



Recent ecological studies indicate that temperate forests—including those characteristic of western Patagonia—are prime habitat for human foragers (Freeman et al. Reference Freeman, Erick Robinson, Darcy Bird and Anderies2020; Tallavaara et al. Reference Tallavaara, Eronen and Luoto2018). Archaeological evidence suggests, however, that Patagonia's forests were colonized relatively late and only ever occupied sparsely; perhaps they were used seasonally or as necessitated by unfavorable conditions on the adjacent steppe (e.g., Arrigoni Reference Arrigoni1997; Aschero et al. Reference Aschero, Laura Moya and Martínez1995; Belardi et al. Reference Belardi, García, Campán and Borrero1998; Bellelli et al. Reference Bellelli, Mariana Carballido and Scheinsohn2003; Borrero Reference Borrero1994–1995; Borrero et al. Reference Borrero, Mondini, Muñoz and Wickler2004; Goñi Reference Goñi1988; Mena Reference Mena1991, Reference Mena2000; Méndez and Reyes Reference Méndez and Reyes2008; Méndez et al. Reference Méndez, Reyes, Delaunay, Velásquez, Trejo, Hormazábal, Solari and Stern2016; Scheinsohn et al. Reference Scheinsohn, Szumik, Leonardt, Salemme, Santiago, Alvarez, Piana, Vázquez and Mansur2009; Silveira Reference Silveira1987, Reference Silveira, Belardi, Fernández, Goñi, Guráieb and De Nigris1999). This pattern has been explained in terms of foraging efficiency: the largest game species in forests (e.g., huemul deer, Hippocamelus bisulcus) are generally smaller, less gregarious, and more cryptic than their steppe counterparts (e.g., guanaco, Lama guanicoe), making hunting in forests relatively unprofitable (Mena Reference Mena1995; Scheinsohn et al. Reference Scheinsohn, Szumik, Leonardt, Salemme, Santiago, Alvarez, Piana, Vázquez and Mansur2009). Nonetheless, resource diversity is high in forests, and biodiversity is correlated with ecosystem stability (less variable food availability), which decreases subsistence-related risk and potentially makes forests attractive habitats. Correspondingly, some scholars argue that certain of Patagonia's forested environments were permanently inhabited, if only during specific prehistoric periods (Mena Reference Mena, Zangrando, Barberena, Gil, Neme, Giardina, Luna, Otaola, Paulides, Salgán and Tivoli2013; Pérez and Smith Reference Pérez and Smith2007).
Of course, both interpretations could be correct. Forest use likely differed across space and through time, but because Patagonia's wooded environments have not been studied as extensively as those of the steppe or coast, we know relatively little about prehistoric human occupation of southern Andean forests (Méndez et al. Reference Méndez, Reyes, Delaunay, Velásquez, Trejo, Hormazábal, Solari and Stern2016). For example, it remains unclear whether sparse forest records are the result of limited prehistoric use, limited archaeological sampling, or loss of evidence through time (Méndez and Reyes Reference Méndez and Reyes2008). Each newly described forest site, therefore, has much to contribute to our understanding of woodland adaptations in Patagonia.
Within this context, we report data from our recent excavations at the Alero Largo site (Río Ibáñez–6), located in the Aysén Region in Chilean Patagonia. The area surrounding this large rockshelter was forested—at times heavily—prior to historic period fires set to clear the land for animal husbandry. The temperate deciduous forests that today surround the site offer a variety of resources, including tubers, seasonally available fruits and mushrooms, cervids, and small game (e.g., rodents and birds), as well as wood for fuel, tools, and shelters. Río Ibáñez–6's chronological, lithic, archaeofaunal, and macrobotanical records provide evidence that people occupied the Ibáñez Valley earlier than previously documented and that they may have used a wider variety of forest resources than previously understood. From these data we develop hypotheses regarding prehistoric forest use in Aysén that, when more fully tested, will inform our broader understanding of the suitability of forests for human foragers.
Environmental Setting
Alero Largo (600 m asl) is the local name for a site known to archaeologists as Río Ibáñez-6 (RI-6), the sixth site located during the preliminary survey of the Ibáñez River Valley (Bate Reference Bate1970). The Ibáñez River is approximately 80 km long and crosses several ecological zones as it descends from headwaters on the east side of Mount Hudson (an active stratovolcano), trending east–southeast along the southern base of the Cerro Castillo range to its mouth at General Carrera/Buenos Aires Lake (Figure 1). RI-6 is located in the middle Ibáñez Valley (area ≈6,000 km2), which is characterized by glacial topography, including kettle lakes on elevated plains and in the valley bottom, and abrupt relief with hills and ridges rising in all directions (>1,000 m relief).
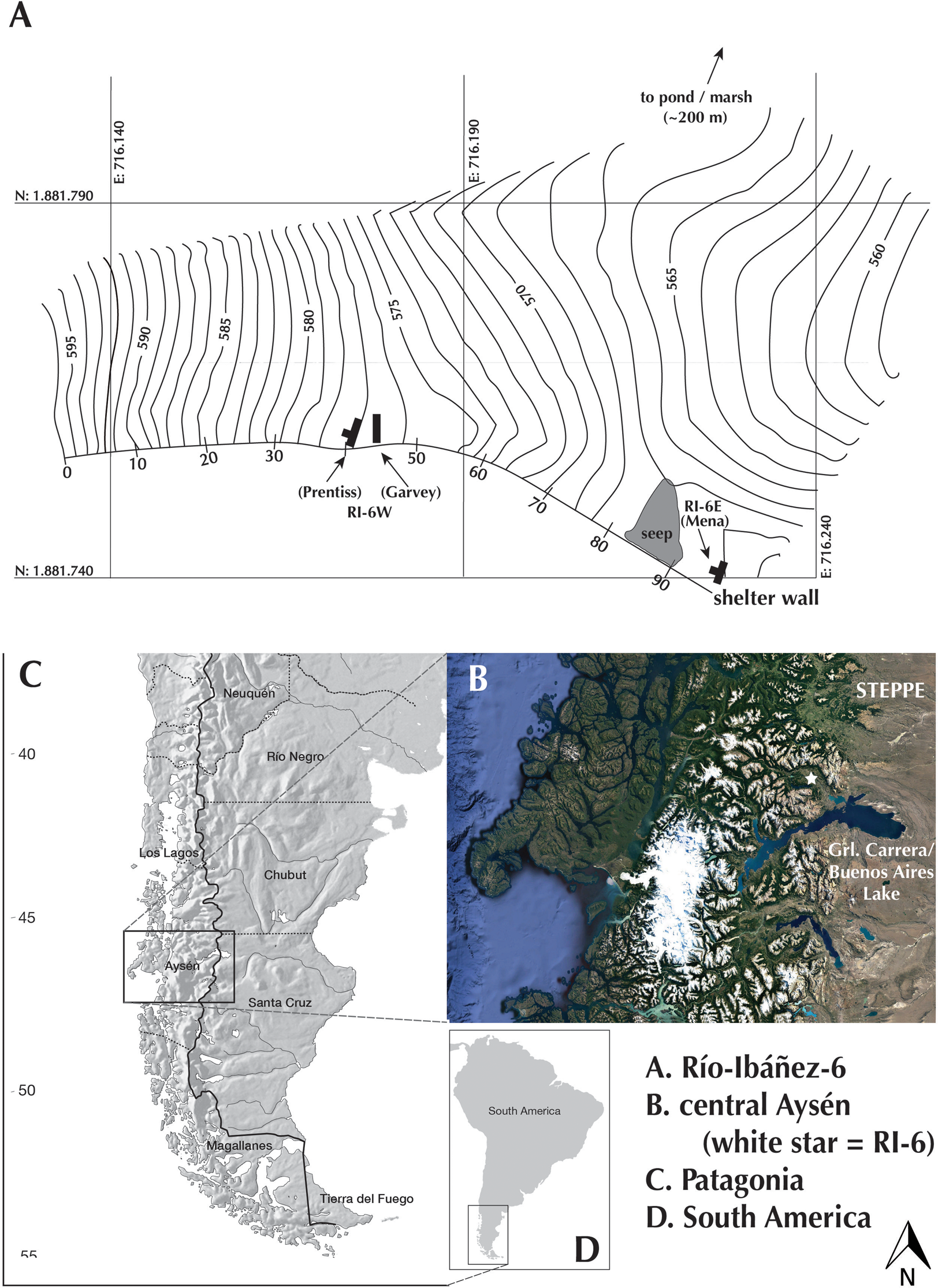
Figure 1. Map of study area. (A) Río Ibáñez-6, indicating the two sectors (west and east), previous excavations and their respective PIs, and location of the pond/marsh mentioned in the text (original by Enrique Valdés Vásquez, modified by Raven Garvey; see also Garvey and Mena Reference Garvey and Mena2016:Figure 1; Prentiss et al. Reference Prentiss, Walsh, Barnett, Murphy and Kuenstle2015:Figure 2); (B) regional geography; (C) Patagonia; and (D) South America.
Today, the region receives ~800 mm of precipitation annually, primarily as snow between May and September. Winds in the valley are frequent and strong. In the immediate vicinity of RI-6, dense lenga (Nothofagus pumilio; beech) forest is now dotted with open, grassy parkland, though forest cover was more complete prior to the devastating fires of the mid-twentieth century (Mena Reference Mena2016a). Huemul dominates archaeofaunal assemblages in the valley; tuco-tuco (a gopher-sized rodent; Ctenomys sp.) and pichi (dwarf armadillo; Zaedyus pichiy) are also common. A freshwater pond (area ≈73,000 m2) and associated marsh are located 300 m north of RI-6. This pond and similar waterbodies in the region are potential sources of fish, fowl, and birds’ eggs, but as with wild plants (Pérez de Micou Reference Pérez de Micou1979–1982), there is virtually no evidence of their prehistoric use in the Ibáñez Valley. However, a recent protein residue analysis tentatively identified weakfish proteins on one stone specimen from the western portion of RI-6 (Garvey and Mena Reference Garvey and Mena2016). We take this up in the discussion section.
Paleoenvironmental studies indicate that Aysén Province was completely covered by ice during the Last Glacial Maximum, and that glacial retreat began around 14,000 cal BP (Clapperton Reference Clapperton1993). There is a well-documented pulse of Nothofagus forest development between about 9000 and 4500 cal BP (Martel-Cea and Maldonado Reference Martel-Cea and Maldonado2010), which coincided with warmer, wetter conditions during the Holocene Climate Optimum (Waldmann et al. Reference Waldmann, Daniel Ariztegui, James Austin, Charles Stern and Dunbar2010), a multi-millennium, global-scale climate event characterized by elevated temperatures (Albanese and Frison Reference Albanese, Frison and Arthur Bettis1995). One of several major eruptions of Mount Hudson (H2) occurred about 3290 cal BP (Weller et al. Reference Weller, Carmen Miranda, Villa-Martínez and Stern2014), at which time there also occurred a pronounced decrease in Nothofagus and a sharp increase in Poaceae (Markgraf et al. Reference Markgraf, Whitlock and Haberle2007). Between 3100 and 1300 cal BP, some of the region's forests retracted (Martel-Cea and Maldonado Reference Martel-Cea and Maldonado2010; Reyes et al. Reference Reyes, Méndez, Maldonado, Velásquez, Trejo, Cárdenas and Abarza2009). The protohistoric plant regime was established by around 1200 cal BP (de Porras et al. Reference de Porras, Maldonado, Abarza, Cárdenas, Francois, Martel-Cea, Stern, Méndez and Reyes2012), followed by deliberate, widespread burning of forests from AD 1910/1920 to the mid-1950s as Criollo/Euro-American settlers cleared land for livestock grazing (Szeicz et al. Reference Szeicz, Zeeb, Bennett and Smol1998).
General Characteristics and Excavation History
At RI-6, cultural materials are concentrated beneath the north-facing overhang of a massive andesitic toba outcrop (Figure 2). The natural shelter is approximately 155 m long, is oriented east–west, and presently affords ~720 m2 of covered space inside the drip line, although evidence suggests that the covered area may once have been substantially larger (Garvey Reference Garvey2016). On the shelter walls there are more than 150 pictographs, which are primarily negative handprints in reddish mineral pigment (Artigas and Muñoz Reference Artigas and Muñoz2015). Efforts to date these pictographs have been largely unsuccessful (Cordero et al. Reference Cordero, Muñoz and Artigas2019), but it is noteworthy that they extend below the modern ground surface.

Figure 2. RI-6. (A) View south toward outcrop/overhang and site; (B) view north from site showing proximity of the pond/marsh. (Photographs courtesy of Raven Garvey.) (Color online)
The site was first archaeologically tested in the context of a 2012 field school jointly directed by Drs. Anna Prentiss (University of Montana) and Francisco Mena (Centro de Investigación en Ecosistemas de la Patagonia). Excavations in two sections of the site, RI-6W and RI-6E (Figure 1), produced modest stone and bone assemblages and, in the west (RI-6W), occupational surfaces with shallow hearth features (Prentiss et al. Reference Prentiss, Walsh and Barnett2012, Reference Prentiss, Walsh, Barnett, Murphy and Kuenstle2015, Reference Prentiss, Barnett and Walsh2016). Huemul and rodents dominate the faunal assemblages, and small amounts of Chenopodium sp. were present in RI-6E (Belmar and Meneses Reference Belmar and Meneses2013). In 2015 and 2016, we excavated four additional 1 × 1 m units at RI-6W, just east of those excavated in 2012 (Figure 1). These more recent excavations deepen our understanding of the site's occupation history and use.
Stratigraphy and Dating
In 2015–2016, four units were excavated to varying depths, determined by productivity (i.e., consecutive sterile levels) and safety; Unit 3 was terminated 310 cm below the surface, and adjacent units were excavated to 150 cmbs to facilitate access and reduce risk (Figure 3). Strata across the four excavation units are generally well defined, and Strata A–E are present in all four units; F–H are present only in Unit 3, owing to its greater depth. Exfoliation of the shelter walls is evident throughout the excavation. Large blocks are most common in units closest to the shelter's exterior, which might account for the microbedding and generally more intricate stratigraphy observed in these units, perhaps as a result of complex interactions between large stone masses and aeolian sediments. There is little evidence of postdepositional disturbance, except for soil coloration consistent with rodent burrowing in the lowest levels of Unit 4 (Figure 3) and an anomalous radiocarbon age (2020 ± 25 BP; UGAMS-24461; charcoal; δ13C = −27.0‰) beneath the H2 tephra (see the later discussion and Supplemental Table 1). A comprehensive study of site formation has not yet been completed, but independent preliminary assessments by two of the site's principal investigators (Garvey Reference Garvey2016; Prentiss et al. Reference Prentiss, Walsh and Barnett2012, Reference Prentiss, Walsh, Barnett, Murphy and Kuenstle2015, Reference Prentiss, Barnett and Walsh2016) suggest that runoff has not significantly affected the site's integrity. Neither do we believe that wind (e.g., natural deposition of macrobotanicals) or wildfires have contributed significantly to the composition or state of the deposits (see the later discussion).

Figure 3. Line drawing of west wall profile showing all four excavation units.
Characteristics of Stratum F (Figure 3) align well with those of a tephra derived from a major eruption of Mount Hudson around 3290 cal BP (H2; Naranjo and Stern Reference Naranjo and Stern1998; Weller et al. Reference Weller, Carmen Miranda, Villa-Martínez and Stern2014). Like the H2 tephra, RI-6W's Stratum F is approximately 10 cm thick, yellowish with a high proportion of angular pumice, and located more than a meter below the modern ground surface (70–120 cmbs)—in this case, it is somewhat deeper than average (~180 cmbs), perhaps because of the differential accumulation of sediments in a shelter occupied by humans versus the lake cores and cut banks throughout the valley on which the average depth is based.
The H2 tephrachronological benchmark is supplemented at RI-6 by a total of nine radiocarbon dates between about 170 and 7400 cal BP (Supplemental Table 1). In the eastern section of the site (RI-6E), the basal date (ca. 6000 cal BP; Mena Reference Mena and Mena2016b) derives from a bone found immediately overlying bedrock. Reaching bedrock in the western section will require significant expansion of the excavation to safely step down more than 3 m below the surface. Accordingly, the earliest date in the western section (6500 ± 30 BP; UGAMS-24462; charcoal; δ13C = −26.4‰; 260 cmbs; see also Supplemental Table 1) may not reflect its earliest use. Indeed, culture-bearing sediments continue for at least 50 cm below the material dated to 6500 ± 30 BP, though no sample suitable for reliable dating was recovered from them.
There are no reversals in the radiocarbon sequence, but as mentioned, one date is anomalous. A charcoal sample collected from Stratum G (248 cmbs; Figure 3) returned a date of 2020 ± 25 BP (referenced earlier; 1941 cal BP, estimated as the median of the calibrated age range at two standard deviations using Calib version 7.1 [Stuvier et al. 2020], and the SHCal13 calibration curve [Hogg et al. Reference Hogg, Hua, Blackwell, Buck, Guilderson, Heaton and Niu2013]). The sample was located roughly 60 cm below the H2 tephra (ca. 3290 cal BP) and just 12 cm above the collection location of the charcoal sample that returned a date of 6500 ± 30 BP (referenced earlier; 7370 cal BP). Two explanations are possible: (1) postdepositional disturbance, although none was evident in the unit from which the samples were collected or during illustration of stratigraphic profiles or scrutiny of the photographic record; and (2) accidental relocation of younger charcoal during excavation. Entering and exiting Unit 3 as it approached 310 cmbs became increasingly difficult, both because of its depth and, at the depth of the anomalous date, the unit's area was constricted due to pedestalling of large rocks in more superficial levels (Figure 3). It is possible that charcoal from higher up in the exposed profile was dislodged as excavators navigated the space. Clarification of this dating anomaly is a priority going forward.
Material Record
Our excavations followed both natural and arbitrary (10 cm) levels to maximize stratigraphic control. We recorded the three-dimensional location of all artifacts larger than 3 cm observed in situ and passed all excavated sediments through 4 mm hardware mesh to recover any additional cultural materials. As described in the section on macrobotanicals, we also collected soil samples for the recovery and identification of microremains.
Lithics
Our excavations produced 441 lithic artifacts (Supplemental Table 2). These were analyzed using an attribute-based classification scheme (after Andrefsky Reference Andrefsky1998), including lithic material type, artifact morphometrics (e.g., length), flake completeness, presence of cortex, and the location and extent of any retouch.
Five primary raw material types were identified macroscopically: cryptocrystalline silicates (75% of lithic assemblage; hereafter CCS), obsidian (10%), nonvitreous volcanic stone (andesite, rhyolite, and basalt; together, 7%), quartzite (2%), and unmodified quartz crystals (see also Supplemental Table 2). With the exception of obsidian, all these types can be found in modest quantities (and varying qualities) within the valley (Gómez Reference Luisa2013). The RI-6W obsidian has not yet been geochemically identified, but Gómez (Reference Luisa2013) reports that obsidian artifacts from other sites in the valley derive from the Pampa del Asador source, located ~500 km away (Stern Reference Stern, Belardi, Fernández, Goñi, Guráieb and De Nigris1999).
Debitage dominates the assemblage (89%), and most is relatively small (11–17 mm maximum dimension). A flake completeness analysis of 40% of the site's debitage shows that the assemblage consists primarily of complete flakes (56%) and proximal fragments (22%). Cortex appears on only six specimens—two flakes of obsidian, three of CCS, and one of an unidentified coarse-grained material. Jointly, these attributes suggest that late-stage tool production or maintenance was the most common knapping task performed on site. The tools themselves (N = 19) are predominantly (74%) formal, including retouched scraper-type unifaces (n = 7) and bifaces (n = 5), most with haft elements (i.e., projectile points, n = 3). There are also four retouched/used flakes, one modified cobble (e.g., grinding stone), and two cores.
Archaeofauna
A total of 3,367 pieces of animal bone were recovered during the 2015–2016 field seasons (Supplemental Table 3). These pieces were quantified following Lyman (Reference Lyman1994) and taxonomically identified using local comparative collections and reference materials. Most (92%) were taxonomically identified, and the assemblage is strongly dominated by Artiodactyla (96%). Within this taxon, finer distinctions are complicated by significant fragmentation, but identified species include Hippocamelus bisulcus (3% of Artiodactyla) and Ovis aries (NISP = 2). Rodentia is the second-most populous taxon (3.5% of identified specimens), 70% of which are Ctenomys sp. (tuco-tuco). Other represented taxa are Zaedyus pichiy (dwarf armadillo, Quirquincho peludo; NISP = 12), Lepus europaeus (European hare; NISP = 1), and Aves (NISP = 4). Domesticated sheep and European hare are exclusive to the uppermost stratum.
Despite high taxonomic identifiability, skeletal parts were less readily determined (17%; N = 579) due to fragmentation. Skeletal part representation nonetheless suggests that whole animals were taken to the site throughout the occupation sequence. Axial (45%) and appendicular (55%) elements are comparably represented, but their fragmentation rates differ: 40% and 70%, respectively. Long bones and phalanges (metapodials) are particularly fragmented, which may reflect processing for marrow. Alternatively, given that 90% of the percussion marks on these bones are located medially where marrow concentrations are not especially high, it is possible that the target was cortical material for tool manufacture. Only two bone tools were recovered from the site; both are awls or punches made on ungulate long bones. Other pieces of bone with signs of anthropic intervention are likewise few (N = 55; <2% of assemblage); these signs include flake scars, percussion and cut marks, and evidence of scraping and drilling. These marks are exclusive to Artiodactyla (Supplemental Table 3).
Of the 130 specimens with characteristics indicative of age at death (all Artiodactyla), 59% are consistent with juveniles between two and four months old (unfused epiphyses) or with subadults (18% have partially fused epiphyses); the remaining 41% are fully fused adults (after Vila et al. Reference Vila, Cristián Saucedo, Ramilo, Corti, Duarte and González2010). There are no clear temporal patterns in these data, and proportions of age classes are largely invariant through time.
In addition to the bone tools mentioned previously, we recovered a possible awl preform, a polished bone flake, and a single bead—a near-perfect circle with a 1.3 mm hole in the center—produced from the shell of an as-yet-unidentified taxon (Figure 4).

Figure 4. Sample of artifacts from RI-6W: (a–b) Projectile point fragments; (c, e–f) unifacial tools; (d) worked cobble; (g–h) bone tools; and (i) bead (figure prepared by John Klausmeyer, University of Michigan Museum of Anthropological Archaeology). (Color online)
Macrobotanical Remains
To recover macrobotanical remains, we analyzed sediments from a column sample extracted from Unit 3 (Figure 3; terminal column depth = 300 cmbs). Macrobotanical remains were not present in the heavy fraction, but the light fraction—viewed under low-power (7–45×) stereo microscopy—produced a modest record (N = 3,444), representing 18 taxa across 12 families (Supplemental Table 4). All identified specimens belong to taxa with known alimentary, medicinal, or technological utility. Most specimens derive from herbaceous plants, but it was rarely possible to determine provenance (endemic, native, or adventitious) because so few are identifiable to species (n = 2), and most of the represented genera/families have broad distributions across the region.
Approximately 20% of the assemblage is thermally altered (n = 802). Only 8% of these are taxonomically identifiable, but more than three-quarters of the unidentified specimens (78%) represent a single taxon, dubbed “Indeterminate-1.” This taxon is concentrated (99%) between 130 and 180 cm below the surface. Taxonomically identified, thermally altered specimens are dominated by Galium sp. (n = 39) and Chenopodium sp. (n = 19). Some charred remains (11%) were not identifiable due to poor preservation, fragmentation, and thermoalteration, which may indicate processing by humans, chemical diagenesis, or both.
Of the uncharred remains (n = 2,642), 96% are taxonomically identifiable and represent 17 taxa across 12 families. Three taxa are adventitious: Trifolium sp. (clover), Erodium sp. (alfilerillo), and Urtica dioica (common nettle); Erodium and Urtica were very likely introduced from Europe. Amaranthaceae (n = 1,024) are second only to Urtica sp. (n = 1,397) in their abundance, and nearly all (93%) of the Amaranthaceae identifiable to genus are Chenopodium sp.
Ceramic
We also recovered a single ceramic sherd, the first found in a stratigraphic context in the Ibáñez Valley. It is an undecorated body sherd with limited diagnostic potential; until we complete special analyses of the specimen, we are unable to interpret its significance or evaluate potential connections to ceramics in neighboring regions.
Temporal Patterns
To facilitate discussion of temporal patterns at RI-6W, we identify three primary chronological blocks: Block 3 predates the H2 eruption (ca. 3290 cal BP) and includes a radiocarbon age of about 7400 cal BP; Block 2 is a midden deposit that postdates the H2 eruption and includes a radiocarbon age of 1630 ± BP (UGAMS-24463; charcoal; δ13C = −26.5‰; 1479 cal BP); and Block 1 includes occupations likely dating to the last 500 years (Figure 3).
Block 3: >3290 cal BP (Strata F–H, ~170–300 cmbs)
Only Unit 3 was excavated to a depth sufficient to expose sediments from this chronological block. Although the associated material record is smaller than other blocks in an absolute sense, it has the highest artifact concentration by excavated volume, suggesting at least comparably frequent or intensive use of the site during the middle Holocene. It bears mentioning that most of the lithics (91%) and bone (98%) associated with this block derive from Stratum G. Unsurprisingly, Stratum F—the H2 tephra—contains very few artifacts, and the scarcity of materials in Stratum H (270+ cmbs) may signal very low-intensity use of the site in the early middle Holocene (>7400 cal BP). Incidentally, our earliest radiocarbon age is shortly after an eruption of Mount Hudson (H1; ca. 7500–7800 cal BP; Stern Reference Stern2008) that was as large and explosive as Krakatoa (AD 1883) or Pinatubo (AD 1991), blanketing much of Patagonia in tens of centimeters of tephra (Weller et al. Reference Weller, Carmen Miranda, Villa-Martínez and Stern2014). Specific effects of this eruption/ashfall event are not well documented in the immediate area, but it is believed to have had a significant impact on Patagonia's human populations (Miotti and Salemme Reference Miotti and Salemme2004; Salemme and Miotti Reference Salemme and Miotti2008). We revisit this in the discussion section.
Consistent with the rest of the stratigraphic sequence, the Block 3 lithic assemblage is dominated by CCS (67%). Nonetheless, this block has both the highest proportion and highest raw count of obsidian in the entire sequence (20%, n = 18). Again, the significance of this find is amplified by the relatively small volume of excavated sediment associated with this chronological block (Figure 3). Obsidian does not occur naturally in the valley, and geochemical analyses will help us better understand material conveyance throughout the region. Regardless of its source, the high proportion of obsidian may indicate greater mobility or broader interaction spheres earlier in the site's occupation than later, when obsidian grows scarce and eventually falls out of the assemblage in Block 1 (cf. Gómez Reference Luisa2013). There are only two chipped-stone tools—a used flake and a knife-type tool of blade proportions struck from a unidirectional core. Four small, smooth pebbles recovered from strata in this block are inconsistent with naturally occurring stones in the immediate area and are of suspected cultural origin, though their significance is presently unknown.
The density of bone by excavated volume is higher in this stratigraphic unit than any other, and four specimens show evidence of cutting and percussive blows; 5% are thermally altered. Conversely, macrobotanical remains are few compared to Blocks 1 and 2 (2% of the assemblage, n = 69) and are concentrated in the uppermost 10 cm level. Most specimens were not taxonomically identifiable, but Galium, Indeterminate 1, and Juncaceae predominate, whereas Amaranthaceae and Chenopodium—which dominate younger deposits—are virtually absent. Nearly half of all macrobotanical remains are thermally altered (47%). It remains unclear whether plants were procured in smaller amounts during these early occupations or whether the seeds’ relative sparsity simply reflects time-dependent attrition.
Block 2: ≤1500 cal BP–circa 3290 cal BP (Stratum E, ~80–170 cmbs)
Block 2 constitutes the culturally richest stratum in the sequence, accounting for 45% of all bone found at the site and 36% of all lithics, including 58% of all stone tools. It contains almost 20% of the site's macrobotanicals, which is significant given the otherwise dramatic decline in plant remains below Block 1's Stratum C (see Figure 3 and the supplemental material).
As in the Block 3 occupations, the full complement of lithic raw materials is represented in Block 2, and the deposit is strongly dominated by CCS (78% of assemblage). Gómez (Reference Luisa2013) has demonstrated that high-quality CCS is available locally as cobbles in river and stream channels, but until we can implement a regionwide geochemical assessment of both geological and archaeological CCS, we cannot verify that the RI-6 materials were obtained in the valley. It is clear, however, that the obsidian (N = 16; 10% of assemblage) originated elsewhere. Although obsidian is less abundant in this block than in earlier deposits, its continued presence may reflect relatively high mobility or interaction with groups from outside the valley (compare with Block 1).
There is a high degree of bone fragmentation, and the skeletal part could not be determined in most (82%) cases. Block 2 contains the most substantial evidence of burning both in the frequency of thermally altered bone (13% of block assemblage) and macrobotanicals (93%) and in the prevalence of ash and carbon spicules in the matrix. Similar evidence of burning at this depth below the surface was not evident in the 21 shovel tests dug between the shelter and adjacent pond in 2015 (Figure 5; Garvey Reference Garvey2016), suggesting contained burns (e.g., hearths) within the site, rather than a widespread burn event (e.g., forest fire). Like the bone assemblage, macrobotanicals in Block 2 (burned and unburned) are highly fragmented, which may reflect extensive plant processing. However, the degree of thermal alteration evident in this stratum could indicate a distinct treatment of refuse, perhaps necessitated by longer or more frequently recurrent stays. This is a hypothesis for future testing.

Figure 5. Plan view map of the 2015 shovel test survey between the rock shelter and adjacent pond.
The plant taxon dubbed “Indeterminate 1” is nearly exclusive to this chronological block (99%), with just seven specimens in other strata (F and G, Block 3); its potential use or uses are unknown. Galium is also concentrated in Block 2 (82%), and members of this genus have both food and medicinal value, as well as documented Patagonian use in the production of red-hued pigments (Nacuzzi and Pérez de Micou Reference Nacuzzi and de Micou1983); this latter use is particularly interesting considering RI-6's reddish pictographs. The appearance in Block 2 of Chenopodium, which is absent from the older Block 3, could simply be associated with an apparent increase in plant procurement, but we present an alternative hypothesis in the discussion section.
The artifact assemblage is consistent with long-term or serial occupation and the performance of multiple tasks. The lithic component, for example, consists not only of hunting accoutrements (two finely made projectile points and a possible preform) but also five heavily retouched scraper-type unifaces and a used cobble exhibiting marginal battering (e.g., hammerstone), facial smoothing, and discoloration (e.g., grinding stone). A bone awl-type tool and a preform of similar morphology likewise contribute to a picture of diverse activities having been performed at the site during this period. Finally, Block 2 produced a small piece of polished bone whose purpose is unknown; it is the site's only shell bead.
Block 1: ≤500 cal BP (Strata A–D, 0-80 cmbs)
Our excavations did not produce a radiocarbon age for this block, but available chronological data suggest that Block 1 represents the period circa 500 cal BP to the present (Prentiss et al. Reference Prentiss, Walsh, Barnett, Murphy and Kuenstle2015, Reference Prentiss, Barnett and Walsh2016): this hypothesis is supported by the first appearance of European plant and animal taxa. This chronological block contains a high volume of cultural material (42% of all bone, 75% of macrobotanicals, and 18% of lithics), but as in Block 3, these are not evenly distributed among strata. Together, Strata C and D account for 86% of the block's bone and stone materials and 50% of its macrobotanicals. The following summary refers to all Block 1 materials jointly, except where useful distinctions between Strata A/B and C/D are specified.
Lithics are relatively abundant in this block, but only 27 specimens were recovered from the most superficial deposits (Strata A/B); nearly half are unmodified quartz crystals of suspected but unconfirmed cultural origin. Modern-era collecting of stone tools may be partly responsible for their scarcity in these strata, because many local people report having personal collections, which we can attest, having inspected some of them. Consistent with other chronological blocks, all lithic raw material types are present in Block 1, dominated by CCS (76% of assemblage). Obsidian is poorly represented (6%) and is only found in Strata C/D. Likewise, the block's six stone tools—a projectile point, a used flake, two heavily retouched scraper-type unifaces, a core, and a river cobble with one worn face—were all recovered from Strata C/D. The tool assemblage also includes a bone awl and one ceramic sherd that, when considered with the stone tools, suggest a wide range of activities were performed at the site during this period.
As in earlier deposits, Block 1 faunal remains are highly fragmented and dominated by artiodactyls; the skeletal part could not be determined in 74% of cases. Introduced species Ovis aries (sheep) and Lepus europeaeus (European hare) enter the record in Strata A/B. These may be unrelated to human subsistence or indicate either use of the shelter by Criollos/Euro-Americans or incorporation of the species into local diets; evidence is currently insufficient to explain these species’ presence. Finally, although there is no unequivocal evidence of fish consumption at the site, our previous identification of fish proteins at RI-6W (Garvey and Mena Reference Garvey and Mena2016) derives from a specimen recovered between 10 and 20 cm below the surface, corresponding to this chronological block.
Perhaps the most noteworthy aspect of the Block 1 assemblage is its macrobotanical record. Nearly 75% of all plant remains recovered from the site are found in these most superficial deposits. Very few (1%) of these remains are thermally altered, in stark contrast to adjacent Block 2, where 93% are burned. Similarly, only 3% are taxonomically unidentifiable due to fragmentation, whereas plant remains in Block 2 are highly fragmented and largely unidentified. There is, nonetheless, considerable overlap in taxonomic representation between the two blocks (Supplemental Table 4). Differences in seed abundance and state (thermal alteration, fragmentation) between Blocks 1 and 2 may reflect different approaches to food preparation or refuse disposal rather than use/disuse.
Plant genus Urtica is concentrated (99.5%) in the upper 70 cm of the soil column (Strata A–C), and a significant portion (72% of all Urtica) is found in Stratum C. The proliferation of Urtica—species of which were introduced from Europe (Weigend and Leubert Reference Weigend and Luebert2009)—may indicate (1) arrival in the valley of Criollos/Euro-Americans in the final decades of the nineteenth and first decades of the twentieth century or (2) invasive colonization of disturbed earth by European species—transported long distances by wind or wildlife—following a major burn event around AD 1450 (Szeicz et al. Reference Szeicz, Zeeb, Bennett and Smol1998). Either scenario would be consistent with our estimate of this soil packet's age. Species of Urtica have many recorded alimentary and medicinal uses that may well have been known to Native inhabitants of RI-6, whether through local experimentation or via Criollos/Euro-Americans.
Coincident with the proliferation of Urtica in Stratum C, we see a dramatic increase in Chenopodium spp., a shade-intolerant genus that readily colonizes disturbed soils. This increase is also consistent with a hypothesized reduction in the valley's dense beech forests due to a burn event. As with Urtica, a “natural” proliferation of Chenopodium does not preclude its use by local people, who were already aware of the plants’ utility if their (thermally altered) presence in lower strata is any indication. According to some sources, all members of the genus have parts edible to humans (e.g., Pardo and Pizarro Reference Pardo and Pizarro2005; PFAF 2015), and Chenopodium saponins may have been an important fishing technology (see the later discussion).
Finally, although the first appearance of Poaceae (grasses) in Stratum C is consistent with our forest-reduction hypothesis, its virtual absence from the record—even in the uppermost excavation level, which reflects the modern, parkland-dotted landscape—supports our interpretation of the macrobotanical record as one largely generated through human action: we would expect a wind-borne seed rain to include substantial amounts of Poaceae.
Discussion
Our data indicate that stone, animal, and possibly plant procurement were broadly similar through the middle and late Holocene. Locally available lithic materials dominate throughout the sequence, indicating a certain level of landscape familiarity from the earliest documented occupation (sensu Borrero Reference Borrero1994–1995). Tool discard rates are low in all periods, suggesting either that stone was highly conserved, or that perishable materials (e.g., wood) constituted a fair fraction of the tool assemblage, or both (for supporting use-wear evidence, see Méndez et al. Reference Méndez, Reyes, Delaunay, Velásquez, Trejo, Hormazábal, Solari and Stern2016; for a discussion of perishable tools, see Garvey Reference Garvey2021; Garvey and Mena Reference Garvey and Mena2016). The bone assemblage is highly fragmented, but among identifiable bones throughout the sequence, ungulates dominate and both adult and subadult animals are represented, as are axial and appendicular elements in roughly equal proportion. Few plant taxa occur exclusively in early or late deposits, again indicating continuity between time periods. Ongoing studies of the region's rock art continue to enrich our understanding of prehistoric use of the Ibáñez Valley (e.g., Artigas and Muñoz Reference Artigas and Muñoz2015; Cordero et al. Reference Cordero, Muñoz and Artigas2019). Presently, however, we see little evidence for a substantive change in use of the site; that is, from subsistence-focused to something better described as social or ritual (contra the lead author's original position; see Garvey and Mena Reference Garvey and Mena2016).
Despite these continuities, there are also interesting dissimilarities among chronological blocks, indicating smaller-scale but important behavioral differences and perhaps reflecting regional-level changes in forest use through time. First, the early record (Block 3) indicates both familiarity with local lithic raw materials and an important influx of nonlocal obsidian, suggesting either relatively high mobility or interregional exchange. Discarded tools are few, consistent with relatively short-term occupations (again with the caveat that stone may not have been the primary tool material). Artiodactyls account for nearly 90% of the Block 3 assemblage, where macrofloral remains are scarce due either to preservation or a relatively low reliance on plant foods. In later occupations, conversely, tool discard rates are higher, tool forms are more varied, and obsidian is absent from the uppermost strata, along with a relatively high frequency of macrofloral remains and a relatively low frequency of artiodactyls (e.g., just 17% in Stratum A).
These trends and their timing appear to be consistent with patterns observed elsewhere in the Ibáñez Valley and greater forested Aysén (e.g., the Cisnes Valley). The middle Holocene record—between about 9500 and 3000 cal BP—is particularly sparse in Aysén, coincident with a period of maximum forest expansion related to the Holocene Climatic Optimum (de Porras et al. Reference de Porras, Maldonado, Abarza, Cárdenas, Francois, Martel-Cea, Stern, Méndez and Reyes2012, Reference de Porras, Maldonado, Quintana, Martel-Cea, Reyes and Méndez2014). At RI-6W, this corresponds to Block 3, which, as just established, is consistent with high mobility and brief, limited-activity stays, despite being richer in cultural materials than similarly aged deposits throughout the Ibáñez Valley. Effective moisture began to decrease around 3000 cal BP, which led both to the opening of Aysén's forest and reduced precipitation on the adjacent steppe (de Porras et al. Reference de Porras, Maldonado, Abarza, Cárdenas, Francois, Martel-Cea, Stern, Méndez and Reyes2012). Méndez and colleagues (Reference Méndez, Reyes, Delaunay, Velásquez, Trejo, Hormazábal, Solari and Stern2016) hypothesize that the resultant decrease in steppe productivity accounts for the coinciding increase in site frequency in Aysen's forests. At RI-6, the chronological block corresponding to this climatic event (Block 2) is a midden deposit reflecting relatively intensive use and varied activities, as well as continued connections to places beyond the valley (e.g., the prevalence of nonlocal obsidian). In both periods, therefore, the RI-6W record can be interpreted in terms of a broad regional pattern of landscape use involving both the forested Andean slopes and adjacent steppe, as has been demonstrated in other parts of Patagonia (Borrero Reference Borrero, Mondini, Muñoz and Wickler2004; Méndez et al. Reference Méndez, Reyes, Delaunay, Velásquez, Trejo, Hormazábal, Solari and Stern2016; Scheinsohn et al. Reference Scheinsohn, Szumik, Leonardt, Salemme, Santiago, Alvarez, Piana, Vázquez and Mansur2009).
Although our data do speak to seasonality, they cannot resolve the question whether use of Aysénian forests was strictly seasonal during the periods represented in Blocks 3 and 2. RI-6W's archaeofaunal and macrobotanical data indicate that the site was routinely—though not necessarily exclusively—occupied during summer, early fall, or both. Subadult artiodactyls are present in all chronological blocks, and among huemul—ecologically, the most likely artiodactyl species in this assemblage—most births occur in November and December (austral summer). Additionally, our macrobotanical data support a spring-summer-fall occupation: granting that some plants’ seeds could be stored through the winter, few of the taxa represented are valued for their seeds (with Chenopodium being a possible exception), suggesting that the plants were simply collected when in seed, which for many temperate plants occurs between late spring and fall.
Finally, patterns evident in RI-6W's most recent deposits may lend support to Mena's (Reference Mena, Zangrando, Barberena, Gil, Neme, Giardina, Luna, Otaola, Paulides, Salgán and Tivoli2013) hypothesis that the valley became a closed system (i.e., not reliant on steppe resources) during late prehistory. Obsidian drops out of the record in the most superficial deposits, which may indicate a disruption of connections to external sources (whether via direct or indirect procurement), and plant use seems to increase dramatically, which is often associated with reduced mobility (e.g., Hitchcock Reference Hitchcock2004 and authors therein). Simultaneously, artiodactyl remains apparently decrease, perhaps caused by the loss of habitat to forest fires (Szeicz et al. Reference Szeicz, Zeeb, Bennett and Smol1998) or overhunting as human use of the valley increased.
On account of their size (relative to guanaco), solitary nature, and cryptic habits, huemul may not have been a key resource for most prehistoric Patagonians (Borrero and Borrazzo Reference Borrero, Borrazzo, Borrero and Borrazzo2011; Méndez et al. Reference Méndez, Reyes, Delaunay, Velásquez, Trejo, Hormazábal, Solari and Stern2016). However, if small groups of hunter-gatherers did come to inhabit the Ibáñez Valley on a full-time basis in late prehistory (Mena Reference Mena, Zangrando, Barberena, Gil, Neme, Giardina, Luna, Otaola, Paulides, Salgán and Tivoli2013), huemul's importance would likely have increased. In fact, both subsistence models (e.g., Bettinger et al. Reference Bettinger, Garvey and Tushingham2015) and ethnographic and archaeological evidence (e.g., Broughton et al. Reference Broughton, Cannon, Bayham and Byers2011) indicate that, based on body size, huemul would have been the highest-ranking species in the Ibáñez Valley during that time. Subsistence models also indicate that a decline in high-ranking huemul—as is suggested by RI-6W's archaeofaunal record—is often overcome through resource diversification (increased dietary breadth). In most places and times, plant foods are considered among the lowest-ranking resources because of their high processing times (Bettinger et al. Reference Bettinger, Garvey and Tushingham2015). So, in addition to greater reliance on plant foods at RI-6W, we would expect an increase in small game and fish to meet daily caloric—and protein and fat—requirements.
Despite the presence of numerous lakes and ponds, as well as the notable concentration of archaeological sites in the lakes region of the Ibáñez River's middle course, there is little evidence of prehistoric fish exploitation in the valley. In fact, the only evidence currently comes from RI-6W, where we analyzed stone specimens for protein residues via crossover immunoelectrophoresis (Garvey and Mena Reference Garvey and Mena2016). The analysis indicates the possible presence of proteins from a member of the Perciformes order, likely Percichthyidae (temperate perches) or Perciliidae (southern basses). The stone specimen that produced a reaction to Perciformes may not have been anthropically modified (Garvey and Mena Reference Garvey and Mena2016; cf. Prentiss et al. Reference Prentiss, Walsh, Barnett, Murphy and Kuenstle2015, Reference Prentiss, Barnett and Walsh2016), and even though the reaction was replicated, it is a “questionable positive”—indicating that the stone specimen “may have been used to process [fish], but that the proteins are degraded” (Garvey and Mena Reference Garvey and Mena2016:10). Nonetheless, the result is compelling, if difficult to interpret in the absence of fish bone and fishing gear from RI-6W and other sites in the valley. Still, we hypothesize that a closed-valley system (sensu Mena Reference Mena, Zangrando, Barberena, Gil, Neme, Giardina, Luna, Otaola, Paulides, Salgán and Tivoli2013) may have required fishing and that even a rich prehistoric fishing economy could be virtually invisible.
As described earlier, Amaranthaceae is the second-most abundant taxon in our macrofloral assemblage, an overwhelming majority of which belongs to the genus Chenopodium, which strongly dominates the uppermost strata of Block 1. A chemical compound with multiple uses—saponin—is concentrated in Chenopodium roots and bran (Dong et al. Reference Dong, Xiushi Yang, Fenxiang Zhang and Xue2020; Lim et al. Reference Lim, Park and Yoon2019). Relevant to this discussion, saponin-rich plants have a well-known ethnohistorical use in fishing. In California, for example, many tribes (e.g., Luiseño, Miwok, Yokut) introduced pulverized soaproot (Chlorogalum pomeridianum)—also high in saponin—into waterbodies to stupefy fish, which could then be gathered easily without the use of any durable fishing technology (Campbell Reference Campbell1999). In the Ibáñez Valley, fish could have been gutted and boned at the source (e.g., the freshwater pond a few hundred meters away), further reducing the likelihood of fishing-related evidence. Alternatively, fish bones may have been consumed, providing necessary dietary calcium. There is extensive ethnographic documentation of fish bone consumption (Jarvenpa and Brumbach Reference Jarvenpa and Brumbach2006; Kroeber and Barrett Reference Kroeber and Barrett1960; Raymond and Sobel Reference Raymond and Sobel1990; Sepez Reference Sepez2001; Sinclair Reference Sinclair1953; Steward Reference Steward1943; Stewart Reference Stewart1977; for a comprehensive list, see also Morin Reference Morin2020), and numerous examples of fish bones in archaeological human excrement (e.g., Callen Reference Callen1965; Heizer and Napton Reference Heizer and Napton1969; Sobolik Reference Sobolik1991).
RI-6W's conspicuous lack of eggshell, noted by Prentiss and colleagues (Reference Prentiss, Walsh, Barnett, Murphy and Kuenstle2015, Reference Prentiss, Barnett and Walsh2016), could be explained similarly. Like fish bone, eggshell may have been comminuted and consumed as a source of dietary calcium, a common modern practice with a deep history evidenced by studies of archaeological human coprolites (e.g., Callen Reference Callen1965; Heizer and Napton Reference Heizer and Napton1969; Stiger Reference Stiger1977). These hypotheses require further testing but could, if supported, simultaneously explain the prevalence of Chenopodium in the RI-6 macrobotanical sample and the lack of fish bone (and eggshell) in local faunal assemblages. It could also clarify late prehistoric use of the Ibáñez Valley if, in fact, populations did become permanent forest residents at this time.
Final Considerations
RI-6W's material record provides supporting evidence for established patterns of prehistoric forest use while also stimulating new hypotheses—particularly regarding late prehistoric occupation of the Ibáñez Valley, as described earlier, but also related to first use of Aysen's forested environments. Prior to our excavations, the earliest dates in the region were registered at Fontana rockshelter and Las Guanacas cave in the Ibáñez Valley (Mena Reference Mena1983, Reference Mena2000) and Las Quemas rockshelter in the Cisnes Valley (Méndez et al. Reference Méndez, Reyes, Delaunay, Velásquez, Trejo, Hormazábal, Solari and Stern2016), all between 5990 and 6270 cal BP (RI-6E is just slightly younger, at 5975 cal BP; Mena Reference Mena and Mena2016b). The earliest dated occupation at RI-6W is more than a millennium earlier, at about 7400 BP. It remains unclear whether RI-6W is indeed an outlier or whether the current lack of similarly aged deposits elsewhere in the region is better explained by incomplete archaeological sampling, poor preservation in forested environments, or both (Méndez and Reyes Reference Méndez and Reyes2008). Regardless, understanding the circumstances under which RI-6W was first occupied can help us better understand conditions that impel forest use more generally.
The middle Holocene was a time of forest expansion (de Porras et al. Reference de Porras, Maldonado, Abarza, Cárdenas, Francois, Martel-Cea, Stern, Méndez and Reyes2012). If woodlands were not the preferred habitat for prehistoric foragers (Borrero Reference Borrero, Silverman and Isbell2008; Mena Reference Mena1995), an initial occupation during peak forest density requires explanation. Data from RI-6W and other regional sites are currently insufficient to resolve this issue, but a well-documented natural disaster provides a foundation for future research. RI-6W is just ~65 km downwind of Mount Hudson whose H1 eruption around 7500–7800 cal BP deposited tens of centimeters of tephra as far south as Tierra del Fuego (Stern Reference Stern2008). Going forward, pollen coring in the pond adjacent to RI-6W will help determine whether, immediately adjacent to RI-6W, forest growth was halted—reversed even—following the H1 eruption. If so, we might hypothesize that forest denudation actually improved human foraging returns, expanding the ranges of large-bodied, gregarious game (guanaco) into what is today beech forest but was, after the eruption, effectively an extension of the steppe.
Finally, to address the apparent disjunction between Aysén's sparse forest record and recent ecological work indicating the high suitability of temperate forests for human foragers (Freeman et al. Reference Freeman, Erick Robinson, Darcy Bird and Anderies2020; Tallavaara et al. Reference Tallavaara, Eronen and Luoto2018), we advocate the continued coupling of explicit hypotheses and expectations with extensive fieldwork, including archaeological survey and excavation, and local paleoenvironmental reconstruction. We argue, too, for continued exploration of the complementary use of the forest and steppe and for paying particular attention to circumstances that might have fueled seasonal forest use by steppe dwellers (e.g., pushed in by deleterious steppe conditions vs. drawn in by seasonal forest diversity/abundance such as fruits, mushrooms, or beech masts; Garvey Reference Garvey2021). It is only by these means that we will better understand forest use—in southern Patagonia, and other world regions.
Acknowledgments
This work was conducted under permits CMN No 6388 (2015–2016) from the Consejo Monumentos Nacionales de Chile. We thank the community of Cerro Castillo for their hospitality and for allowing us to share in their enthusiasm regarding the region's prehistory. We are very grateful to Don Lino Moreira for generously providing access to the Alero Largo site. The Municipality of Cerro Castillo permitted our use of the Research Station at Escuela Vieja. Simón Sierralta, María Luisa Gómez, and Renata Gutiérrez participated in site excavations. The Centro de Investigación en Ecosistemas de la Patagonia provided research space and temporary artifact storage, and the Museo Regional de Coyhaique provides long-term care and storage of the collections. Funding (to RG) was provided by the Regents of the University of Michigan. Francisco Mena provided help and advice through various stages of this project. Lastly, we thank three anonymous reviewers for their insightful feedback, which helped us improve the article.
Data Availability Statement
All excavated archaeological materials used in this study are curated at the Museo Regional de Aysén, Coyhaique, Chile.
Competing Interests
The authors declare no competing interests.
Supplemental Material
For supplemental material accompanying this article, visit https://doi.org/10.1017/laq.2022.18.
Supplemental Table 1. Radiocarbon Dates and Chronological Blocks.
Supplemental Table 2. Lithic Data.
Supplemental Table 3. Archaeofaunal Data.
Supplemental Table 4. Macrobotanical Data.







 Open access
Open access