, Manera, Bosi, DePasquale, D'Amelio, Castaldelli and Giari2016; Mattiucci et al., Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020). Conversely, C. rudolphii B has been reported in fish from freshwater environments, including chub Squalius cephalus, barbel Barbus barbus, goldfish Carassius carassius and big-scale sand smelt Atherina boyeri sampled from river and lake systems in central Italy (Mattiucci et al., Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020), and in common bream Abramis brama and in European carp Cyprinus carpio from other parts of Europe (Molnár et al., Reference Molnár, Székely, Baska, Müller, Zuo, Kania, Nowak and Buchmann2019). Interestingly, the absence of C. rudolphii B in fish sampled in brackish water was recently reported (Mattiucci et al., Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020), which strengthens the hypothesis that this sibling species has a life cycle adapted to freshwater ecosystems.
With respect to adult stages, parasitological data on the occurrence of sibling species of C. rudolphii in great cormorants are available from different areas, particularly from brackish and freshwater ecosystems of northeastern and central Italy (Li et al., Reference Li, D'Amelio, Paggi, He, Gasser, Lun, Abollo, Turchetto and Zhu2005; Mattiucci et al., Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020), from coastal brackish water ponds in Sardinia (Amor et al., Reference Amor, Farjallah, Piras, Burreddu, Garippa and Merella2020), from freshwater ecosystems of the pre-Alpine area (Carmeno et al., Reference Carmeno, Rusconi, Castelli, Prati, Bragoni, Santoro, Postiglione, Sassera and Olivieri2022) and from coastlines and the waterways of different regions in southern Italy (Cammilleri et al., Reference Cammilleri, D'Amelio, Ferrantelli, Costa, Buscemi, Castello, Bacchi, Goffredo, Mancini and Cavallero2023).
Geographically, Israel has a unique position with respect to migratory routes of many bird species that winter in Africa, with tens of thousands of piscivorous birds also staying and over-wintering in Israel (Nemtzov, Reference Nemtzov2002). The only Contracaecum species described so far in Israel in birds are C. micropapillatum, C quadripapillatum, C. gibsoni and, C. multipapillatum E (Caffara et al., Reference Caffara, Tedesco, Davidovich, Locke, Gustinelli, King, Nuytten, Nuzzo and Fioravanti2023); larval stages of C. quadripapillatum and C. multipapillatum E have also been described in fish (Davidovich et al., Reference Davidovich, Tedesco, Caffara, Yasur-Landau, Gustinelli, Drabkin, Minkova, Aflalo, Morick and Fioravanti2022, Reference Davidovich, Tedesco, Caffara, Luci, Cantori, Morick, Fioravanti and Gustinelli2023b), while C. multipapillatum s.l. has been reported by Smirnov et al. (Reference Smirnov, Ofek and Ron2021). To the author's knowledge, no information on the distribution of sibling species of C. rudolphii in piscivorous birds or other intermediate/paratenic hosts from Israel are available so far.
This study aimed to investigate the occurrence and distribution patterns of C. rudolphii A and B in Ph. carbo sinensis collected in Italy and Israel, to acquire further information about the geographical range of these sibling species and to gain deeper knowledge on the ecology of these parasites and their bird host.
Materials and methods
Contracaecum sampling
Two thousand three hundred eighty-three nematodes of the genus Contracaecum were collected from the gastric mucosa of 28 great cormorant (P. carbo sinensis): 18 birds were shot from 7 localities in Israel under permits of the Israel Nature and Parks Authority 2020/42659 and 2021/42855; while 10 birds were found dead from 3 localities in Italy. In Fig. 1 (Fig. 1A, B), the number of birds collected in each locality of both countries are reported.

Figure 1. Maps of Israel (A) and Italy (B) with detail of the sampling localities together with the numbers of Phalacrocorax carbo sinensis collected from each locality.
The nematodes were washed in saline and preserved in 70% ethanol for morphological and molecular analyses. For some adults, the anterior and posterior portions were preserved in 10% neutral-buffered formalin for SEM.
Molecular study
For molecular analysis, genomic DNA was extracted from 683 adult males by a fast DNA extraction method using Chelex®100 (Sigma-Aldrich, Darmstadt, Germany) (Caffara et al., Reference Caffara, Tedesco, Davidovich, Locke, Gustinelli, King, Nuytten, Nuzzo and Fioravanti2023). The ITS rDNA was amplified with primers NC5_f (5′-GTAGGTGAACCTGCGGAAGGATCATT-3′) and NC2_r (5′-TTAGTTTCTTCCTCCGCT-3′) (Zhu et al., Reference Zhu, Gasser, Podolska and Chilton1998) and then 10 μl were digested with the restriction endonucleases MspI (C. rudolphii cut = 700–300 bp) and then with NsiI to distinguish C. rudolphii A (cut = ~ 850–49 bp) and B (cut = uncut) (modified from Zhu et al., Reference Zhu, D'Amelio, Gasser, Yang, Paggi, He, Lin, Song, Ai and Li2007). A fragment of the cox2 mtDNA was also amplified with primers 211_f (5′-TTTTCTAGTTATATAGATTGRTTTYAT-3′) and 210_r (5′-CACCAACTCTTAAAATTATC-3′) of Mattiucci et al. (Reference Mattiucci, Paoletti, Webb, Sardella, Timi, Berland and Nascetti2008) following the same protocol. From 53 specimens, both the ITS rDNA and cox2 mtDNA were sequenced with an ABI 3730 DNA analyser (StarSEQ, Mainz, Germany) after purification by Nucleo-Spin Gel and PCR Clean-up (Mackerey-Nagel, Düren, Germany). The DNA trace files were assembled with Contig Express (VectorNTI Advance 11 software, Invitrogen, Carlsbad, CA, USA), and the consensus sequences of the ITS rDNA and cox2 mtDNA were compared with published data by BLAST tools (https://blast.ncbi.nlm.nih.gov/Blast.cgi). Multiple sequence alignments were performed using BioEdit 7.2.5 (Hall, Reference Hall1999), p-distance and maximum-likelihood (ML) tree (GTR + G + I substitution model for ITS, bootstrap of 1000 replicates) were obtained using MEGA 7 (Kumar et al., Reference Kumar, Stecher and Tamura2016). The ITS1 and ITS2 rDNA sequences were concatenated (after deleting the 5.8S rDNA) and used to build a ML tree together with the sequences of Contracaecum spp. reported by Mattiucci et al. (Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020) and Ascaris suum (MH030604) as outgroup. The cox2 mtDNA gene was also aligned with the sequences reported by Mattiucci et al. (Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020), plus Pseudoterranova ceticola (DQ116435) and Anisakis pegreffii (MT912471) as outgroups. The phylogenesis was performed by the Bayesian analysis (BI) with MrBayes 3.2.7a software (Ronquist et al., Reference Ronquist, Teslenko, van der Mark, Ayres, Darling, Höhna, Larget, Liu, Suchard and Huelsenbeck2012), with GTR + G model, 4 heated Markow chains runs for 200 000 generations with sampling frequency set at 500, discarding the first 25% of the samples from the cold chain. Posterior probabilities were estimated to assess support for each branch (significant support >0.90). To infer the population genetics of C. rudolphii A and C. rudolphii B, 52 sequences newly generated plus 89 sequences of the 2 siblings from Italy, Spain and 1 from Poland, retrieved from GenBank were aligned and analysed by DnaSP V6.12.03 (Rozas et al., Reference Rozas, Ferrer-Mata, Sánchez-DelBarrio, Guirao-Rico, Librado, Ramos-Onsins and Sánchez-Gracia2017): number of haplotypes (hn), diversity of haplotypes (hd), private haplotype (ph), and nucleotide diversity were determined. TCS network of haplotypes was constructed by PopART (Clement et al., Reference Clement, Posada and Crandall2000).
The sequences generated in this study have ̴been deposited in GenBank under accession numbers OR263194-OR263246 (ITS rDNA) and OR269666-OR269717 (cox2 mtDNA).
Morphological study
For morphological study 93 males and 13 females, randomly selected, were observed under a dissection microscope to evaluate gross morphology and to record total length (TL), then by light microscope (Leica Microsystems, Wetzlar, Germany) with the aid of a digital Nikon DS-Fi1 camera and image-acquisition software (Nikon Nis-Elements D3.0). The central part of the worms, devoid of taxonomic informative features was removed for DNA extraction. Anterior and posterior portions of the parasite body were clarified in Amman's lactophenol to measure internal structures. Morphometric analysis was carried out following Yamaguti (Reference Yamaguti1935), Hartwich (Reference Hartwich1964), and Baruš et al. (Reference Baruš, Sergeeva, Sonin, Ryzhikov, Ryšavý and Ryzhikov1978).
For scanning electron microscopy (SEM), anterior and posterior portions of male and female specimens of genetically identified C. rudolphii A and C. rudolphii B were dehydrated through a graded ethanol series, dried in hexamethyldisilazane, sputter-coated with gold palladium, and observed using a Phenom XL G2 Desktop SEM (Thermo Fisher Scientific, Eindhoven, The Netherlands) operating at 5 kV.
Results
Molecular analyses
All the 683 Contracaecum subjected to PCR-RFLP confirmed they belong to C. rudolphii s.l. and, 300 were members of the sibling species C. rudolphii A (NsiI = 840-49 bp) while 383 were C. rudolphii B (NsiI = uncut) (Fig. 2). Table 1 reports the distribution of the 2-sibling species together with the sampling locality for both countries under study. In most cases we detected mixed infections between the 2-sibling species, especially in cormorants from Italy.

Figure 2. PCR-RFLP pattern of C. rudolphii A and C. rudolphii B after digestion with MspI (C. rudolphii s.l.: 700-300 bp) and NsiI (C. rudolphii A: uncut; C. rudolphii B: 850-49 bp).
Table 1. Distribution of the adults of C. rudolphii A and C. rudolphii B collected from great cormorant and the number of specimens identified by molecular methods, together with the sampling locality and years of sampling for both countries under study

All the sequences of the ITS rDNA of 53 specimens were of good quality and the BLAST search returned 99–100% similarity with C. rudolphii A (20 specimens) and C. rudolphii B (33 specimens). Among each sibling species the sequences were identical to each other and showed a p-distance of 0.2% between A and B-F and 0.1–0.2% between B and A-F. Interestingly the alignment of the siblings A and B newly obtained in the present study together with the other sibling species in the C. rudolphii complex showed the presence of an indel represented by an insertion of ‘GTTCGTGTG’ in all but not in C. rudolphii B. The ML tree showed a well resolved branch (99%) with a cluster containing C. rudolphii s.l., with C. rudolphii B basal to all the rest of the sibling species of the complex (Fig. 3).

Figure 3. Maximum-likelihood tree based on the concatenated ITS1-ITS2 rDNA sequences showing the relationship between C. rudolphii A (condensed, containing 20 newly generated sequences: GenBank accession numbers OR263194, OR263197-OR263205, OR263223-24, OR263226-34, plus 2 concatenated reference sequences AJ634782 + AJ634785 of C. rudolphii A, Li et al., Reference Li, D'Amelio, Paggi, He, Gasser, Lun, Abollo, Turchetto and Zhu2005), C. rudolphii B (condensed, containing 33 sequences, GB acc. n. OR263195-96, OR263202, OR263206-22, OR263225, OR263235-46, plus 2 concatenated reference sequences AJ634783 + AJ634786 of C. rudolphii B, Li et al., Reference Li, D'Amelio, Paggi, He, Gasser, Lun, Abollo, Turchetto and Zhu2005) described in the present study (in bold) and the congeneric Contracaecum species. The tree is drawn to scale, with branch length measured in the number of substitutions per site.
Concerning the cox2 mtDNA gene, 52 sequences were newly generated (20 C. rudolphii A and 32 C. rudolphii B). Among C. rudolphii A the p-distance was 0–0.2%, while among C. rudolphii B was 0–0.4%; between the 2 siblings were 0.8–0.9% as with C. rudolphii F (the only cox2 available). Moreover, the alignment of the 2 siblings showed the presence of several transitions: 4 A/G, 11 G/A, 1 C/T and 1 T/C (C. rudolphii A / C. rudolphii B), that are the only variations observed between the 2 siblings. The BLAST search gave 99–100% similarity with the 2 siblings, respectively. The BI inference phylogenetic tree obtained indicate that C. rudolphii A and C. rudolphii B form 2 distinct clades with high probability values (Fig. 4) including the same siblings retrieved from GB (MK496476 and MK496482, respectively) and with C. rudolphii F more related to C. rudolphii B. Similarly, the other Contracaecum species form a well-supported clade separated from the abovementioned siblings.

Figure 4. Bayesian inference (BI) tree based on the cox2 mtDNA sequences showing the relationship between C. rudolphii A (condensed, containing 20 sequences, GB acc. n. OR269671, OR269673, OR269678-79, OR269681-88, OR269690-92, OR269705-09, plus 1 reference sequence MK496476 of C. rudolphii A, Mattiucci et al., Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020), C. rudolphii B (condensed, containing 32 sequences, GenBank accession numbers: OR269666-70, OR269672, OR269674-77, OR269680, OR269689, OR269693-OR269704, OR269710-17, plus 1 reference sequence MK496482 of C. rudolphii B, Mattiucci et al., Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020) described in the present study (in bold) and the congeneric Contracaecum species. The posterior probability is reported for each branch.
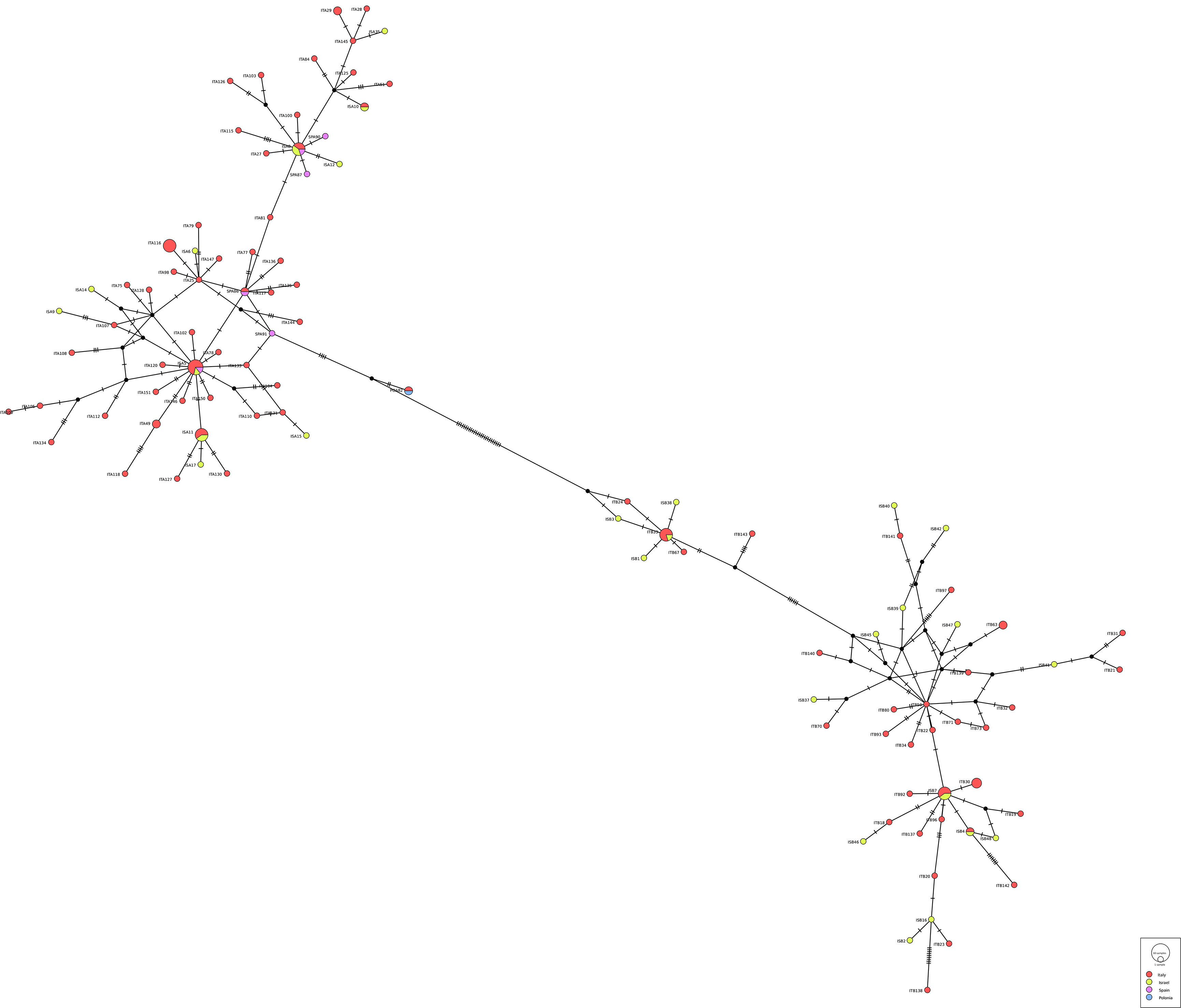
Regarding the genetic diversity between the 2 siblings, the number of haplotypes obtained among 141 sequences (83 C. rudolphii A + 58 C. rudolphii B) from Italy, Israel and Spain (plus 1 from Poland) was 106, of which 92 were private haplotypes with a haplotype's diversity of 0.99 (±0.003) and a nucleotide diversity of 0.044 (±0.0013). Concerning the haplotypes frequency, the most represented is the Hap12 of the C. rudolphii A haplogroup shared by 7 specimens from all the geographical regions (Italy, Israel and Spain). The second most representative group of haplotypes are Hap17-18 and 80, all belonging to C. rudolphii A haplogroup, containing 5 mixed geographical haplotypes each except the latter (Hap80) composed only by specimens from Sardinia. Finally, the haplotypes 3 and 14 contain C. rudolphii B from Italy and Israel. In any case the 2 siblings (A and B) never mixed together. Analysing the genetic diversity among each sibling, we observed in C. rudolphii A 60 haplotypes out of 83 sequences analysed, most of them private, with a haplotype's diversity of 0.984 (±0.006) and a nucleotide diversity of 0.0105 (±0.00062); while among C. rudolphii B haplotypes were 46 out of 58 sequences with a similar haplotype's diversity of 0.985 ± 0.008 and a nucleotide diversity of 0.0170 (±0.0014). The distribution of the haplotypes frequencies between the 2 siblings is reported in Fig. 5 (see also supplementary material, S1 and S2) as a complex web of haplotypes composed by a double system of star-like network: 1 star encloses the haplogroup A while the other contain the haplogroup B, separated by 26 mutation. No clear geographical differentiation in the distribution of the haplogroups has been detected.

Figure 5. TCS network of haplotypes recorded in the present study, with indications of only the haplotypes detected at least 5 times. All the rest of haplotypes are reported in the supplementary figure S1 and table S1.
Morphological descriptions
Morphological analysis allowed to identify all adult males (n = 93) and females (n = 13) collected as C. rudolphii s.l. Morphometric features of specimens identified as C. rudolphii A and C. rudolphii B by PCR-RFLP and sequencing are reported in Table 2.
Table 2. Morphometric features of mature male and female specimens of C. rudolphii A and C. rudolphii B from Ph. carbo sinensis from present study

Measurements are in μm unless otherwise stated.
Abbreviations TL, total length; Oe, oesophagus length; IC, intestinal caecum length; VA, ventricular appendix length; NR, distance of nerve ring from anterior end; Ta, tail length; RSp, right spicule length; LRp, left spicule length; Vu distance of vulva from anterior end.
Main morphological details of C. rudolphii s.l. are as follows.
Adults with transversely striated cuticle, more marked at anterior end, forming a conspicuous cephalic collar (Figs 6A,B, 7C,D, 8A,B, 9A); 3 well developed lips; 2 pyriform cephalic papillae on dorsal lip (Figs 6A, 7C), 1 cephalic papilla on each subventral lip (Figs 6B, 8A, 9A,B); interlabia well developed, with large base and bilobed tip (Fig. 8B); excretory pore opening at base of ventral interlabium (Fig. 6A); oesophagus muscular, with small globular ventricle; ventricular appendix directed posteriorly; intestinal caecum 2 to 3 times longer than ventricular appendix, directed anteriorly.

Figure 6. SEM micrographs of C. rudolphii A adult male. (A) Apical view of anterior end showing the dorsal lip (dl) and 2 ventral lips (vl) separate by interlabia, and the excretory pore opening at the base of ventral interlabium (arrow). (B) Lateral view of anterior end. (C) Anterior end with detail of the amphid. (D) Caudal end with everted spicules. (E) Spicules with detail of the spicule tip. (F) Caudal end showing the pattern of post-cloacal papillae.

Figure 7. SEM micrographs of C. rudolphii B adult female. (A) Ventral view of anterior end. (B) Detail of amphid. (C) Dorsal view of anterior end. (D) Apical view of anterior end. (E) Detail of vulva. (F) Caudal end.

Figure 8. SEM micrographs of Contracaecum rudolphii B, adult male. (A) Lateral view of anterior end. (B) Subapical view of anterior end, showing interlabia with bilobed tip. (C) Detail of lip edge. (D) Caudal end showing post cloacal papillae and section of spicules. (E) Caudal end with everted spicules and detail of the spicule tip. (F) Detail of caudal end showing post cloacal papillae (pcp) and phasmid (ph).

Figure 9. SEM micrographs of Contracaecum rudolphii B, adult female. (A) Lateral view of anterior end. (B) Detail of ventral lip. (C) Lateral view of anterior portion. (D) Detail of amphid. (E) Detail of vulva. (F) Caudal end.
Males with tail conical, curved at tip (Figs 6F, 8D), with 27–40 pairs of pre-cloacal papillae, forming 2 subventral lines, 2 pairs of paracloacal papillae, 2 pairs of distal subventral papillae and 2 pairs of distal sublateral papillae, 1 pair of phasmids (Figs 6F, 8D, F); spicules sub-equal, folded, with longitudinal alae and pointed tips (Figs 6D,E, 8D,E).
Females larger than males; vulva around second quarter of body length (Figs 7E, 9E); tail conical (Figs 7F, 9F), with rounded tip. Eggs subspherical.
Discussion
The lack of morphological features useful to discriminate among the C. rudolphii A and C. rudolphii B confirms the need of coupling the traditional parasitological observations with the molecular approach based at least on 2 molecular markers, e.g. ITS rDNA and cox2 mtDNA to reach the correct identification.
All adults collected from the Ph. carbo sinensis sampled in Italy and Israel belong to C. rudolphii s.l. confirming great cormorant as the main definitive host for this complex of species in EU (Amor et al., Reference Amor, Farjallah, Piras, Burreddu, Garippa and Merella2020; Mattiucci et al., Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020; Carmeno et al., Reference Carmeno, Rusconi, Castelli, Prati, Bragoni, Santoro, Postiglione, Sassera and Olivieri2022; Cammilleri et al., Reference Cammilleri, D'Amelio, Ferrantelli, Costa, Buscemi, Castello, Bacchi, Goffredo, Mancini and Cavallero2023) as well as in Israel, where C. rudolphii s.l. had never been described.
The molecular analyses (PCR-RFLP and sequencing) performed on 683 adults from 28 Ph. carbo sinensis (10 from Italy and 18 from Israel) allowed the identification of the sibling species C. rudolphii A and C. rudolphii B, often in mixed infections (Table 1). The co-occurrence of both species in the same bird host has been already reported in other studies (Szostakowska et al., Reference Szostakowska, Myjak and Kur2002; Szostakowska and Fagerholm, Reference Szostakowska and Fagerholm2007; Amor et al., Reference Amor, Farjallah, Piras, Burreddu, Garippa and Merella2020; Mattiucci et al., Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020; Carmeno et al., Reference Carmeno, Rusconi, Castelli, Prati, Bragoni, Santoro, Postiglione, Sassera and Olivieri2022; Cammilleri et al., Reference Cammilleri, D'Amelio, Ferrantelli, Costa, Buscemi, Castello, Bacchi, Goffredo, Mancini and Cavallero2023); similar distributions have been observed also in our study. In Italian specimens, the 2 siblings showed a similar proportion (184 A vs 195 B), while the birds from Israel were more frequently infected with C. rudolphii B (116 A vs 188 B) with only 4 birds out of 18 examined showing a mixed infection.
The phylogenetical analyses of both genetic markers showed a well-supported separation between C. rudolphii A and C. rudolphii B, with C. rudolphii B more closely related to C. rudolphii F than to C. rudolphii A in the cox2 (Amor et al., Reference Amor, Farjallah, Piras, Burreddu, Garippa and Merella2020; Mattiucci et al., Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020; Roca-Geronès et al., Reference Roca-Geronès, Fisa, Montoliu, Casadevall, Tobella, Bas, Palomba and Mattiucci2023), but not in ITS rDNA. Unfortunately, except for C. rudolphii F, no cox2 mtDNA sequences are available for the other siblings (C, D, E) included in C. rudolphii complex to better clarify the relationship among the complex even by this more evolving gene.
Concerning the population structure, we observed a high genetic variability as demonstrated by the high numbers of haplotypes in both C. rudolphii A (83 sequences/60 haplotypes) and C. rudolphii B (58 sequences/46 haplotypes) populations; similar results were obtained also by Amor et al. (Reference Amor, Farjallah, Piras, Burreddu, Garippa and Merella2020) in Sardinia (Italy) for cox2 mtDNA (C. rudolphii A: n. haplotypes 33 out of 158, hd = 0.985, nucleotide diversity = 0.013; C. rudolphii B: nucleotide diversity 7/22, hd = 0.952, nucleotide diversity = 0.021). In Spain, Roca-Geronès et al. (Reference Roca-Geronès, Fisa, Montoliu, Casadevall, Tobella, Bas, Palomba and Mattiucci2023) analysed only C. rudolphii A, that was considered as single population due to the low genetic diversity (n. haplotypes 40/56, hd = 0.969, nucleotide diversity = 0.00681). In the same study, the comparisons with the sequences of C. rudolphii A from 2 Italian areas showed similar values as obtained in the present study (Tyrrhenian: n. haplotypes 80/110, hd = 0.986, nucleotide diversity = 0.00871; western Sardinia: n. haplotypes 28/33, hd = 0.979, nucleotide diversity = 0.0117). The haplotypes diversity in any case reflect the geographical distributions of the sequences analysed (Amor et al., Reference Amor, Farjallah, Piras, Burreddu, Garippa and Merella2020; Roca-Geronès et al., Reference Roca-Geronès, Fisa, Montoliu, Casadevall, Tobella, Bas, Palomba and Mattiucci2023). Amor et al. (Reference Amor, Farjallah, Piras, Burreddu, Garippa and Merella2020) speculate about the low genetic diversity among the 2 siblings, suggesting that the high infection rates in definitive hosts slow down the genetic drift, but also it could be due to the parasites dispersal model mediated by the host dispersal dynamic. The migrations of the definitive hosts and the population structure could influence the genetic structure of this Anisakidae, as reported by Cipriani et al. (Reference Cipriani, Palomba, Giulietti, Marcer, Mazzariol, Santoro, Aco Alburqueque, Covelo, López, Santos, Pierce, Brownlow, Davison, McGovern, Frantzis, Alexiadou, Højgaard, Mikkelsen, Paoletti, Nascetti, Levsen and Mattiucci2022) for Anisakis spp.
The SEM observations allowed for the first time a detailed characterization of the external features of C. rudolphii B, and added further morphological information about C. rudolphii A, which was already examined using SEM by Abollo et al. (Reference Abollo, Gestal and Pascual2001). Particularly, we provide the first detailed SEM characterization of the spicule morphology in both sibling species, with special reference to their folded appearance and to the morphology of the distal extremity.
Overall, compared to other siblings of the species complex (Abollo et al., Reference Abollo, Gestal and Pascual2001; Shamsi et al., Reference Shamsi, Norman, Gasser and Beveridge2009) and to C. rudolphii s.l. (Amato et al., Reference Amato, Monteiro and Amato2006; Li et al., Reference Li, Xu and Zhang2013) previously analysed by SEM, our specimens show a similar external morphology of anterior and posterior extremity. The pattern of post-cloacal papillae in adult males and the conspicuous cuticular collar in both males and females are particularly distinctive characters that support the identification of C. rudolphii s.l.
Generally, cormorants from central Italy showed a similar proportion of adult stages of C. rudolphii A and C. rudolphii B while the ones from Israel showed a higher proportion of C. rudolphii B. It has been suggested that the different feeding ecology and wintering behaviour of different populations of Ph. carbo sinensis, could be one of the ‘drivers’ of the differential spatial distribution of C. rudolphii A and C. rudolphii B in the different aquatic ecosystems, i.e. brackish/marine and freshwater environments, respectively (Mattiucci et al., Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020). Moreover, abiotic factors related to early stages of the parasites, have been supposed to contribute to the differential occurrence of the 2 sibling species in the 2 aquatic ecosystems (Moravec, Reference Moravec2009; Mattiucci et al., Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020).
Recent work suggested the possible role of the migration routes of wintering populations of cormorants in the Mediterranean Sea in influencing the distribution and genetic structure of C. rudolphii (Roca-Geronès et al., Reference Roca-Geronès, Fisa, Montoliu, Casadevall, Tobella, Bas, Palomba and Mattiucci2023). Cormorants migrate from Europe and winter in Israel during November–March, forming large colonies along the Mediterranean and Red Sea coasts and at inland streams and wetlands (Nemtzov, Reference Nemtzov2008). Large numbers of C. rudolphii A could be acquired by cormorants feeding in coastal areas during their migration across the Mediterranean Sea. Furthermore, most of Israel's wetlands are exploited for fish farming, mainly in freshwater fishponds; these fish are particularly susceptible to predation by piscivorous birds during migration stopovers (Nemtzov, Reference Nemtzov2002) and could provide opportunities for maintaining the life cycle of C. rudolphii B. In our study, all the great cormorants from both countries have been sampled in winter; interestingly, in 2 cases the birds from Israel were sampled in an area very close to the sea but they were infected only by C. rudolphii B, therefore we could hypothesize that they arrived in these wintering sites already parasitized. To date, larval stages of C. rudolphii have never been reported in marine/brackish/freshwater fish from Israel. This could either be due to biotic/abiotic factors unfavourable to the development of C. rudolphii larvae, or to difficulties in the detection of small larval stages (much smaller than those of other Contracaecum spp.) during field inspections. Despite their adaptation to different aquatic ecosystems, larvae of both C. rudolphii A and C. rudolphii B have generally shown a low specificity for their fish host (Mattiucci et al., Reference Mattiucci, Sbaraglia, Palomba, Filippi, Paoletti, Cipriani and Nascetti2020), therefore the availability of suitable paratenic hosts does not seem to be a limiting factor for the successful establishment of the parasite life cycle.
This work presents the first data on the occurrence of sibling species of C. rudolphii in Israel and provides additional data on the distribution of C. rudolphii A and B in Italy, confirming the high prevalence and intensity of infection observed in Ph. carbo sinensis from other Italian areas. Based on these results, we stress the importance of carrying out targeted investigations aimed at evaluating the occurrence of C. rudolphii in marine and freshwater fish from Israel, to understand the epidemiology of the parasite in this important wintering stopover.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0031182023000902.
Data availability
The DNA sequences generated in this study have been deposited on the public database GenBank under accession numbers OR263194-OR263246 (ITS rDNA) and OR269666-OR269717 (cox2 mtDNA).
Acknowledgements
The authors thank Professor Roberta Galuppi and Dr Giorgia Bignami of DIMEVET for their help in the collection of samples. They also thank Omer Ben-Asher and Ziv Riklis from the GIS Unit in the Ministry of Agriculture and Rural Development for the preparation of maps.
Author's contributions
M. C. and P. T. wrote the main manuscript text, carried out all the analyses and prepared all the figures. A. C., V. L. and P. A. G. provided the technical support for S. E. M., and molecular analyses. N. D. and S. R. carried out the sampling and necropsies. A. G. and M. L. F. revised the manuscript. All authors reviewed the manuscript and approved the final version.
Financial support
This study was supported by the Israeli Veterinary Services and by the University of Bologna, Italy. Scanning Electron Microscopy acquired thanks to the Department of Excellence Project 2018-2022 funded by the Italian Ministry of Education, Universities and Research.
Competing interest
None.
Ethical standards
Not applicable.














 Open access
Open access