


β-Carotene (βC) is one of the most common carotenoids in the human diet( Reference Britton, Liaaen-Jensen and Pfander 1 ), and its primary function in the body is provitamin A activity( Reference Grune, Lietz and Palou 2 ). All-trans (at)-βC is the most abundant isomer in fresh plant foods, but thermal processing can increase the proportion of 9-cis (9c)- and 13-cis (13c)-βC isomers up to 50 %( Reference Chandler and Schwartz 3 ). Little is known about the vitamin A (VA) efficacies of the cis-βC isomers, which have been collectively assigned a bioavailability equivalent to half that of at-βC( 4 ). Therefore, it is imperative to determine the unique bioefficacies of the cis-βC isomers to better understand the impact of thermal processing methods on the VA value of plant foods.
VA deficiency is a global health concern, affecting 190 million children under 5 years of age( 5 ). The introduction of provitamin A carotenoid-biofortified staple crops is a novel approach to alleviate VA deficiency and prevent issues observed with preformed VA supplementation and fortification, such as cyclical liver stores and hypervitaminosis risk( Reference Tanumihardjo 6 ). However, staple crops undergo thermal processing before consumption; thus, the impact of isomerisation may need to be considered when establishing biofortification targets in order to ensure adequate delivery of VA to target populations.
A study by Deming et al. ( Reference Deming, Baker and Erdman 7 ) has determined the individual bioavailabilities of 9c- and 13c-βC isomers in gerbils to be 38 and 62 % that of at-βC, respectively, using doses that ranged between 141 and 418 nmol βC/d or 132 and 392 nmol VA/d according to the Institute of Medicine (IOM)'s recommended values of 2 μg at-βC to 1 μg retinol activity equivalents for supplemental βC( 4 ). These orally administered doses greatly exceeded the utilisation rate (10·8 nmol VA/100 g body weight/d( Reference Lee, Lederman and Hofmann 8 )) and the current recommendation of the National Research Council for a typical adult gerbil (15·1 nmol VA/d, assuming an average feed intake of approximately 6 g( 9 )), and therefore they are not reflective of typical dietary intake that would be achieved from staple crops. Howe et al. ( Reference Howe, Maziya-Dixon and Tanumihardjo 10 ) found that βC-enhanced cassava feeds providing 10·9–27·5 nmol βC/d maintained VA status in gerbils despite 40–50 % cis-βC isomer content in cassava. Additionally, in an in vitro study using isomeric mixtures, the 9c-βC isomer enhanced overall total carotene in solution and resulted in constant or enhanced micellar incorporation( Reference Levin and Mokady 11 ). Thus, the objective of the present study was to compare the bioefficacies of the 9c-, 13c- and at-βC isomers in VA-depleted gerbils using doses reflective of dietary intakes from staple foods. Conversion to retinol was determined using total liver VA stores compared with a positive preformed retinol-supplemented group and a negative VA-depleted group.
Materials and methods
Chemicals and dose preparation
Crystalline at-βC (Sigma-Aldrich), 9c-βC and 13c-βC (gifts from Hoffmann-La Roche, Inc.) were prepared for purification by dissolution in n-hexane. βC isomers were purified on an open column using Al as the stationary phase and n-hexane as the mobile phase. The purities of all βC isomers were confirmed as >99 % with HPLC. For stock solutions, purified βC in hexane was condensed to 10 ml under N2, mixed with 20 ml cottonseed oil in an amber vial, and dried to remove remaining hexane and ensure clear solutions. The removal of hexane was confirmed by repeated weighing until a constant weight was achieved. Using spectrophotometry, concentrations were calculated with
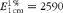 $$E _{1\hairsp cm}^{1\,\%} = 2590 $$
for the at-βC isomer at 450 nm,
$$E _{1\hairsp cm}^{1\,\%} = 2590 $$
for the at-βC isomer at 450 nm,
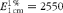 $$E _{1\hairsp cm}^{1\,\%} = 2550 $$
for the 9c-βC isomer at 445 nm and
$$E _{1\hairsp cm}^{1\,\%} = 2550 $$
for the 9c-βC isomer at 445 nm and
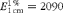 $$E _{1\hairsp cm}^{1\,\%} = 2090 $$
for the 13c-βC isomer at 443 nm(
Reference Schierle, Hardi, Faccin, Britton, Liaaen-Jensen and Pfander
12
). Specific volumes of stock solution were pipetted into amber vials containing the appropriate amount of oil to achieve the target dose and were mixed by sonication; final doses were validated by dissolution of the dose in hexane followed by spectrophotometry where the reference cell contained an equal amount of oil dissolved in hexane. Doses were divided and stored at − 70°C to minimise isomerisation. Lack of isomerisation during dose preparation was confirmed by running an aliquot of the oil dose dissolved in isopropanol on the HPLC system. A new bottle was opened at each study week.
$$E _{1\hairsp cm}^{1\,\%} = 2090 $$
for the 13c-βC isomer at 443 nm(
Reference Schierle, Hardi, Faccin, Britton, Liaaen-Jensen and Pfander
12
). Specific volumes of stock solution were pipetted into amber vials containing the appropriate amount of oil to achieve the target dose and were mixed by sonication; final doses were validated by dissolution of the dose in hexane followed by spectrophotometry where the reference cell contained an equal amount of oil dissolved in hexane. Doses were divided and stored at − 70°C to minimise isomerisation. Lack of isomerisation during dose preparation was confirmed by running an aliquot of the oil dose dissolved in isopropanol on the HPLC system. A new bottle was opened at each study week.
Study design
Male Mongolian gerbils (Meriones unguiculatus) (study 1, n 56; study 2, n 57; Charles River Laboratories), aged 34 d old, were group-housed (n 2–3 per cage) throughout the study. Room temperature and humidity were held constant (12 h light–dark cycle). Gerbils were acclimatised to the dosing procedure before the start of the treatments by orally administering 40 μl cottonseed oil/d using a positive displacement pipette. Gerbils were weighed on a daily basis for about 2 weeks and thereafter were weighed twice per week. Animal handling procedures were approved by the College of Agriculture and Life Sciences Animal Care and Use Committee at University of Wisconsin-Madison.
Gerbils consumed ad libitum water and VA- and carotenoid-free, semi-purified, powdered feed for the duration of the study. The feed was designed in consultation with a feed nutritionist (Harlan-Teklad) to meet the gerbils' energy and nutrient requirements without any known source of VA or βC (Table 1). Feed intake was determined by measuring the feed in a clean bowl and subtracting the remainder after a 24 h period. After depletion (study 1, 3 weeks; study 2, 4 weeks), a baseline group (study 1, n 6; study 2, n 7) was killed via exsanguination while under isoflurane anaesthesia to determine initial serum and liver VA concentrations. The remaining cages were sorted into weight-matched treatment groups (n 10 per group). To ensure 90 % power with a 5 % significance level, nine gerbils per treatment group were necessary to detect a 30 nmol difference in liver retinol concentration, assuming a standard deviation of 20 nmol. The sample size was increased to ten gerbils per group to account for the animals that may fail to thrive during the experimental period or to mitigate outliers.
Table 1 Composition of a vitamin A (VA)- and carotenoid-free gerbil feed

* Provided per kg of purified diet: biotin, 0·4 mg; calcium pantothenate, 66·1 mg; folic acid, 2 mg; inositol, 110·1 mg; menadione, 49·6 mg; niacin, 99·1 mg; p-aminobenzoic acid, 110·1 mg; pyridoxine-HCl, 22 mg; riboflavin, 22 mg; thiamin-HCl, 22 mg; vitamin B12 (0·1 % in mannitol), 29·7 mg; ascorbic acid (97·5 %), 1016·6 mg.
In the present study, three groups received daily oral doses of at-, 9c- or 13c-βC in cottonseed oil, and the other groups received doses of VA-free cottonseed oil (VA − ) or retinyl acetate in cottonseed oil (VA+). In study 1, a low βC dose of 15 nmol/d was designed to provide twice the molar equivalent of the daily VA utilisation rate of a 70 g gerbil( Reference Lee, Lederman and Hofmann 8 ). In study 2, a moderate βC dose of 30 nmol/d was designed to provide twice the molar equivalent of the current VA recommendation of 2·5 nmol/g feed( 9 ), assuming a typical feed intake of approximately 6 g/d. The retinyl acetate dose provided the proportion of 1 μg retinol:2 μg at-βC in accordance with the current equivalents of the IOM for oil βC supplements( 4 ). Following a 3-week (study 1) or a 4-week (study 2) treatment period, gerbils were killed. Blood was allowed to clot at room temperature for 30 min and centrifuged for 15 min at 2200 g at 4°C in vacutainer tubes (Becton Dickinson). Serum and liver samples were stored at − 70°C.
Tissue extraction procedures
Samples were analysed under gold light to prevent photo-oxidation and isomerisation. C-23 apo-carotenol was used as the internal standard in all serum and liver samples. A modified procedure was used for the analysis of serum( Reference Tanumihardjo and Howe 13 ). In study 1, 500 μl serum was analysed, reconstituted in 100 μl of methanol–dichloroethane (50:50), and 25 μl injected. In study 2, to assess the isomeric profile, 1000 μl serum was analysed, reconstituted in 80 μl of methanol–butanol (70:30), and 50 μl injected.
Liver (approximately 0·7 g) was ground with approximately 4 g sodium sulphate, extracted with dichloromethane, filtered and made up to 50 ml. For retinoid and carotenoid analyses, aliquots of 5 and 15 ml, respectively, were dried under N2, reconstituted in 100 μl of methanol–dichloroethane (50:50), and 25 or 50 μl injected.
HPLC analyses
βC isomers were separated from the oil doses and liver samples using a C30 YMC carotenoid column (4·6 × 250 mm, 3 μm; Waters, Inc.) at 450 nm, as described previously( Reference Howe and Tanumihardjo 14 ), on a Waters HPLC system consisting of a 1525 binary HPLC pump, a 717 autosampler and a 996 photodiode array detector. The samples were held at 4°C in the autosampler to prevent isomerisation or degradation.
For serum analysis in study 1 and all liver analyses, retinol and retinyl esters were separated using a Resolve C18 column (3·9 × 300 mm, 5 μm; Waters, Inc.) at 325 nm. Solvent A was acetonitrile–water (85:15) with 10 mm-ammonium acetate and solvent B was acetonitrile–methanol–dichloroethane (85:10:5) with 10 mm-ammonium acetate. Gradient elution was performed at 2 ml/min using the following procedure: 100 % A for 3 min, 7 min linear gradient to 100 % B, 15 min hold and 2 min reverse gradient to 100 % A. Identities of βC isomers, retinol and retinyl esters were determined using HPLC-purified standards. Total liver VA stores were calculated by summing retinol and identifiable retinyl esters. For serum analysis in study 2, retinol isomers were separated using a Zorbax ODS C18 column (4·6 × 250 mm, 5 μm; Dupont) at 325 nm with an isocratic mobile phase of methanol–water–butanol (50:27:23) with 10 mm-ammonium acetate run at 1 ml/min. Identities of the retinol isomers were confirmed using the elution order determined by MacCrehan & Schönberger( Reference MacCrehan and Schönberger 15 ) and HPLC spectra. All tissue samples were run on a Waters HPLC system consisting of a Delta 600 quaternary HPLC pump, a 600 controller and a 2487 dual absorbance detector, and the samples were injected manually. C-23 alcohol was used as the internal standard, and standard curves were constructed for the quantification of carotenoids and retinoids.
Statistical analysis
Results are presented as means and standard deviations. Data were analysed using SAS software (version 8.2; SAS Institute). Outcomes of interest (gerbil body and liver weights, serum retinol, and liver retinol and carotenoids) were evaluated using a one-way ANOVA. Differences between groups were determined using the least significant difference test at α < 0·05. An arcsine-square-root transformation was used to homogenise variance expressed as percentages. Bioconversion factors of βC to retinol were calculated using differences in total liver VA in the βC-treated groups compared directly with the VA+group after correction for the VA − group.
Results
Gerbil weights
In study 1, the final gerbil weights did not differ between the experimental groups (77·7 (sd 5·15) g) but were found to be higher than that of the baseline group (67·4 (sd 6·62) g, P= 0·0043). The final liver weights did not differ among the groups (3·05 (sd 0·42) g). In study 2, the final gerbil weights (68·8 (sd 5·63) g) and liver weights (2·47 (sd 0·35) g) were found to be similar among all the groups. Feed intakes did not differ within study 1 (6·42 (sd 1·07) g feed/d) or study 2 (5·05 (sd 0·69) g feed/d) (Table 2).
Table 2 Gerbil feed consumption in two studies administering low (15 nmol) and moderate (30 nmol) doses of β-carotene (βC) daily (Mean values and standard deviations)

c, cis; at, all-trans; VA, vitamin A.
Serum retinol and carotenoids
In both studies, total serum retinol concentration did not differ among the treatment groups, and carotenoids were not detected. In study 1, the range of serum retinol concentrations was 1·43 (sd 0·12) μmol/l in the 13c-βC group to 1·70 (sd 0·18)μmol/l in the VA − group. In study 2, the range was found to be 1·19 (sd 0·12) μmol/l in the at-βC group to 1·27 (sd 0·14) μmol/l in the baseline group, and the relative ratio of isomers varied across the treatment groups (Fig. 1). Serum retinol concentrations differed between studies 1 and 2 (P< 0·0001). The composition of 9c-retinol in serum differed among the groups (P= 0·032). The proportion of 9c-retinol was similar among the 9c-βC, 13c-βC, at-βC and VA+groups (1·94 (sd 0·50) %), and the proportion for the baseline and VA − groups (1·59 (sd 0·19) %) was found to be lower than that for the 13c-βC and at-βC groups (2·09 (sd 0·51) %), but did not differ from each other or the 9c-βC and VA+groups (1·77 (sd 0·46) %). The composition of 13c-retinol did not differ among the groups (3·16 (sd 1·95) %). The composition of at-retinol differed from baseline (96·7 (sd 0·50) %, P= 0·021) in the 13c-βC and at-βC groups, but did not differ from each other (94·1 (sd 1·94) %) or the VA − , 9c-βC and VA+groups (95·1 (sd 1·76) %).

Fig. 1 Serum retinol concentrations in Mongolian gerbils (Meriones unguiculatus) fed a vitamin A (VA)- and carotenoid-free feed at baseline after 4 weeks (study 2) of depletion (baseline, n 7), or treated with VA-free cottonseed oil (VA − ), moderate doses of 30 nmol 9-cis-, 13-cis- or all-trans β-carotene (9c- ■, 13c- ![]() or at-βC
or at-βC ![]() ) in cottonseed oil, or retinyl acetate in cottonseed oil (VA+) for an additional 4 weeks. VA intake in the VA+ group was matched to one-half of the theoretical VA intake in the βC groups. Values are means (n 10), with standard deviations represented by vertical bars. The treatment groups did not differ significantly (P>0·05; ANOVA followed by least significant difference test).
) in cottonseed oil, or retinyl acetate in cottonseed oil (VA+) for an additional 4 weeks. VA intake in the VA+ group was matched to one-half of the theoretical VA intake in the βC groups. Values are means (n 10), with standard deviations represented by vertical bars. The treatment groups did not differ significantly (P>0·05; ANOVA followed by least significant difference test).
Liver retinol and carotenoids
In study 1, concentrations of liver retinol did not differ among the experimental groups, and the βC isomers did not maintain baseline liver stores in all the groups (P= 0·006; Fig. 2(A)). Total liver retinol concentrations did not differ among the VA+, 9c-βC, 13c-βC and at-βC groups, and the βC isomers failed to maintain baseline liver VA stores in all of these groups, except the VA+group (P= 0·0026; Fig. 2(B)). The 9c- and 13c-βC groups did not differ from the VA − group. In study 2, the βC isomers maintained baseline liver retinol status in all groups, except the VA − group, which did not differ from the 9c- and 13c-βC groups (P= 0·0019; Fig. 3(A)). Total liver retinol stores were found to be highest in the VA+group and lowest in the VA − group (P< 0·0001; Fig. 3(B)). The VA − group did not differ from the 9c-βC group, but all the βC isomers resulted in similar total liver stores.

Fig. 2 Liver retinol (A) and total liver retinol (B) concentrations corrected for liver weight in Mongolian gerbils (Meriones unguiculatus) fed a vitamin A (VA)- and carotenoid-free feed at baseline after 3 weeks (study 1) of depletion (baseline, n 6), or treated with VA-free cottonseed oil (VA − ), low doses of 15 nmol 9-cis-, 13-cis- or all-trans β-carotene (9c-, 13c- or at-βC) in cottonseed oil, or retinyl acetate in cottonseed oil (VA+) for an additional 3 weeks. VA intake in the VA group was given at 1 μg retinol equivalent to 2 μg βC provided to the βC groups. Liver retinol was determined by pooling values for retinol and retinyl esters. Values are means (n 10), with standard deviations represented by vertical bars. a,b,cMean values with unlike letters were significantly different (P< 0·05; ANOVA followed by least significant difference test).

Fig. 3 Liver retinol (A) and total liver retinol (B) concentrations in Mongolian gerbils (Meriones unguiculatus) fed a vitamin A (VA)- and carotenoid-free feed at baseline after 4 weeks (study 2) of depletion (baseline, n 7), or treated with VA-free cottonseed oil (VA − ), moderate doses of 30 nmol 9-cis-, 13-cis- or all-trans β-carotene (9c-, 13c- or at-βC) in cottonseed oil, or retinyl acetate in cottonseed oil (VA+) for an additional 4 weeks. VA intake in the VA group was given at 1 μg retinol equivalent to 2 μg βC provided to the βC groups. Liver retinol was determined by pooling values for retinol and retinyl esters. Values are means (n 10), with standard deviations represented by vertical bars. a,b,cMean values with unlike letters were significantly different (P< 0·05; ANOVA followed by least significant difference test).
Carotenoids were not detected in the liver of the baseline, VA − and VA+groups in either study. In studies 1 and 2, total liver βC concentration was found to be highest in the 9c-βC group followed by the at-βC and 13c-βC groups (study 1, P< 0·0001; study 2, P< 0·0001; Table 3). The presence of the administered βC isomer and the other two isomers in the liver demonstrated in vivo isomerisation of the βC doses, and the relative proportions of the βC isomers varied among these groups (Table 3).
Table 3 Total liver β-carotene (βC) and isomeric compositions in Mongolian gerbils (Meriones unguiculatus) administered with low (15 nmol) and moderate (30 nmol) doses of 9-cis-, 13-cis- or all-trans βC (9c-, 13c- or at-βC)* (Mean values and standard deviations; n 10 per βC group)

a,b,cMean values within a column with unlike superscript letters were significantly different (P< 0·05).
* The limit of detection for βC was < 0·007 nmol.
Bioconversion factors and utilisation rates
Bioconversion factors for the βC-treated groups were calculated by determining the total liver VA stores above the negative control and by directly comparing this value to that of the VA+group. Bioconversion factors ranged from 115 to 168 % and 140 to 175 % of the IOM's recommended values in studies 1 and 2, respectively (Table 4). Comparison of the 9c- and 13c-βC isomers with the at-βC isomer resulted in mass equivalents of 2·9 and 1·6 μg 9c-βC and 2·6 and 1·4 μg 13c-βC to 1 μg at-βC for studies 1 and 2, respectively. In study 1, the utilisation rate for the VA − group, which was deprived of VA for 21 d beyond baseline, was found to be 5·1 μg/100 g body weight and for the VA+group, which was supplemented cumulatively with 84·6 μg retinol in the form of acetate, was 7·5 μg/100 g body weight. In study 2, the utilisation rate for the VA − group, which was deprived of VA for an additional 14 d in the extended depletion and treatment periods, was found to be 2·7 μg/100 g body weight, and for the VA+group, which received a total of 225·7 μg retinol, was 9·1 μg/100 g body weight.
Table 4 Bioconversion factors in Mongolian gerbils (Meriones unguiculatus) administered with low (15 nmol) or moderate (30 nmol) doses of β-carotene (βC)*

IOM, Institute of Medicine; at, all-trans; c, cis.
* n 10 per βC group.
† The IOM reference value for supplemental cis-βC isomers was based on the current retinol activity equivalents from food, which assumes a bioavailability that is equivalent to 50 % that of at-βC for all cis-βC isomers.
‡ Determined by direct comparison with bioconversion factors for the at-βC isomer in studies 1 and 2.
Discussion
To assess the impact of thermal processing methods on plant foods, it is essential to determine the unique VA bioefficacies of cis-βC isomers at physiological levels. To our knowledge, no other studies have investigated the bioefficacies of βC isomers using amounts reflective of dietary intakes in animals with carotenoid metabolism similar to that of humans. In studies 1 and 2, the bioefficacies of 9c-, 13c- and at-βC in oil at low (15 nmol) and moderate (30 nmol) doses were compared directly with preformed VA supplements. In study 1, the βC isomers did not maintain liver retinol stores; however, in study 2, at higher doses, all βC isomers maintained baseline liver stores and the VA supplement exceeded baseline VA status. Bioconversion factors were 15–75 % less efficient than the current reference values of the IOM. Factors for the supplemental cis-βC isomers were similar in each study, with midpoints of 6·4 and 5·3 μg cis-βC:1 μg retinol, in studies 1 and 2, respectively. Bioconversion factors for the supplemental at-βC isomer were found to be 2·3 and 3·5 μg at-βC:1 μg retinol in both studies, which are similar to the reported values of 2·5–4·6 in previous gerbil studies( Reference Howe, Maziya-Dixon and Tanumihardjo 10 , Reference Tanumihardjo and Howe 13 , Reference Howe and Tanumihardjo 14 , Reference Davis, Jing and Howe 16 ).
The utilisation rates within and between each study were not the same, but similar to those reported in previous gerbil studies( Reference Lee, Lederman and Hofmann 8 , Reference Davis, Jing and Howe 16 ). For the VA − group, the utilisation rate in study 2 was almost half of that in study 1, indicating that the gerbils were conserving and recycling more retinol during the period of prolonged deprivation. The higher utilisation of the VA+group in study 2 probably reflects the more concentrated preformed retinyl acetate dose that they received( Reference Green, Green and Lewis 17 ). The βC doses failed to maintain baseline liver VA stores in study 1, suggesting that the utilisation rates of gerbils exceeded the amount of bioavailable provitamin A provided. Lee et al. ( Reference Lee, Lederman and Hofmann 8 ) determined the VA utilisation rate of a VA-deplete adult gerbil to be 3·1 μg/100 g body weight. The difference in utilisation rates between study 1 (i.e. 5·1 μg/100 g body weight) and Lee et al. ( Reference Lee, Lederman and Hofmann 8 ) may be due to the gerbils having a higher requirement as they were 16–26 d younger, or due to the variation in the level of depletion( Reference Davis, Jing and Howe 16 , Reference Green, Green and Lewis 17 ). In study 2, the depletion period was extended to reach deficiency, which resulted in a similar utilisation rate (i.e. 2·7 μg/100 g body weight) to that of the Lee et al.'s value, and doses were increased to allow for adequate VA storage in the βC-treated groups.
It is important to consider the impact of VA status on bioconversion factors. The utilisation rate slows and recycling becomes more efficient during the state of deficiency and the converse occurs when the status is sufficient( Reference Green, Green and Lewis 17 , Reference Tanumihardjo 18 ). This departure in utilisation rates between the VA-deplete and VA-replete groups can therefore lead to the inflation of conversion factors, which may partially explain the higher conversion factors in study 1 compared with study 2 for the cis-βC isomers. The VA − group had higher VA status in study 1 than in study 2 where the depletion and treatment periods were extended. Furthermore, bioconversion of βC to retinol in humans and gerbils is dependent on dose size( Reference Tang, Qin and Dolnikowski 19 , Reference Simon, Tanumihardjo, Clevidence, Culver and Wrolstad 20 ) and is inversely related to VA status( Reference Ribaya-Mercado, Solon and Solon 21 , Reference Haskell, Jamil and Hassan 22 ), which may explain the difference in bioconversion factors for the at-βC isomer between the two studies because the dose was doubled in study 2.
In study 2, the relative VA values of the 9c- and 13c-βC isomers were similar with bioefficacies of 63 and 70 % that of the at-βC isomer, respectively. In previous studies, the relative VA values of the 9c- and 13c-βC isomers compared with that of the at-βC isomer were 33–61 and 48–74 %, respectively, in rats( Reference Duel, Johnston and Meserve 23 – Reference Sweeny and Marsh 25 ) and 38 and 62 %, respectively, in gerbils( Reference Deming, Baker and Erdman 7 ). The variations in the VA values determined in the previous studies may be due to the differences in the amount (0·6–224·4 μg βC/d) and duration (7–28 d) of dosing, the depletion period (20–56 d) or the basis of calculation (i.e. body-weight gain v. liver VA)( Reference Deming, Baker and Erdman 7 , Reference Duel, Johnston and Meserve 23 – Reference Sweeny and Marsh 25 ). The gerbil provides a more advantageous model for studying carotenoid metabolism compared with the rat because, like humans, they are capable of cleaving provitamin A carotenoids to retinal and absorbing physiological doses intact( Reference Lee, Lederman and Hofmann 8 , Reference Pollack, Campbell and Potter 26 , Reference House, Apgar and Smith 27 ). Although rats are efficient cleavers of βC in the small intestine, they only absorb βC intact under conditions of great dietary excess( Reference Ribaya-Mercado, Holmgren and Fox 28 – Reference Grolier, Agoudave and Azais-Braesco 30 ). The present study demonstrates more efficient conversion of the cis-βC isomers, particularly 9c-βC, in gerbils than that found by Deming et al. ( Reference Deming, Baker and Erdman 7 ), which, again, is likely attributable to the larger dose size used in that study.
The observation of higher total liver βC concentration found in the 9c-βC group suggests that the 9c-βC isomer may be more efficiently delivered to the liver. In vitro studies have suggested that the cis-βC isomers are similarly or more efficiently incorporated into micelles during digestion( Reference Levin and Mokady 11 , Reference Ferruzzi, Lumpkin and Schwartz 31 – Reference Aherne, Daly and Jiwan 33 ), but data are inconsistent regarding the efficiency of cis-βC isomer uptake by intestinal cells( Reference Ferruzzi, Lumpkin and Schwartz 31 – Reference Aherne, Daly and Jiwan 33 ). Based on accumulation in human serum, the at-βC isomer appears to be absorbed and/or packaged into the chylomicra more efficiently than the cis-βC isomers, with 13c-βC being the predominant cis-βC isomer( Reference Rushin, Catignani and Schwartz 34 – Reference Deming, Teixeria and Erdman 37 ). Additionally, in human subjects fed supplements or Dunaliella bardawil containing an at-βC:9c-βC ratio of approximately 50:50, the at-βC isomer appears to be preferentially incorporated into the chylomicra and VLDL( Reference Stahl, Schwarz and Sies 38 – Reference Ben-Amotz and Levy 40 ). However, in vivo interconversion of βC isomers, documented previously in human subjects( Reference You, Parker and Goodman 41 ) and gerbils( Reference Deming, Teixeria and Erdman 37 ) and confirmed in the present study, may have a positive impact on the VA values of the cis-βC isomers, and result in inaccurate interpretations of their bioavailabilities when confirmed solely by the presence of the isomer of interest. More work is necessary to elucidate common in vivo patterns for and mechanisms of βC isomer interconversion as none is apparent from the isomeric composition found in the gerbil liver in these studies. Furthermore, enzymatic isomerisation of cis-βC isomers to at-retinal by β-carotene 15,15′-mono-oxygenase 1 was observed in rats, with 9c-βC converting to a mixture of 9c-, 13c- and at-retinal and 13c- and at-βC converting primarily to at-retinal( Reference Nagao and Olson 42 ). In a previous study, the composition of retinol isomers in the liver of gerbils was determined to be similar among groups dosed with βC isomers and the negative control, with cis-βC isomers constituting approximately 10 % of total liver VA( Reference Deming, Teixeria and Erdman 37 ). Similarly, the isomeric composition of serum retinol in study 2 was found to be 3·3–6·5 % of cis-retinol isomers, which supports the in vivo isomerisation of βC and/or retinol. Together, these findings demonstrate that the isomeric compositions of liver and serum retinol do not reflect the dietary intake of βC, and that the at-retinol isomer appears to be preferable for the storage and circulation of VA.
The lack of the difference observed in liver VA stores among the 9c-, 13c- and at-βC groups and bioefficacies of the cis-βC isomers >60 % that of the at-βC isomer contradicts the idea that the cis:trans ratio significantly decreases the VA value of plant foods. These findings support the conclusion made by Howe et al. ( Reference Howe, Maziya-Dixon and Tanumihardjo 10 ) that regular, moderate intakes of cis-βC isomers from plant foods allow for efficient absorption and bioconversion to VA in gerbils. The cassava feeds used by Howe et al. contained 41–47 % of cis-βC isomers; by using a bioefficacy of cis-βC isomers equivalent to two-thirds that of the at-βC isomer, the VA value may have been decreased by only 14–16 %. In fact, Howe et al. ( Reference Howe, Maziya-Dixon and Tanumihardjo 10 ) demonstrated a conversion factor of 3·7 μg βC:1 μg retinol, which is similar to the values from plant foods containing much lower concentrations of cis-βC isomers( Reference Mills, Simon and Tanumihardjo 43 , Reference Arscott, Howe and Davis 44 ). Similar bioconversion factors for thermally processed provitamin A-biofortified cassava have recently been determined in women, ranging from 4·2–4·5 μg βC:1 μg retinol depending on oil content( Reference La Frano, Woodhouse and Burnett 45 ). Additionally, the matrix of certain plant foods can have a negative impact on the VA value( Reference Tanumihardjo, Palacios and Pixley 46 ), in which case thermal processing can increase βC bioavailability( Reference Aherne, Daly and Jiwan 33 , Reference Rock, Lovalvo and Emenhiser 47 , Reference Bresnahan, Arscott and Khanna 48 ). It is therefore important for nutritionists and plant breeders to holistically consider the impacts of processing on matrix effects and carotenoid concentrations when establishing provitamin A biofortification targets in staple crops.
Liver is the primary storage site of retinol in mammals and is considered the best measure of VA status( Reference Tanumihardjo 18 ). Serum retinol concentrations do not provide an effective measure of status because of homeostatic regulation over a wide range of liver stores( Reference Tanumihardjo 18 ), as demonstrated by similar concentrations among all the treatment groups within both studies. In that regard, the difference between serum retinol concentrations between studies 1 and 2 is interesting, considering that the gerbils in study 2 received twice the supplemental doses and liver VA concentrations did not differ between the two studies. Subclinical deficiency has recently been defined as liver stores < 0·1 μmol retinol/g liver( Reference Tanumihardjo 18 ); according to this definition, the VA − group in study 1 was nearing deficiency and the VA − group in study 2 was deficient, but serum retinol concentrations were not low (i.e. concentrations were >0·7 μmol/l). Additionally, the absence of carotenoids in serum is expected because the lipid profile of gerbils differs from that of humans in that the major circulating lipoprotein is HDL and carotenoids are predominantly associated with LDL during fasting( Reference Dosti, Mills and Simon 49 ).
In summary, the 9c-, 13c- and at-βC isomers prevented retinol deficiency in gerbils when provided in amounts reflective of moderate dietary intake, and the cis-βC isomers demonstrated bioefficacies >60 % that of the at-βC isomer when baseline liver stores were maintained. The results of these studies suggest that mild thermal processing modestly decreases the VA value of plant foods through the generation of the cis-βC isomers. Furthermore, the currently assigned bioavailability of the cis-βC isomers, equivalent to 50 % that of the at-βC isomer( 4 ), may need to be clarified as being dependent on dose size and VA status. More research is needed to elucidate the mechanism of in vivo isomerisation of cis-βC to at-βC and to assess the bioavailabilities of cis-βC isomers from plant foods in human subjects.
Acknowledgements
We thank Peter Crump, Senior Information Processing Consultant at the University of Wisconsin-Madison College of Agriculture and Life Sciences Statistical Consulting Service, for providing statistical assistance.
The present study was supported by USDA Hatch WIS01528, a College of Agricultural and Life Sciences Honors Scholarship (to K. A. B), a gift from Standard Process, Inc., and HarvestPlus. HarvestPlus (www.harvestplus.org) is a global alliance of agriculture and nutrition research institutions working to increase the micronutrient density of staple food crops through biofortification. The views expressed herein do not necessarily reflect those of HarvestPlus. The funders had no role in the design, analysis or writing of this article.
The authors' contributions are as follows: K. A. B. conducted the study; C. R. D. conducted the analyses; K. A. B. and S. A. T. analysed the data, designed the research and wrote the manuscript. All authors read and approved the final version of the paper.
Part of this work was presented at Experimental Biology Meetings in 2011 and 2013.
The authors have no conflicts of interest to declare.







