


16.1 Introduction
In this chapter, the history of historiography and the philosophy of history is brought to the aid of the history and philosophy of science (Reference UebelUebel 2017; Reference RothRoth 2020; Reference VirmajokiVirmajoki 2020; Reference KuukkanenKuukkanen 2012). Narrative has sometimes been taken to define historical knowledge, and to define it in contrast with scientific knowledge. The Narrative Science Project undermines this contrast (Reference WiseMorgan and Wise 2017; Reference MorganMorgan 2017; Reference WiseWise 2017; Reference Cristalli, Herring, Jones, Kiprijanov and SellarsCristalli 2019; Reference GriesemerGriesemer 1996). If narrative is a constitutive feature of scientific knowledge then perhaps the making of historical and scientific knowledge is more similar than has otherwise been assumed or allowed. For historians and philosophers who have investigated the so-called historical sciences, most prominently geology, palaeontology, evolutionary biology and natural history (Reference CurrieCurrie 2018; Reference ClelandCleland 2011; Reference RudwickRudwick 1985; Reference GallieGallie 1955; Reference Richards, Nitecki and NiteckiRichards 1992; Reference HubálekHubálek 2021), or who have attended to science’s archival practices (Reference DastonDaston 2017; Reference StrasserStrasser 2019; Reference LeonelliLeonelli 2016), such similarities might already seem obvious.Footnote 1 But the approach taken here extends beyond these bounds.
Not only is it the case that scientific knowledge contains more narrative than has been appreciated, but historical knowledge contains less. Historical knowledge has existed in many forms, not all of which are indebted to narrative. Chronicles and genealogies are among the most well-known alternatives which do not assimilate to narratives, although they may possess narrativity. If it is the case that within historiography we recognize narrative as only one part of our epistemic apparatus (working with chronicles and genealogies), and we also find narrative at work in science, then perhaps there is something about the relations between chronicle, genealogy and narrative within historiography that might be illuminating within the sciences. This chapter argues that this is indeed the case. It does so through an analogy between, on the one hand, the making of new historiographical fields and subfields, and, on the other, the making of new scientific fields and subfields. It argues that the process is relational, with field-forming choices taken by individual historians and scientists being made to a considerable extent through reflection on the apparent field-forming choices made by others. The content of these choices tracks the terrain of chronicle, genealogy and narrative. Sections 16.2 and 16.3 acclimatize the reader to thinking with these three forms of knowledge within historiography. Section 16.4 applies them to a case study in the sciences.Footnote 2
16.2 Three Forms of Historical Knowledge
Chronicles are some of the earliest known examples of historical writing and thought (Reference BreisachBreisach 2007; Reference AurellAurell 2004). While their variety of contents and styles is considerable, they can be grouped together thanks to their sharing some key exaggerated features. A convenient example is included in the University of Leeds digital collection of Medieval Manuscripts, which are freely available online.Footnote 3 The ‘Anonimalle Chronicle’, which can be found there, is a fourteenth-century manuscript which exemplifies key features of a chronicle (Reference Childs and TaylorChilds and Taylor 1991). A chronicle can be eclectic, but establishes rough terms for what it will include, bounded by some geographical or temporal limit. It records people and events deemed important. For example, in the case of the Anonimalle, this includes the Peasants’ Revolt. There is little thematic or argumentative ordering, as chronicles are mainly organized according to sequences and chronology (Reference SpiegelSpiegel 2016). While there may be some evidence of forward referencing from past events to future ones, so that there is room for some overarching narrativity, these features are muted (Reference PollardPollard 1938). At different times, and in different cultures, what it has meant to produce a factual account, and the means by which a chronicle’s evidences and descriptions have been assessed as reliable, have varied considerably. Today, key distinctions between historiographical approaches very often hinge on changes in the chronicle. For instance, while feminist historiography has inspired many significant and ongoing changes, the most fundamental has been recognition that the chronicles of history have been drawn ridiculously narrowly. The same can be said for those urging for global history-making, or environmental history-making or animal history. To boil things down, we can say that making a chronicle concerns choices of relevance and irrelevance, facing epistemic constraints of the present and the absent.
When it comes to genealogy, the most complete digitized work in the Leeds online collection is the ‘Biblical and genealogical chronicle from Adam and Eve to Louis XI of France’. This fifteenth-century manuscript includes a genealogical tree of the pedigrees of French kings and their descendants. It achieves this record both in tables and through tree diagrams showing these relations.Footnote 4 While there are many earlier examples of genealogical working and thinking, in Europe it was not until the twelfth and thirteenth centuries that this form came to be developed into a prose genre in its own right, its primary function being to define and legitimize particular lines of descent and their authority (Reference SpiegelSpiegel 1983; 1993). A historical genealogy finds ways to pick out certain objects that it can follow over time, objects bounded by some privileging rationale. The choice of ending point will have a direct and immediate effect on the overall message or moral, a choice which the genealogical author is considered responsible for. Genealogy was given a new lease of life in the second half of the twentieth century, adopted by a large and diverse set of historians and sociologists who found it could be fruitfully applied to histories of concepts and ideas. This mode is most commonly associated with Foucault, although his own broader debts in arriving at genealogy are worth remembering, as are alternative approaches to genealogical history (Reference RothRoth 1981; Reference BevirBevir 2008; Reference Prescott-CouchPrescott-Couch 2015). In addition, many publics commit themselves to making genealogies, be it of their DNA, or of their own family history, all of which has become big business (Reference NelsonNelson 2008; Reference Tutton and PrainsackTutton and Prainsack 2011). Often in genealogical research it is the finding of connections that matters over and above any explaining which those connections might achieve. In contemporary historiography, genealogies often distinguish themselves from narrative histories (discussed shortly), by explicitly resisting the latter’s epistemic expectations and genre conventions, particularly by denying closure. In its defining features, genealogy concerns choices of following and unfollowing, facing epistemic constraints of what it is possible or impossible to follow.
As with chronicles and genealogies, there is a wide range of different examples of narrative histories (Reference MomiglianoMomigliano 1990; Reference LevineLevine 1987; Reference BentleyBentley 1999). Nevertheless, while some form of narrativity is present in chronicles and genealogies, that narrative itself can be recognized as requiring its own care and attention within historical epistemology allows us to mark out a third distinct form of historical knowledge (Reference WhiteWhite 1987). I cannot account for the multiple potential origins of this form’s emergence, although it presumably occurred in piecemeal fashion somewhere between The Iliad and Braudel’s The Mediterranean. This form brings together a range of evidence to serve an argument or set of arguments, organized in the form of a narrative or set of narratives. A narrative has to know its end before it begins, and its terms are bounded by the questions it pursues. The motivations and justifications which take it from beginning to end (or from the end back to the beginning) are drawn from some present-centred interests which help to determine its informational order (we need to know X before we get to Y if we are to truly appreciate or agree to Z). Sometimes the written account will be narrated much like an unfolding novel, other times it is intended for a narrative to be read into it. Even when leaning into the grandiose and the rhetorical, their ambitions remain factual. At times the presence of narrative in historical knowledge, as offering something too much like fantasy or storytelling, has been contentious (Reference WhiteWhite 1984; Reference SpiegelSpiegel 2007), but today it remains the dominant and preferred form of historical knowledge, facing little meaningful scepticism. Those who recognize narrative as providing a means of explanation in its own right can hold any rhetorical or storytelling features at arm’s length.Footnote 5 To boil its key features down: narrative concerns choices of beginnings and endings, and makes connections – explicit or implicit – between elements of evidence which constitute an argument about, or give an explanation of, their subject.
16.3 The Analytical Apparatus: Six Elements of Historical Knowledge
The three forms are not incompatible with one another, and most examples of historical knowing and understanding will contain aspects of all three. A narrative history necessarily adopts some chronicles and not others, while treating some objects genealogically and not others. A chronicle will necessarily serve some genealogical interests better than others and be more amenable to some narrative syntheses and not others. A genealogy necessarily includes some chronicles and not others and occupies some narrative worlds more than others. Having described them, I argue that the making of fields and subfields of history, and indeed the making of any given historian’s identity as a historian, is achieved by the combining of different choices concerning the chronicles, genealogies and narratives that one adopts or rejects. This understanding is partially inspired by Gabrielle Spiegel, particularly her work on the ‘social logic of the text’ (Reference Spiegel1990), and earlier work that I completed with Paolo Palladino on biological time (Reference Berry and PalladinoBerry and Palladino 2019). Some of these choices are aesthetic, others political, others correspond to competing epistemic goals and values. Other aspects of these choices concern the kind of time in which one wishes to situate one’s research objects and audiences. Different fields, subfields and historians assess the value of historical knowledge encoded in these three forms according to their own criteria. This process is relational: seeing in what one person or group is doing an excellence or an excess, and in some other person or group something improper or deficient. It is relational because these assessments help to motivate and justify change (or stasis) in oneself. When two or more historians arrive at the same or similar evaluative criteria, or when they share an emphasis on the importance of one or the other forms for a particular topic, we may discern the beginnings of a subfield.
It is this state of affairs within historiography, which, so this chapter argues, is paralleled in the sciences.Footnote 6 However, the descriptions of the three forms provided thus far has been too general for the purposes of making analogies between history and science. We need smaller focal points.
The six elements running down the rows of Table 16.1 have been intuited from reading history of historiography, narrative theory and philosophy of history. They concern ways in which the chronicle, the genealogy and the narrative are distinguishable. Some of these six elements are taken quite directly from the existing work of other scholars, and these debts will be clear in citations. However, the gloss which each is given serves the unique aims of this chapter. Section 16.4 applies these elements analogically to a case in the sciences.
1. Means of construction. When a chronicle is being compiled, one is primarily faced with choices of inclusion or exclusion, determined by whatever criterion one has adopted. When a genealogy is being composed, even if the key figures or subjects are picked out, the primary choices one faces concern which to follow when, and which to cease following when. As for the construction of a narrative, the most essential manoeuvre for the building of a narrative world concerns when to start (and why), and when to stop (and why), two decisions which are really one.
2. Means of ordering. The question of ordering has helped motivate the Narrative Science Project from the outset (Reference MorganMorgan 2017). The organizer of a chronicle works under the expectation that each entry will be ordered chronologically. Chronology may also matter for the organizer of a genealogy, but this will be mixed with a selectivity towards a particular object, the phenomenon of interest, that which is being traced over time. The term ‘material overlap’ – which I use to describe this means of ordering – I take from Reference GriesemerGriesemer (2000), which he introduced to help characterize what is interesting about evolutionary dynamics in particular, but which I think is extendable to anything lineal. When it comes to narrative, despite very clearly important concerns which often guard historians against presentism, the producer of a narrative will be inescapably presentist, and indeed that presentism contributes to a narrative’s value. Their materials will therefore be ordered according to their argumentative ambitions in the present.
3. Likely modes of narrativity. The six modes of narrativity that run across this row are all directly taken from Reference RyanRyan 1992, who also lists many more.Footnote 7 I have chosen the six which best help differentiate the three forms. To explain them very briefly, an embryonic narrative has some of the most important features for narrative-making without any identifiable plot (the historical chronicle is one of Ryan’s illustrative examples). A deferred narrative provides no narrative of its own but intervenes into something which might eventually become one (the example given is that of a newspaper report). Multiple narrativity keeps many plots in play at once but without requiring that they be related or interact (the suggested examples are The Decameron and The Arabian Nights). Braided narrativity might have plots which interact, join, depart from one another, etc. (Ryan’s preferred examples are family sagas and soap operas). Underlying narrativity is read into some source material without being stated in explicit narrative form (examples are offered from everyday life, such as witnessing a fight and interpreting it as the outcome of some longer set of events).Footnote 8 Historians commonly use the latter mode of narrativity in an effort to create distance between the materials presented and their own preferred narrative, or when they are attempting to delay the selection of one narrative over alternatives. Last, figural narrativity, which again arises outside of any explicitly stated narrative, and occurs when some source or other conjures up in our minds a stand-in, a figure, of one kind or another. One of Ryan’s preferred examples is the making of nation states into characters on a global historical stage.
4. Reflexivity. This row places the three on a scale, from low reflexivity to high. A chronicle requires little reflexivity on the part of the chronicler once the criteria for selection are established. As such, it might be better to say that the reflexivity of the chronicler is required prior to the making of the chronicle, rather than it being an explicit feature of the account. A genealogy requires a little more reflexivity because the choices concerning what to follow or unfollow, and when, cannot be specified prior to the composition, but will be more dependent on author choices. Last, a narrative history expects a very reflexive and self-conscious author, even when an externalizing ‘all-seeing’ voice is adopted.
5. Ending. A chronicle is not building to some ending or other but ends at some arbitrary point. It could easily continue, without any effect on its overall structure or significance, but it simply does not. A genealogy can only last as long as do the materials it is addressing. It could continue, provided the object continues, and different ending points can produce different lessons. The ending of a narrative history, meanwhile, will have been baked into it from the beginning, because it needs to be coherent with the questions it pursued (Reference RothRoth 2017; Reference MorganMorgan 2017).
6. Orientation to the world. This last distinguishing element is the most difficult to explain concisely. Very briefly, when we pick up either a chronicle, a genealogy, or a narrative history, we are also picking up the relationship between reader and world which each generates or assumes. This element is similar to ‘rhetorical structure’ as conceived in genre theory (Reference FrowFrow 2015), establishing the posture of the audience.Footnote 9 Chronicles, by and large, are written such that they are set against the backdrop of ‘all time’, or ‘God’s time’. ‘These are the chronicles of X’, some disembodied voice impresses upon us, ‘and they were recorded because they are important’. Genealogies instead orientate audiences by facts of existence, i.e., that some things which once were are no more, other things which have been are still, and still others that have not yet been, might. Finally, narrative histories orientate a reader between three points: (A) the world of the narrative; (B) the world as it has been known to the reader, and (C) an argument which is taken to describe and explain B through A.Footnote 10 Narrative history is a large and complex modelling exercise.
Having explained these elements, Table 16.1 can now be used both as a diagnostic for detecting the presence of the three forms of historical knowledge and as a means by which to more clearly distinguish between them. These elements will now be applied analogically to a case in the sciences – more specifically, the development of the field of synthetic biology.
Table 16.1 Reading history of historiography, narrative theory and philosophy of history
16.4 Narrative in the Sciences: The Case of Synthetic Biology
Synthetic biology is, among other things, an epistemic programme of reform. This programme is sometimes ‘imposed’ on the biological sciences by engineering-trained outsiders, but just as often is pursued by biologists and biochemists from within (Reference O’MalleyO’Malley 2009). The creation of this field, or subfield, has been relational in the sense that synthetic biologists explain their own aims and ambitions largely through comparison and contrast with alternative existing fields (the primary alternatives being molecular biology and engineering). On my terms, this is the normal state of affairs for all areas of science and historiography, which make and remake themselves by these relational claims and choices on a daily basis. But in the case of synthetic biology the process has been particularly pronounced and instructively explicit. Indeed, there are a vast array of things which synthetic biology seeks to, and often has to, distinguish itself from, in order to marshal any autonomy. The list of competitors ready to swallow it up include biochemistry, systems biology, genetics, microbiology, data-centric biology, molecular biology, developmental biology, biochemical engineering and biotechnology. Indeed, it might not make sense to conceive of synthetic biology as existing outside of the parallel development of some of these alternatives, particularly systems biology (which it has grown up alongside), each dealing with ‘epistemic competition’ in similar ways, although emphasizing different aspects of biological knowledge (Reference Gross, Kranke and MeunierGross, Kranke and Meunier 2019). Nor has synthetic biology entirely settled in one identity or another, with both biological engineering and design biology being alternatives sometimes adopted by practitioners and institutions.
This chapter explains the emergence of synthetic biology not only as a direct result of the influence of key charismatic individuals (Reference Campos, Harman and DietrichCampos 2013), nor only thanks to the availability of novel experimental commodities (Reference BerryBerry 2019), nor only by attachment to the aspirations of national and international patrons (Reference Schyfter and CalvertSchyfter and Calvert 2015), nor only as a product of techno-futurist venture capital (Reference Raimbault, Cointet and JolyRaimbault, Cointet and Joly 2016), alongside all the other candidate features of importance which scholars have already addressed, but also as the bringing together of a set of epistemic choices made relative to other subfields. The epistemic choices in question track the terrain of chronicle, genealogy and narrative.
Figure 16.1 contains most of the essential features one might need in order to illustrate the epistemology of synthetic biology. From the point of view of this chapter, the knowledge production and interpretation practices found in this image exemplify all six of the scientific analogues for the elements of historical knowledge explained above. It is an exemplary image of synthetic biology, and was intended to be, published as it was in a PhD thesis completed in a laboratory dedicated to bringing plants to synthetic biology and synthetic biology to plants (Reference Pollak WilliamsonPollak Williamson 2017).Footnote 11 It was produced to test predictions concerning the relative strengths of different promoters (lengths of DNA that raise the rate at which some other DNA in the cell gets transcribed, to ensure it gets expressed) in different regions of plant tissue. Such promoters are prototypical ‘parts’ for synthetic biologists. What parts are is sometimes a fraught question, but are in general lengths of DNA with some characterized and specified function. Here ‘characterized’ means that data has been generated describing their behaviour in one or several biological, chemical or biochemical contexts. The promoter parts in question were being tested for inclusion in a new registry of standardized parts, which would enable more plant scientists to work with the organism in question, Marchantia polymorpha, as a model organism (Reference Pollak WilliamsonDelmans, Pollak Williamson and Haseloff 2017). The combining of different radiating or fluorescing reporters with different microscope technologies provides some of the most important historical background to this particular case study (Reference WorliczekWorliczek 2020). In Figure 16.1, we see fluorescent proteins (FPs), which are the source of the dots that you can see in rows 1, 2 and 4, combined with confocal microscopy (the particular microscope technology which took these images).Footnote 12 Fluorescence in row 3 is produced by a dye. Figure 16.1 is also demonstrating a particular method, known as ‘ratiometry’. All of these features (standardized parts, registries, fluorescent proteins, confocal microscopy and ratiometry) will be explained in more detail below.
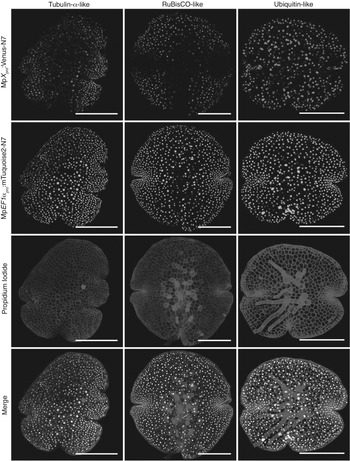
Figure 16.1 Patterns of expression of different promoters transferred to three plants
Columns 1–3 show that their expression is localized to different areas of plant tissue, visualized by monitoring two distinct fluorescent proteins (rows 1–2) attached to those promoters. Images taken through confocal microscopy, including one taken against a dye-stained background, which illuminates distinct plant cells (row 3), all overlaid in a composite image (row 4).
For some scholars, my decision to treat synthetic biology as a relatively well-defined field or subfield with a distinctive collective epistemic culture would be problematic, on the grounds that it is not quite so distinctive as it thinks itself to be and is substantially coextensive with existing fields of biological research. In this respect, my attention to confocal microscopy in combination with FPs will not be an effective way to explain or argue for the particular emergence of synthetic biology, because the tools and methods that I am focusing on matter far more widely than in synthetic biology alone, right through molecular and developmental biology (Reference BaxterBaxter 2019). For instance, Hannah Landecker has already recognized the importance of live cell imaging techniques (as seen in Figure 16.1) throughout the biological sciences, and attended to the novel epistemic perspective which they enable (Reference LandeckerLandecker 2012). Likewise, regarding the emphasis I will place on improving data quality and management: these characteristics have been studied extensively by Reference LeonelliSabina Leonelli (2016) in the wider phenomena of ‘data-centric’ biology. But, for myself, the point is that whatever significance these methods and representations might have throughout the biological sciences they are nevertheless regularly claimed on behalf of synthetic biology – and not entirely illegitimately, thanks to reasons that correspond to their epistemic choices and preferences. Of course, the extent to which actors’ epistemic choices and preferences are practised coherently, and the extent to which actors live up to their own self-image, are always important questions. But they simply fall outside the analytical bounds of this particular chapter.
16.4.1 Means of Construction
For synthetic biology, the most important choices in this respect concern inclusion and exclusion. Synthetic biology pushes for the rigour and standardization of engineering, rejecting the artisanal choices of molecular biology. As such, it emphasizes the chronicle form of knowledge.
Synthetic biologists often start with worries and complaints that too many molecular biologists have been recording too many insignificant details, with insufficient rigour, too idiosyncratically, too selfishly, for too long. On their estimation, the terms for deciding what should be included and excluded in shared records have not been attended to with sufficient care and scrutiny, and so they want to increase the relevance and value of what is recorded. These claims and goals are analogous to choices in historiography regarding what to incorporate into the chronicle. The promoter parts used in Figure 16.1, and the parts registry to which they were submitted, are a useful icon to think with in this context, particularly as discussion surrounding them has developed considerably since the early 2000s when they were first posited as necessary (Reference Frow and CalvertFrow and Calvert 2013; Reference Stavrianakis and BennettStavrianakis and Bennet 2014). The idea of a ‘parts registry’ (with an attendant physical repository for samples) is partially built on the back of earlier national and international infrastructures for the sharing of materials and services in biology, such as GenBank. But parts registries are also novel to synthetic biology in ways which are directly related to its dissatisfaction with molecular biology. The quality of a part’s characterization data is intended to be higher, and collected more rigorously and in more standardized ways than has been common in molecular biology, excluding anything esoteric or artisanal. On my terms, the application of the skills, technologies and methods which constitute parts, is intended to expand, and systematically to improve on, biology’s chronicle, in contradistinction with the repository and data collection practices performed by others. One should imagine historians choosing to replace or alter existing chronicles on the grounds that they were made inexpertly, or under the guidance of misleading prejudices.
16.4.2 Means of Ordering
Synthetic biology uses new techniques and technologies to see through multiple scales of life during processes of material overlap, avoiding the retrospective stitching together of dead bits and pieces which it considers characteristic of molecular biology. As such, it emphasizes the genealogical form of knowledge.
Images like those in Figure 16.1 are taken to evidence the synthetic biologist’s particular powers of observation, precision, and control over their materials as those organisms and materials undergo processes of material overlap. This is all the more so when used to demonstrate the desired activity of a molecular part, which in turn evidences their competence in designing and making. These technologies for visualization provide biological scientists finer-grained detail in the investigation of phenomena as they happen over time, at both molecular and phenotypic levels simultaneously, all while the cell, tissue or organism in question is still alive. On my terms, this concerns synthetic biology’s and molecular biology’s means of ordering. Both groups prize observing objects undergoing processes of material overlap at molecular and phenotypic scales simultaneously, but (so it is argued) only synthetic biologists have prioritized developing truly reliable means for doing so. By contrast, so it is said, molecular biologists addressing questions of biological development in relation to molecular and phenotypic scales have worked more indirectly, often retrospectively piecing together developmental sequences from dead matter, or addressing molecular and phenotypic levels separately rather than simultaneously (Reference AnkenyAnkeny 2001; Reference SchürchSchürch 2017). On my terms, synthetic biology claims to arrive at the same kind of genealogical knowledge sought by molecular biology, but better and more reliably. One should imagine historians finding apparent evidentiary smoking guns at the centre of developing historical phenomena, or finding new potential paths of connection.
16.4.3 Likely (or Available) Modes of Narrativity
Where molecular biology measures its success by greater or lesser incorporation into the narrative of evolution, synthetic biology measures success in making simpler and more finely tuned systems, more like engineering than those found in nature. As such, it once again emphasizes the chronicle form of knowledge.
The next feature of Figure 16.1 that we will discuss, its use of ratiometry, exemplifies the mode of narrativity which synthetic biology prefers. Massimiliano Simons has identified the emergence of ‘postcomplex’ life sciences in the twenty first century. This refers to ‘sciences [that] do not imply a denial of the complexity of nature at the experimental level, but rather […] desire to transcend it’ (Reference SimonsSimons 2019: 151). This is a very helpful way to understand what synthetic biologists are getting up to in general, and with ratiometry in particular.
In biology, ratiometry is a practice that was first developed in the biomedical sciences as an improvement on earlier fluorescence-based diagnostic and observational techniques (Reference Haidekker and TheodorakisHaidekker and Theodorakis 2016). Because living cells and tissues are so context sensitive, and subject to multiple complex influences, actors in biomedicine began to develop dyes that fluoresce at two different wavelengths of light. Measuring both frequencies provided a check on the overall biochemical context, while also gathering the actual reaction data which one is interested in. This is because the second fluorescence measurement can be used as a constant reference point. In Figure 16.1, the reference signal is found in Row 2. If the reference point behaves bizarrely, one has a reason to question the validity of the experiment and the data it yields. If, however, the reference point behaves well, one can gather even more precise data concerning the reaction of interest by monitoring the ratio between the two outputs, hence ‘ratiometrics’. While not all synthetic biologists use ratiometry, it is nevertheless precisely the kind of effort which the field celebrates and can be appreciated as forming part of their productive response to the ‘problem’ of biological ‘noise’ (Reference Knuuttila and LoettgersKnuuttila and Loettgers 2014). Interestingly, the practice and the term ‘ratiometric measurement’ originated outside of biology and biomedicine, within electrical engineering, where electrical rather than optical signals were used (Reference Holloway and NwaohaHolloway and Nwaoha 2013). The precision of the ratiometric results arrived at increases the chance that whatever design constraints are impinging on this biological context will become easier to spot, if not now then at some point in the future. This is an embryonic or deferred narrativity, more akin to what one finds in a chronicle, which synthetic biology prefers over and above the more figural or underlying evolutionary and biochemical narratives, which drive, and are prized in, molecular biology. In making this kind of choice, existing outside of evolutionary narrative-making, synthetic biology is by no means alone (Reference Love, Bueno, Chen and FaganLove 2018). One should imagine historians increasing sceptical pressure on certain evidences and standards of assessment, which often requires or affords a postponement on overarching conclusions.
16.4.4 Reflexivity
Synthetic biology interprets molecular biology as possessing low reflexivity because the latter has rarely considered the authors of protocols, metadata and other foundational sources as worthy of recognition. By contrast, synthetic biology increases authorial pride over protocols, metadata and so on. As such, it emphasizes the narrative form of knowledge.
At the same time as improving the materials used, synthetic biology also promises to improve the ways they are used, by prioritizing thoughtful, planned and well-managed sharing. The ambition is not only to increase the number of parts available for the synthetic biologist to work with, but to improve their capacity to work with them by also collecting as much useful data and metadata concerning their use as is feasible (Reference McLaughlin, Myers, Zundel and MısırlıMcLaughlin et al. 2018). Fluorescent proteins, and the parts they are used to characterize, are themselves expected to be created more reflexively in synthetic biology than they have been in molecular biology through the adoption of some explicit standard, which, it is hoped, will ensure their compatibility with other parts that have been made according to the same standard (Reference Peccoud, Blauvelt, Cai and CooperPeccoud et al. 2008). Sabina Leonelli’s research on the broader phenomena of data-centric biology is important here, and data ‘curators’, the class of experts permeating the biological sciences on whom Leonelli focuses, are particularly pronounced in synthetic biology (Reference LeonelliLeonelli 2016). These claims and goals emphasize authorial reflexivity. On my terms, by emphasizing greater reflexivity concerning authorship, synthetic biology is emphasizing the value of narrative knowledge for the field, in contrast with those fields outside of it. One should imagine curators and archivists developing international standards for description, the making of definitive translations of historical texts or the making of more widely accessible historical archives.
16.4.5 Ending
Where molecular biology seeks to find what has been and why, synthetic biology arrives at what can be, and (hopefully) an eventual understanding as to why, ending as engineering often does with something that works. As such, it emphasizes the genealogical form of knowledge.
Synthetic biology’s preferences for narrativity (section 16.4.3) are coextensive with its preferred ending points, which are often the demonstrations of what they can now make with their materials rather than necessarily a reflection on what questions they might answer (Reference SchyfterSchyfter 2013). Whatever complexity is present in the cells of Figure 16.1, and whatever overall biochemical or evolutionary narrative which these findings might contribute to, this research programme cuts through all of that, in order to produce a simpler, immediate and fine-grained picture of a system of protein expression, one which might be further refined or made tuneable, as in engineering. Of course, this does not stop researchers also considering evolutionary significances, but these are simply not a requirement for the field. On my terms, such endings most closely resemble genealogical knowledge, because parts and constructs could always be developed further, characterized further, put to work in more places, etc. But after they have been shown effective in at least one or two places the synthetic biologist can choose to stop. Nor are they required to place them in evolutionary context. These then are their preferred kinds of ending, which are demonstrative ones. One should imagine historians engaging in re-enactments, or developing new uses for old sources, or new methods by which to study sources, or simply finding new sources. These are all sufficient, as useful and valuable endings, in their own right.
16.4.6 Orientation to the World
Where molecular biology illuminates evolutionary lineages, and the persistence or loss of forms over time, synthetic biology renders organisms in the image of its argument for engineering. As such, it once again emphasizes the narrative form of knowledge.
When it comes to how synthetic biology orientates an audience to the world, design principles are key. This is a topic which Sara Green has addressed through the concept of ‘constraint-based generality’ in the closely aligned field of systems biology. ‘Constraint-based generality makes possible the identification of general principles underpinning a class of systems exhibiting similar structural or dynamic patterns’ (Reference GreenGreen 2015: 635). Where systems biologists attempt to find and understand these patterns in extant biological organisms and systems, synthetic biologists look for structural patterns not only in existing systems but also in the parts which they make (Reference KoskinenKoskinen 2017). When submitting parts to the registry and attempting to standardize the ways in which these parts are characterized, synthetic biologists are building up biology’s recognized design space, one part at a time. This is the world which synthetic biology orientates a reader to – some possible designed one rather than a merely evolutionary and natural historical one (Reference KellerKeller 2009; Reference Knuuttila and KoskinenKnuutilla and Koskinen 2021). Molecular biology therefore asserts its authority through contrast with an ‘actual’ genealogy of evolution and natural history. Interestingly, one of the ways in which Michel Morange allows that synthetic biology might prove liberating for evolutionary biology is by undermining its otherwise uniformitarian tendencies when it comes to the actual narratives of evolution (Reference MorangeMorange 2009: 374).Footnote 13 As synthetic biology’s constructs serve as justification for the approach and embody their argument, I interpret synthetic biology as producing an orientation to the world which is much more like narrative history rather than chronology or genealogy. Just as historians orientate the audience towards the world through their historical model, so do synthetic biologists orientate the audience to the world through their designed synthetic one.
16.5 Conclusion
In this chapter, I have offered six elements distinguishing three forms of knowledge, arguing that choices concerning these six elements are means by which fields and subfields define and differentiate themselves in relation to other fields and subfields, regarding their understanding of the world and their practices. While the three forms in question were derived from analyses of historiography, the chapter argues that they also undergird knowledge-making in the sciences. Table 16.1 lists these elements and the forms of knowledge they most typically align with, turning the table into a diagnostic tool. I have used the table to analyse features of synthetic biology present within Figure 16.1, illustrating how these six elements are located in practice in a scientific case study of a field’s formation and self-understanding of its knowledge-making. We come to appreciate that synthetic biology differentiates itself by sometimes emphasizing ‘chronicular’ knowledge, other times genealogical knowledge, other times narrative knowledge. When it comes to its means of construction and narrativity, it emphasizes the chronicle. When it comes to its research endings and preferred ordering of phenomena, it emphasizes genealogy. When it comes to its level of reflexivity and orientation to the world, it emphasizes the narrative form. If the overall picture has been effectively communicated, the reader will be able to recognize ways in which the process of making new scientific fields and their knowledge claims turns on similar considerations to those which arise in the process of making new historical fields and their knowledge claims. The aim has been to advance understanding in at least two directions.
First, the historians and scientists who populate any given period are rarely considered together and are typically treated as requiring different and distinct analytical apparatus. But this need not be the case. The kinds of analysis which some historians of science already make concerning the narrative, genre and literary conventions which have mattered for science throughout time (Reference Pomata, Ginzburg and BiasioriPomata 2018; Reference BucklandBuckland 2013; Reference BeerBeer 1983; Reference DearDear 1991) could be good starting points for such an approach. Aspects of these accounts will be illuminating for the history of science and historiography alike. As such, a future direction which historians and philosophers could take would involve looking across the waxing and waning of modes of narrativity, or preferred endings, or means of construction, etc., of a given period in science in tandem with historiographical fashions concerning the same, in the search for shared patterns of epistemic change. Such shared patterns might direct us towards underlying cultural shifts.
Second, this chapter has established a new framing for examining the formation of fields and subfields in both history and science. This approach prioritizes actors’ categories without remaining beholden to them. It is dynamic, as the six elements of knowledge formation described here can be applied imaginatively to a wide variety of aspects of scientific and historical life – be they publishing norms, experimental practices, representational preferences, intellectual property norms or what they study and how they study it. The differences between chronicles, genealogies and narratives, and the different times when we wish to emphasize the significance of the one or the other, are worth bringing to bear on more cases, particularly where clashing ‘narratives’ are believed to be in play, as between synthetic biology and its immediately adjacent subfields.Footnote 14



 Open access
Open access