


By now, non-alcoholic fatty liver disease (NAFLD) is accounted to be among the most common liver disease worldwide( Reference Blachier, Leleu and Peck-Radosavljevic 1 ). Ranging from simple hepatic steatosis without inflammation to non-alcoholic steatohepatitis, fibrosis and to even cirrhosis and hepatocellular carcinoma, NAFLD encompasses a wide spectrum of disease stages( Reference Hashimoto, Tokushige and Ludwig 2 , Reference Tilg and Moschen 3 ). Results of our group but also of others suggest that besides a general overnutrition and genetic predisposition, alterations at the level of gut, for example alterations of intestinal microbiota and intestinal barrier function subsequently leading to an increased permeation of bacterial endotoxin and elevated endotoxin levels, may be critical factors in the development and progression of NAFLD( Reference Dyson and Day 4 – Reference Volynets, Kuper and Strahl 7 ). However, despite intensive research efforts, molecular mechanisms involved in the development and progression of NAFLD remain unclear and except life-style modifications such as energy-restricted diets and increase of physical activity, efficient treatment and prevention strategies are still lacking.
In humans, the SCFA butyrate, shown to be essential for nourishing intestinal epithelial cells, is mainly produced by intestinal micro-organisms( Reference Berni, Di and Leone 8 ). Furthermore, animal studies suggest that butyrate and micro-organisms synthesising butyrate possess beneficial effects on the development of liver injuries of various aetiologies through improvement of intestinal barrier function( Reference Endo, Niioka and Kobayashi 9 , Reference Liu, Qian and Wang 10 ). In line with these findings, results of Mattace et al. ( Reference Mattace, Simeoli and Russo 11 ) suggest that sodium butyrate (SoB) and its synthetic derivative N-(1-carbamoyl-2-phenyl-ethyl) butyramide may attenuate liver steatosis and inflammation in rats fed a high-fat diet. In addition, results of recent animal studies indicate that treatment with SoB may improve intestinal permeability and reduce serum endotoxin levels, as well as inflammatory cytokines, shown to be involved in the development of liver diseases induced through ingestion of toxins and certain diets, respectively( Reference Endo, Niioka and Kobayashi 9 , Reference Yang, Wang and Li 12 , Reference Wang, Wang and Wang 13 ). Our own group recently showed that oral ingestion of SoB markedly protected mice from the development of a Western-style diet (WSD)-induced NAFLD( Reference Jin, Sellmann and Engstler 14 ). In this study, protective effects were not associated with a protection against the loss of tight-junction proteins in the small intestine but rather with an attenuation of lipid peroxidation and protection against the induction of inducible nitric oxide synthase (iNOS) in livers of mice( Reference Jin, Sellmann and Engstler 14 ). However, molecular mechanisms involved in the beneficial effects of the oral supplementation of SoB are not yet fully understood.
Using a mouse model of feeding a fructose-enriched diet to induce early stages of NAFLD but also differentiated human carcinoma colon-2 (Caco-2) cells as a model of enterocytes and J774A.1 cells as a model of hepatic Kupffer cells, the aim of the present study was to further delineate molecular mechanism involved in the protective effects of an oral supplementation of SoB on the development of NAFLD.
Methods
Animals and treatments
In total, 12-week-old male C57BL/6J mice (Janvier SAS) were housed in a pathogen-free barrier facility accredited by the Association for Assessment and Accreditation of Laboratory Animal Care. All the procedures were approved by the local Institutional for Animal Care and Use Committee. During a 6-week feeding period, mice (n 6/group) were pair-fed (isoenergetic) either a fructose-enriched liquid diet (55 % of total energy intake from fructose, FD) or a fructose-enriched diet supplemented with 0·6 g/kg body weight (Sigma-Aldrich). Isoenergetic pair-feeding of mice in the fructose groups was achieved as detailed in Sellmann et al. ( Reference Sellmann, Jin and Degen 15 ). In brief, intake of the liquid diets was assessed daily within groups, and amount of diet fed was adjusted accordingly among groups for the next day. In addition, naïve mice serving as controls were fed standard pellet chow (Ssniff). Details of diets fed are shown in the online Supplementary Table S1. All mice had free access to plain tap water, and body weight of mice was measured weekly. In week 4, mice were fasted for 6 h and blood samples were collected from the retrobulbar venous plexus to determine fasting blood glucose and insulin levels (Hölzel diagnostics). Homoeostasis model assessment for insulin resistance (HOMA-IR) index was calculated using the formula HOMA-IR=fasting blood glucose (mg/dl)×fasting insulin concentration (mU/l)/405, as previously detailed( Reference Sellmann, Jin and Engstler 16 ). After 6 weeks, mice were anaesthetised with 100 mg ketamin/kg and 16 mg xylazin/kg body weight by intraperitoneal injection. Blood from portal vein was collected just before they were killed. Tissue samples of liver and intestine were either fixed in O.C.T medium (Medite), neutral-buffered formalin or shock-frozen in liquid N2. Samples were then kept in a −80°C freezer until further measurements.
Histological evaluation of liver and goblet cell staining in duodenum sections
Frozen liver sections (10 µm) were stained with Oil red O (Sigma-Aldrich), as described previously( Reference Spruss, Kanuri and Stahl 17 ). Liver sections (5 µm) embedded in paraffin were stained with haematoxylin–eosin (Sigma-Aldrich) to evaluate the histologic features as detailed before( Reference Jin, Sellmann and Engstler 14 ). To determine goblet cell number being indicative of mucous formation( Reference Pelaseyed, Bergstrom and Gustafsson 18 ), duodenal sections (4 µm) embedded in paraffin were consecutively stained with Alcian blue (30 min), periodic acid (10 min), Schiff’s reagent (15 min) (all from Carl-Roth) and counterstained with haematoxylin. Quantitative evaluation of staining was carried out as previously described in detail( Reference Bergheim, Guo and Davis 19 ). Representative photomicrographs of Oil Red O (100×) and haematoxylin–eosin (200×), as well as goblet cell (200×) staining, were captured using the system incorporated in the microscope (Leica DM4000 B LED).
Hepatic TAG determination and clinical chemistry
Hepatic TAG were isolated as described in detail previously( Reference Spruss, Kanuri and Stahl 17 ). TAG levels were determined by using a commercially available kit (Randox). Fasting blood glucose was determined using a blood glucose meter (Contour; Bayer Vital GmbH). Alanine aminotransferase (ALT) activity was determined using a commercially available kit (Randox).
Endotoxin assay
Portal plasma samples were heated at 70°C for 20 min before the measurement. Levels of endotoxin were then determined using a commercially available limulus amebocyte lysate assay with a concentration range of 0·15–1·2 EU/ml (Charles River), as previously described in detail( Reference Haub, Kanuri and Volynets 20 ).
Immnohistochemical staining for liver and duodenum of mice
Paraffin-embedded liver sections (5 µm) were stained for 4-hydroxynonenal (4-HNE) protein adducts and iNOS using a polyclonal antibody (4-HNE: AG Scientific; iNOS: Affinity BioReagents), as described previously( Reference Spruss, Kanuri and Stahl 17 ). Paraffin-embedded duodenal sections (4 µm) were stained for occludin (Invitrogen Corporation), zonula occludens 1 (ZO-1; Life Technologies GmbH) and hydroxyindole-O-methyltransferase (HIOMT; Biozol Diagnostica Vertrieb GmbH) using polyclonal primary antibodies, as described previously( Reference Spruss, Kanuri and Stahl 17 ). In brief, specific binding of primary antibody to the target protein was detected by incubating sections with a peroxidase-linked secondary antibody and diaminobenzidine (Peroxidase Envision Kit; Dako). Photos of eight fields of each tissue section (200× of each liver tissue section; 400× of occludin staining and 200× of HIOMT staining of duodenum tissue) were captured and the extent of staining in the sections was defined as percentage of microscopic field within the default colour range determined by the analysis system incorporated in the microscope (Leica DM4000 B LED). Mean values of eight sections were used as the score of the sample.
RNA isolation and real-time RT-PCR
Total RNA from liver tissue, differentiated Caco-2 cells and J774A.1 cells was isolated using peqGOLD TriFastTM (PEQLAB), and real-time RT-PCR was carried out as detailed by Jin et al.
(
Reference Jin, Engstler and Ziegenhardt
21
) and Wagnerberger et al.
(
Reference Wagnerberger, Spruss and Kanuri
22
). PCR primers for determining melatonin receptor (MT) 2, HIOMT, 18S in human cells, for example Caco-2 cells and PCR primers used to measure expression of chemokine (C–C motif) ligand 2 (CCL-2), toll-like receptor 4 (TLR-4), myeloid differentiation primary response gene 88 (MyD88), insulin receptor (IR), insulin receptor substrate 1 (IRS-1), iNOS, lipopolysaccharide-binding protein (LBP), MT-1 and MT-2, as well as 18S in mouse tissue and cell lines, were designed using Primer 3 software (Whitehead Institute for Biomedical Research). Primer sequences are shown in Table 1. SYBR Green® Supermix (Agilent Technologies) was used to prepare the PCR mix. The comparative cycle threshold (C
t
) method was used to determine the amount of target, normalised to an endogenous reference and relative to a calibrator (
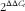 $2^{{\Delta \Delta C_{t} }} $
). The purity of the PCR products was verified by melting curves and gel electrophoresis.
$2^{{\Delta \Delta C_{t} }} $
). The purity of the PCR products was verified by melting curves and gel electrophoresis.
Table 1 Mouse and human primer sequences used for real-time RT-PCR detection

CCL-2, chemokine (C–C motif) ligand 2; TLR-4, toll-like receptor 4; MyD88, myeloid differentiation primary response gene 88; IR, insulin receptor; IRS-1, insulin receptor substrate 1; iNOS, inducible nitric oxide synthase; MT, melatonin receptor; LBP, lipopolysaccharide-binding protein; HIOMT, hydroxyindole-O-methyltransferase.
ELISA
Concentrations of TNF-α, plasminogen activator inhibitor-1 (PAI-1) and superoxide dismutase (SOD)-1 activity in the liver tissue were determined using commercially available kits (TNF-α: AssayPro; PAI-1: LOXO GmbH; SOD activity: Enzo Life Science). Levels of melatonin in duodenum and in cell culture medium were determined with the ELISA kit (IBL International GmbH).
Sodium butyrate determination in mouse plasma
Plasma (15 µl), 15 µl of the internal standard n-valeric acid (0·010 mg/ml in water; Sigma-Aldrich) and 300 µl of n-propanol with 1 % H2SO4 (both from BASF) were combined in a 6-ml tube and incubated at 80°C for 1 h. n-Hexane (500 µl; Th.Geyer) and H2O (3 ml) were added and the tube was shaken. After phase separation, the lower water phase was removed, and the organic phase was washed with 3 ml of H2O. An aliquot (200 µl) was amended with the second internal standard butyric acid butyl ester and the sample was analysed by GC with electron ionisation MS operated in selected ion monitoring mode using an Rtx-2330 column (60 m, 10 % cyanopropylphenyl and 90 % biscyanopropyl polysiloxane; Restek; internal diameter: 0·25 mm; film thickness: 0·1 µm) and helium (quality: 5·0; Sauerstoffwerk) as the carrier gas. After a solvent delay of 7 min m/z 71, m/z 85, m/z 89, m/z 103, m/z 130 and m/z 144 were analysed.
Detection of nitrite
Nitrite content in the cell culture medium was determined using a commercially available colorimetric assay (Griess Reagent System, Promega). In brief, 50 µl of each experimental cell culture medium was mixed with 100 µl of Griess reagent and absorbance was measured at 550 nm wavelength.
Cell culture and treatment
Caco-2 cells (American Type Culture Collection) were maintained in Dulbecco’s modified Eagle’s medium (DMEM; PAN-Biotech GmbH) and cultured as detailed previously( Reference Haub, Kanuri and Volynets 20 ). For experiments, cells were seeded in six-well plates, and after reaching confluence they were differentiated for 9 d. On the last day of differentiation, medium was removed and replaced with fresh medium with SoB (0–6 mm, Sigma-Aldrich) for 6–48 h. Concentrations and time points of treatment were adopted from others( Reference Nazih, Nazih-Sanderson and Krempf 23 ). To determine the melatonin concentration, cell culture medium was collected. For measurement of HIOMT mRNA expression, RNA from cells was extracted as detailed in the online Supplementary Materials.
Cultured J774A.1 macrophages (mouse ascites macrophages; American Type Culture Collection) plated in six-well plates were maintained in DMEM medium supplemented with 10 % fetal calf serum and 1 % penicillin/streptomycin until 70 % confluence was reached. Cells were preincubated with 0–1 mm-melatonin (Sigma-Aldrich) at 37°C for 2 h. Melatonin concentrations used were determined in pilot dose–response studies (data not shown). Then, 25 ng/ml lipopolysaccharide (LPS; Sigma-Aldrich) was added to the medium and incubation was continued for 18 h. Medium was collected to determine nitrite concentration and cells were lysed to obtain RNA for further analysis.
Statistical analysis
Results are expressed as mean values with their standard errors. Non-parametric t test was used to determine statistical differences between the two fructose-diet-treated mouse groups. One-way ANOVA with Tukey’s post hoc test was used to determine the statistical differences when comparing results of cell culture experiments (GraphPad Prism version 6.0; Graph Pad Software). Outliers were identified using the Grubbs’s test. P value<0·05 was considered to be significant.
Results
Plasma butyrate levels, markers of hepatic fat accumulation and inflammation, as well as glucose metabolism
Plasma butyrate levels determined in portal plasma were similar between both FD-fed mouse groups and naïve controls (Table 2). Energy intake and body weight gain, as well as liver weight and liver:body weight ratio, were also similar between FD-fed groups (Table 2). Fasting blood glucose and insulin levels, as well as HOMA index of mice, did not differ between FD-fed groups (Table 2). However, mRNA expression of both IR and IRS-1 was significantly lower in livers of mice fed FD+SoB than in those only fed the FD, which were almost at the level of naïve controls (Fig. 1). Hepatic fat accumulation as determined by assessing liver histology and measuring TAG levels in liver tissue was significantly lower in mice fed FD+SoB when compared with mice only fed FD (TAG: −40 % when compared with FD, P<0·05). However, TAG levels in livers of mice fed FD+SoB were still markedly higher than in naïve controls (Fig. 1(a)–(c)). ALT plasma activity was also markedly lower in mice fed FD+SoB when compared with mice only fed FD; however, as some samples were lost because of haemolysis and as data varied considerably within groups, differences did not reach the level of significance (Fig. 1(d)). Expression levels of CCL-2 mRNA and protein levels of TNF-α and PAI-1 were lower in livers of mice fed FD+SoB when compared with mice only fed FD (Table 2) (CCL-2: approximately −60 %, P<0·05; TNF-α: approximately −30 %, P<0·05; PAI-1: approximately −60 %, P=0·08, all compared with FD). Indeed, concentrations of all three inflammatory markers were almost at the level of those determined in naïve controls in livers of FD+SoB-fed mice.

Fig. 1 Effect of a chronic supplementation of sodium butyrate (SoB) on lipid accumulation and markers of inflammation in livers of fructose-fed mice. Representative photomicrographs of (a) Oil red O staining (100×) and (b) haematoxylin–eosin staining (200×) of liver sections. Red colour in Oil red O staining indicates fat, whereas in haematoxylin–eosin stained tissue sections fat is displayed as ‘white’ droplets, as fat dissolved during embedding of tissue. (c) Quantitative analysis of hepatic TAG content. (d) Alanine aminotransferase (ALT) activity in plasma. Expression of (e) insulin receptor (IR) and (f) insulin receptor substrate (IRS)-1 mRNA in liver tissue of mice. Values are means with standard errors represented by vertical bars. FD, fructose-enriched liquid diet. *P<0·05 between FD- and FD+SoB-fed groups as determined using t test.
Table 2 Effect of an oral supplementation of sodium butyrate on body weight, liver:body weight ratio and markers of liver damage, as well as markers of insulin resistance, in mice fed a fructose-enriched diet for 6 weeks (Mean values with their standard errors)

FD, fructose-enriched liquid diet; SoB, sodium butyrate; CCL-2, chemokine (C–C motif) ligand 2; HOMA-IR, homoeostasis model assessment of insulin resistance; PAI-1, plasminogen activator inhibitor 1.
* Mean value was analysed by t test significantly different from that of the FD group: P<0·05.
Tight-junction proteins and goblet cells in duodenum and toll-like receptor 4 and myeloid differentiation primary response gene 88 mRNA expression, as well as markers of lipid peroxidation and oxidative defence in liver
Protein levels of the tight-junction proteins occludin and ZO-1, as well as numbers of goblet cells in duodenum, did not differ between the two FD-fed mouse groups. However, protein levels of both occludin and ZO-1 in the duodenum were markedly lower in both FD-fed groups than in naïve controls (Fig. 2 and online Supplementary Fig. S1(a)–(c)). In line with these findings, the endotoxin concentration in portal plasma and mRNA expression of LBP in liver also did not differ between FD-fed groups but were markedly higher than in naïve controls (Fig. 3(a) and (b)). However, expression levels of both TLR-4 and MyD88 mRNA were significantly lower in livers of mice fed FD+SoB in comparison with mice only fed FD (Fig. 3(c) and (d)). In line with these findings, protein expression levels of iNOS and 4-HNE protein adducts were also significantly lower in livers of mice fed FD+SoB when compared with those of mice only fed FD (Fig. 4(a) and (b) and online Supplementary Fig. S2). In contrast, activity of SOD-1 was significantly lower in livers of mice fed FD+SoB when compared with mice only fed FD (approximately −50 %) (Fig. 4(c)).

Fig. 2 Effect of a chronic supplementation of sodium butyrate (SoB) on markers of intestinal barrier function. Densitometric analysis of (a) occludin and (b) zonula occludens 1 (ZO-1) protein staining in the duodenum. (c) Number of goblet cells in the duodenum per microscopic field. Values are means with standard errors represented by vertical bars. FD, fructose-enriched liquid diet.

Fig. 3 Effect of a chronic supplementation of sodium butyrate (SoB) on endotoxin levels in portal plasma and mRNA expression of lipopolysaccharide-binding protein (LBP), toll-like receptor 4 (TLR-4) and myeloid differentiation primary response 88 (MyD88) in liver tissue of mice. (a) Endotoxin concentration in portal plasma. Expression of (b) LBP, (c) TLR-4 and (d) MyD88 mRNA in livers of mice. Values are means with standard errors represented by vertical bars. FD, fructose-enriched liquid diet. *P<0·05 between FD- and FD+SoB-fed groups as determined using t test.

Fig. 4 Effect of a chronic supplementation of sodium butyrate (SoB) on markers of lipid peroxidation and oxidative stress in the liver tissue of mice. Densitometric analysis of immunostaining of (a) inducible nitric oxide synthase (iNOS) and (b) 4-hydroxynonenal (4-HNE) protein adducts in liver. (c) Superoxide dismutase-1 (SOD-1) activity in the liver tissue of mice. Values are means with standard errors represented by vertical bars. FD, fructose-enriched liquid diet. *P<0·05 between FD- and FD+SoB-fed groups as determined using t test.
Protein concentration of hydroxyindole-O-methyltransferase and melatonin in duodenum, as well as expression of melatonin receptor-1 mRNA in liver
Protein levels of HIOMT in duodenum of mice fed FD+SoB were significantly higher than those of mice fed only FD (Fig. 5(a) and online Supplementary Fig. S1(d)). In line with the HIOMT protein levels, concentration of melatonin in duodenum was also significantly higher in duodenum of mice fed FD+SoB when compared with mice only fed FD (Fig. 5(b)). Furthermore, MT-1 mRNA expression in livers of mice fed FD+SoB was significantly higher when compared with those only fed FD (Fig. 5(c)).

Fig. 5 Effect of chronic supplementation of sodium butyrate (SoB) on hydroxyindole-O-methyltransferase (HIOMT) protein levels and melatonin concentration in duodenum, as well as mRNA expression of melatonin receptor (MT)-1 in liver of fructose-fed mice. (a) Densitometric analysis of HIOMT staining in duodenal sections and (b) melatonin concentration in duodenum of mice. (c) Expression of MT-1 mRNA in the liver of mice. Values are means with their standard errors represented by vertical bars. FD, fructose-enriched liquid diet. *P<0·05 between FD- and FD+SoB-fed groups as determined using t test.
Expression of hydroxyindole-O-methyltransferase and melatonin receptor-2 mRNA and melatonin protein levels in differentiated human carcinoma colon-2 cells treated with sodium butyrate and expression of melatonin receptor-1/-2 and inducible nitric oxide synthase mRNA, as well as nitrate concentration in lipopolysaccharide-challenged J774A.1 concomitantly treated with melatonin
To further delineate the effect of SoB on melatonin synthesis in enterocytes, differentiated Caco-2 cells being a model of small intestinal enterocytes( Reference Russo, Luciani and De 24 ) were incubated with SoB for 6–48 h. Treatment with 3 or 6 mm-SoB did not change the HIOMT or MT-2 mRNA expression in Caco-2 cells after 6 h. However, expressions of both HIOMT and MT-2 mRNA were significantly induced in Caco-2 cells incubated with 3 mm-SoB when compared with naïve controls after 24 h, whereas in cells incubated with 6 mm-SoB a similar effect was not found (Fig. 6(a) and (b)). Expression of MT-1 mRNA in Caco-2 cells was below the level of detection at all time points and SoB concentrations used. In line with the findings for HIOMT and MT-2, melatonin concentration in the medium also increased in a time- and dose-dependent manner, with levels being significantly higher in media of cells incubated with 6 mm-SoB after 24 h when compared with naïve cells (Fig. 6(c)). After 48 h, melatonin concentration in media of cells treated with 3 or 6 mm-SoB was also higher than in naïve controls; however, differences did not reach the level of significance (P=0·31) (Fig. 6(c)).

Fig. 6 Effect of sodium butyrate (SoB) on hydroxyindole-O-methyltransferase (HIOMT) and melatonin receptor (MT)-2 mRNA expression, as well as melatonin concentration, in human carcinoma colon-2 (Caco-2) cells. Expression of (a) HIOMT and (b) MT-2 mRNA in differentiated Caco-2 cells 6 or 24 h after incubation with 0–6 mm-SoB. (c) Melatonin concentration in medium of differentiated Caco-2 cells 24 or 48 h after incubation with 0–6 mm-SoB. One-way ANOVA with Tukey’s post hoc test was used for statistical analysis of data. Values are means with their standard errors represented by vertical bars. C, control group. *P<0·05 compared with the group of Caco-2 cells without treatment for 24 h; ***P<0·05 compared with the group of Caco-2 cells treated with 6 mm-SoB for 24 h.
To determine whether melatonin has a protective effect on LPS-induced activation of Kupffer cells, J774A.1 cells being a model of Kupffer cells( Reference Kheder, Hobkirk and Stover 25 ) were challenged with LPS after a 2-h pre-incubation with melatonin. Expressions of both MT-1 and MT-2 mRNA were significantly induced in LPS-challenged J774A.1 cells preincubated with melatonin (Fig. 7(a) and (b)). While melatonin had no effect on iNOS mRNA expression or nitrate concentration in naïve J774A.1 cells, LPS-dependent induction of iNOS mRNA and increase of nitrate in the cell culture medium were significantly suppressed in cells concomitantly treated with melatonin (Fig. 7(c) and (d)).

Fig. 7 Effect of melatonin on mRNA expression of melatonin receptors (MT), lipopolysaccharide (LPS)-induced inducible nitric oxide synthase (iNOS) and nitrite concentration in J774A.1 macrophages. Expression of (a) MT-1, (b) MT-2 and (d) iNOS mRNA in the J774A.1 macrophages pretreated with melatonin and challenged with LPS and (c) nitrite concentration in media of these cells. One-way ANOVA with Tukey’s post hoc test was used for statistical analysis of data. Values are means with standard errors represented by vertical bars. C, control group; L, LPS-treated group. † P<0·05 compared with control group without treatment; ‡ P<0·05 compared with LPS-treated group; § P<0·05 compared with melatonin-pre-incubated control group; || P<0·05 compared with melatonin-pre-incubated and LPS-treated group.
Discussion
In the present study, despite receiving 0·6 g/kg body weight per d SoB in their diet, butyrate levels in portal plasma were similar among groups. It has been shown before by others that treating rodents and chicken, respectively, with a bolus dose of SoB resulted in a rapid increase in butyrate plasma levels; however, in these studies, doses of SoB were markedly higher than in the present study (5 v. 0·6 g/kg body weight) and SoB was administered as a bolus once by gavage or intraingluvial, whereas in the present study mice received SoB chronically with their liquid diet( Reference Egorin, Yuan and Sentz 26 , Reference Matis, Kulcsar and Turowski 27 ). Nevertheless, in mice fed FD supplemented with SoB, hepatic fat accumulation was markedly attenuated when compared with FD-fed mice. Furthermore, plasma ALT activity and markers of inflammation such as CCL-2 and TNF-α, but also PAI-1, of these mice were almost at the level of naïve controls. These data are in line with our own previous findings in mice fed a WSD supplemented with SoB but also with those of others( Reference Endo, Niioka and Kobayashi 9 , Reference Mattace, Simeoli and Russo 11 , Reference Jin, Sellmann and Engstler 14 , Reference Vinolo, Rodrigues and Festuccia 28 ). Furthermore, also in line with our own findings but also those of others( Reference Endo, Niioka and Kobayashi 9 , Reference Mattace, Simeoli and Russo 11 , Reference Jin, Sellmann and Engstler 14 , Reference Vinolo, Rodrigues and Festuccia 28 ), the protective effects of the SoB supplementation against the development of NAFLD were associated with a protection against the down-regulation of genes involved in insulin signalling in the liver – for example mRNA expression of IR and IRS-1. However, as the dietary feeding model used in the present study only induces early changes associated with the development of NAFLD and metabolic diseases, fasting glucose and insulin levels, as well as HOMA index, were not yet found to be altered between groups. Indeed, it has been shown before in patients with NAFLD that despite not yet displaying pathologically altered fasting glucose and insulin levels, markers of hepatic insulin signalling like IRS-1 mRNA were markedly lower than in controls( Reference Kanuri, Ladurner and Skibovskaya 29 ). Taken together, these data further bolster the hypothesis that an oral supplementation of SoB may protect mice from the onset of NAFLD and hepatic insulin resistance.
Results of several studies suggest that butyrate may be a critical factor in maintaining intestinal homoeostasis( Reference Leonel and Alvarez-Leite 30 ). Indeed, results of in vitro, as well as in vivo, studies suggest that butyrate may at least in part exert its beneficial effects in intestinal homoeostasis through increasing intestinal integrity and modulation of tight-junction proteins( Reference Endo, Niioka and Kobayashi 9 , Reference Wang, Wang and Wang 13 , Reference Ma, Fan and Li 31 ). Furthermore, both a loss of tight-junction proteins in the upper parts of the small intestine and increased plasma levels of endotoxin and expression of LBP mRNA have repeatedly been associated with the development of NAFLD( Reference Endo, Niioka and Kobayashi 9 ). However, in line with previous findings( Reference Jin, Sellmann and Engstler 14 ), in the present study mice fed the FD fortified with SoB were neither protected from the loss of the tight-junction proteins occludin and ZO-1 nor showed altered numbers of goblet cells in duodenum. In the present study, markers of intestinal barrier function such as tight-junction proteins were only determined in the upper parts of the small intestine. Therefore, it cannot be ruled out that SoB might also affect lower parts of the intestine, thereby adding to the effects observed in the present study. However, in line with the results of tight-junction protein levels, concentration of bacterial endotoxin in portal vein and expression of LBP mRNA in the liver shown to strongly correlate with bacterial endotoxin levels( Reference Douhara, Moriya and Yoshiji 32 , Reference Zuo, He and Liu 33 ) were also similar between the FD groups. Results of animal, as well as human, studies further suggest that an induction of the TLR-4-signalling cascade in the liver may be critical in the development of NAFLD( Reference Wagnerberger, Spruss and Kanuri 22 , Reference Kanuri, Ladurner and Skibovskaya 29 , Reference Csak, Pillai and Ganz 34 – Reference Spruss, Kanuri and Wagnerberger 36 ). Indeed, Rivera et al. ( Reference Rivera, Adegboyega and van 35 ) but also our own group( Reference Spruss, Kanuri and Wagnerberger 36 ) showed that TLR-4-mutant (C3H/HeJ) mice were markedly protected from the development of NAFLD. In the present study, the marked inductions of TLR-4 expression but also of its adaptor protein MyD88 found in FD-fed mice when compared with naïve controls were almost completely attenuated by the supplementation of SoB. Furthermore, expression of iNOS protein, also shown before to be highly regulated through endotoxin- and TLR-4-dependent mechanisms in the liver( Reference Spruss, Kanuri and Uebel 37 ), and concentration of 4-HNE, as well as SOD-1 activity, mediating the clearance of superoxide anion to hydrogen peroxide( Reference Zhu, Zhang and Roneker 38 ), were also only induced in livers of mice fed FD when compared with naïve controls. This effect of the diet was almost completely attenuated in livers of mice fed FD supplemented with SoB. The latter findings are contrary to those of others showing that the anti-inflammatory effects of SoB in a rat model of myocardial ischaemia/reperfusion were associated with a protection against the loss of SOD( Reference Hu, Zhang and Xu 39 ). However, in these studies, SoB was administered intraperitoneally to rats, whereas in the present study mice were fed SoB with their diet. Mechanisms involved in the effects of SoB on SOD-1 activity in livers of mice fed a FD remain to be determined. Mattace et al. ( Reference Mattace, Simeoli and Russo 11 ) also found that the protective effects of the supplementation of butyrate against the development of NAFLD induced by feeding a high-fat diet were associated with a protection against the induction of TLR-4 and iNOS expression in the liver. However, while in previous studies of our own group SoB-feeding was also associated with an attenuation of iNOS induction in the liver, TLR-4 expression in livers was unchanged( Reference Jin, Sellmann and Engstler 14 ). Differences between the two feeding experiments might have resulted from differences in composition of diets fed, for example a WSD rich in fructose but also fat and cholesterol v. a standard diet enriched with fructose in the present study. Taken together, results of the present study but also from previous studies of our own and other groups( Reference Mattace, Simeoli and Russo 11 , Reference Jin, Sellmann and Engstler 14 ) suggest that supplementation of butyrate may protect rodents from the development of liver damage through mechanisms involving a suppression of the induction of TLR-4 and subsequent signalling cascades in the liver. Indeed, butyrate has been suggested to directly alter enzymatic systems involved in oxidative defenses( Reference Russo, Luciani and De 24 ). However, as butyrate levels in portal vein were similar among all groups, the molecular mechanism involved in the antioxidative effects of SoB in the present study seems not to have resulted primarily from a direct effect of SoB on the liver but rather through indirect mechanisms (see below).
Results of older in vitro studies suggest that butyrate may modulate expression of enzymes involved in synthesis of melatonin( Reference Wiechmann and Burden 40 ). Indeed, it has been reported by Deng et al. ( Reference Deng, Lopez and Lynch 41 ) that in cultured Y79 human retinoblastoma cells treatment with 3 mm-SoB for 3 d markedly increased the release of melatonin. Furthermore, both HIOMT and arylalkylamine N-acetyltransferase have been shown to be expressed in enterochromaffin cells but also in enterocytes in the small intestine( Reference Konturek, Konturek and Brzozowski 42 , Reference Al-Ghoul, Abu-Shaqra and Park 43 ), suggesting that melatonin may also be synthesised from its precursor serotonin in the small intestine. In the present study, oral supplementation of SoB was associated with a significant induction of HIOMT and melatonin protein levels in the duodenum. In support of the findings that SoB may alter synthesis of melatonin in enterocytes, we showed that incubation of differentiated Caco-2 cells with SoB resulted in a marked time- and dose-dependent induction of HIOMT and MT-2 mRNA expression and melatonin release from these cells. Indeed, in the present study, melatonin receptors were shown to be induced in the presence of melatonin (see Fig. 7), suggesting that melatonin itself might regulate MT-1 and MT-2 expressions. The apparent discrepancy of the expression of HIOMT and melatonin levels found in Caco-2 cells at the different time points might have resulted from the different detection levels – for example mRNA expression and protein levels. In addition, it could be that as bioavailability of SoB decreased with time this might have affected the expression of HIOMT. This needs to be addressed in future studies. Melatonin has been shown to possess marked antioxidative capacities( Reference Zhang and Zhang 44 , Reference Manchester, Coto-Montes and Boga 45 ). Indeed, results of animal studies suggest that beneficial effects of melatonin in various settings of liver damage were associated with an attenuation of oxidative and inflammatory damage associated with these diseases( Reference Sharma and Rana 46 , Reference Hatzis, Ziakas and Kavantzas 47 ). It has been shown that treatment with melatonin but also its precursor tryptophan reduced levels of pro-inflammatory cytokines and improved parameters of fat metabolism in patients with NAFLD( Reference Celinski, Konturek and Slomka 48 ). Furthermore, Hatzis et al.( Reference Hatzis, Ziakas and Kavantzas 47 ) showed that the protective effects of melatonin treatment in vivo against high-fat diet-induced liver disease were associated with an attenuation of peroxidation and oxidative stress in liver tissue. In the present study, the increased levels of HIOMT and melatonin in the upper parts of the small intestine were associated with a significant induction of MT-1 in the liver but also a marked attenuation of lipid peroxidation and iNOS protein levels. In support of the hypothesis that in the present study the increased synthesis of melatonin in the small intestine of mice might have modulated endotoxin-dependent Kupffer cell-mediated response in the liver, both mRNA expression of iNOS and formation of nitrate were markedly attenuated in LPS-challenged J774A.1 cells when cells were concomitantly treated with melatonin. Furthermore, also somewhat supporting this hypothesis, expressions of MT-1 and MT-2 were also significantly induced in cells treated with melatonin. The slight induction of MT-1 and the even more pronounced induction of MT-2 found in J774A.1 cells treated only with LPS might have resulted from an LPS-dependent induction of melatonin synthesis in these cells. Indeed, results of a recent in vitro study suggest that enzymes involved in the synthesis of melatonin in macrophages are strongly regulated through LPS- and NF-κB-dependent signalling cascades( Reference Muxel, Laranjeira-Silva and Carvalho-Sousa 49 ). Taken together, these data suggest that supplementation of SoB may increase melatonin synthesis in enterocytes in the small intestine, which in turn, probably through MT-dependent signalling cascades, attenuated the induction of iNOS and lipid peroxidation in the liver. However, molecular mechanisms underlying the induction of intestinal melatonin synthesis, as well as pathways involved in the effects of melatonin on the liver, remain to be determined.
Conclusion
In summary, results of the present study are in line with previous findings showing that an oral supplementation of SoB has protective effects on the development of NAFLD( Reference Endo, Niioka and Kobayashi 9 , Reference Mattace, Simeoli and Russo 11 , Reference Jin, Sellmann and Engstler 14 ). Our results suggest that these effects do not result from a protection against the enhanced intestinal permeability and translocation of bacterial endotoxins associated with the development of NAFLD. Rather, our results suggest that a supplementation of SoB enhances intestinal melatonin synthesis and subsequently attenuates the endotoxin-dependent induction of iNOS and formation of reactive oxygen species in the liver. However, molecular mechanisms involved in the SoB-dependent modulation of intestinal melatonin synthesis and the effects of melatonin on iNOS in the liver need to be determined in future studies. Furthermore, future studies will have to delineate whether similar beneficial effects of an oral supplementation of SoB are also found in humans.
Acknowledgements
The project was funded in part by grants from the Federal Ministry of Education and Research and the German Research Society (BMBF, FKZ: 03105084 and 01KU1214A and DFG: BE2376/4-2 all to I. B.).
The contribution of each author was as follows: I. B. designed the research; C. J. J., A. J. E., C. S., D. Z., M. L., G. K., H. L. and M. S. conducted the research; C. J. J. and D. Z. analysed the data; C. J. J. and I. B. wrote the paper; I. B. had primary responsibility for the final content.
There are no conflicts of interest.
Supplementary materials
For supplementary material/s referred to in this article, please visit https://doi.org/10.1017/S0007114516004025











