
388 results

The Future of Qualitative Research in Healthcare
- The Role and Management of Digital Methods
- Coming soon
-
- Expected online publication date:
- August 2024
- Print publication:
- 31 August 2024
-
- Book
- Export citation
The Rapid ASKAP Continuum Survey III: Spectra and Polarisation In Cutouts of Extragalactic Sources (SPICE-RACS) first data release – CORRIGENDUM
-
- Journal:
- Publications of the Astronomical Society of Australia / Volume 41 / 2024
- Published online by Cambridge University Press:
- 03 June 2024, e039
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Becoming and acting as an ally against weight-based discrimination
-
- Journal:
- Industrial and Organizational Psychology / Volume 17 / Issue 1 / March 2024
- Published online by Cambridge University Press:
- 07 March 2024, pp. 142-147
-
- Article
- Export citation
Impact of primary care triage using the Head and Neck Cancer Risk Calculator version 2 on tertiary head and neck services in the post-coronavirus disease 2019 period
-
- Journal:
- The Journal of Laryngology & Otology , First View
- Published online by Cambridge University Press:
- 22 January 2024, pp. 1-6
-
- Article
-
- You have access
- HTML
- Export citation
The predictive role of symptoms in COVID-19 diagnostic models: A longitudinal insight
-
- Journal:
- Epidemiology & Infection / Volume 152 / 2024
- Published online by Cambridge University Press:
- 22 January 2024, e37
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
New Method Using Growth Dynamics to Quantify Microbial Contamination of Kaolinite Slurries
-
- Journal:
- Clays and Clay Minerals / Volume 61 / Issue 6 / December 2013
- Published online by Cambridge University Press:
- 01 January 2024, pp. 517-524
-
- Article
- Export citation
Helium as a Surrogate for Deuterium in LPI Studies
-
- Journal:
- Laser and Particle Beams / Volume 2023 / 2023
- Published online by Cambridge University Press:
- 01 January 2024, e2
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Implementation of a diagnostic stewardship intervention to improve blood-culture utilization in 2 surgical ICUs: Time for a blood-culture change
-
- Journal:
- Infection Control & Hospital Epidemiology / Volume 45 / Issue 4 / April 2024
- Published online by Cambridge University Press:
- 11 December 2023, pp. 452-458
- Print publication:
- April 2024
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Radiofrequency ice dielectric measurements at Summit Station, Greenland
-
- Journal:
- Journal of Glaciology , First View
- Published online by Cambridge University Press:
- 09 October 2023, pp. 1-12
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
We recently reported on the radio-frequency attenuation length of cold polar ice at Summit Station, Greenland, based on bi-static radar measurements of radio-frequency bedrock echo strengths taken during the summer of 2021. Those data also allow studies of (a) the relative contributions of coherent (such as discrete internal conducting layers with sub-centimeter transverse scale) vs incoherent (e.g. bulk volumetric) scattering, (b) the magnitude of internal layer reflection coefficients, (c) limits on signal propagation velocity asymmetries (‘birefringence’) and (d) limits on signal dispersion in-ice over a bandwidth of ~100 MHz. We find that (1) attenuation lengths approach 1 km in our band, (2) after averaging 10 000 echo triggers, reflected signals observable over the thermal floor (to depths of ~1500 m) are consistent with being entirely coherent, (3) internal layer reflectivities are ≈–60$\to$
 –70 dB, (4) birefringent effects for vertically propagating signals are smaller by an order of magnitude relative to South Pole and (5) within our experimental limits, glacial ice is non-dispersive over the frequency band relevant for neutrino detection experiments.
–70 dB, (4) birefringent effects for vertically propagating signals are smaller by an order of magnitude relative to South Pole and (5) within our experimental limits, glacial ice is non-dispersive over the frequency band relevant for neutrino detection experiments.

Implementation of diagnostic stewardship in two surgical ICUs: Time for a blood-culture change
-
- Journal:
- Antimicrobial Stewardship & Healthcare Epidemiology / Volume 3 / Issue S2 / June 2023
- Published online by Cambridge University Press:
- 29 September 2023, pp. s9-s10
-
- Article
-
- You have access
- Open access
- Export citation
-
Background: Blood cultures are commonly ordered for patients with low risk of bacteremia. Liberal blood-culture ordering increases the risk of false-positive results, which can lead to increased length of stay, excess antibiotics, and unnecessary diagnostic procedures. We implemented a blood-culture indication algorithm with data feedback and assessed the impact on ordering volume and percent positivity. Methods: We performed a prospective cohort study from February 2022 to November 2022 using historical controls from February 2020 to January 2022. We introduced the blood-culture algorithm (Fig. 1) in 2 adult surgical intensive care units (ICUs). Clinicians reviewed charts of eligible patients with blood cultures weekly to determine whether the blood-culture algorithm was followed. They provided feedback to the unit medical directors weekly. We defined a blood-culture event as ≥1 blood culture within 24 hours. We excluded patients aged <18 years, absolute neutrophil count <500, and heart and lung transplant recipients at the time of blood-culture review. Results: In total, 7,315 blood-culture events in the preintervention group and 2,506 blood-culture events in the postintervention group met eligibility criteria. The average monthly blood-culture rate decreased from 190 blood cultures per 1,000 patient days to 142 blood cultures per 1,000 patient days (P < .01) after the algorithm was implemented. (Fig. 2) The average monthly blood-culture positivity increased from 11.7% to 14.2% (P = .13). Average monthly days of antibiotic therapy (DOT) was lower in the postintervention period than in the preintervention period (2,200 vs 1,940; P < .01). (Fig. 3) The ICU length of stay did not change before the intervention compared to after the intervention: 10 days (IQR, 5–18) versus 10 days (IQR, 5–17; P = .63). The in-hospital mortality rate was lower during the postintervention period, but the difference was not statistically significant: 9.24% versus 8.34% (P = .17). The all-cause 30-day mortality was significantly lower during the intervention period: 11.9% versus 9.7% (P < .01). The unplanned 30-day readmission percentage was significantly lower during the intervention period (10.6% vs 7.6%; P < .01). Over the 9-month intervention, we reviewed 916 blood-culture events in 452 unique patients. Overall, 74.6% of blood cultures followed the algorithm. The most common reasons overall for ordering blood cultures were severe sepsis or septic shock (37%), isolated fever and/or leukocytosis (19%), and documenting clearance of bacteremia (15%) (Table 1). The most common indications for inappropriate blood cultures were isolated fever and/or leukocytosis (53%). Conclusions: We introduced a blood-culture algorithm with data feedback in 2 surgical ICUs and observed decreases in blood-culture volume without a negative impact on ICU LOS or mortality rate.
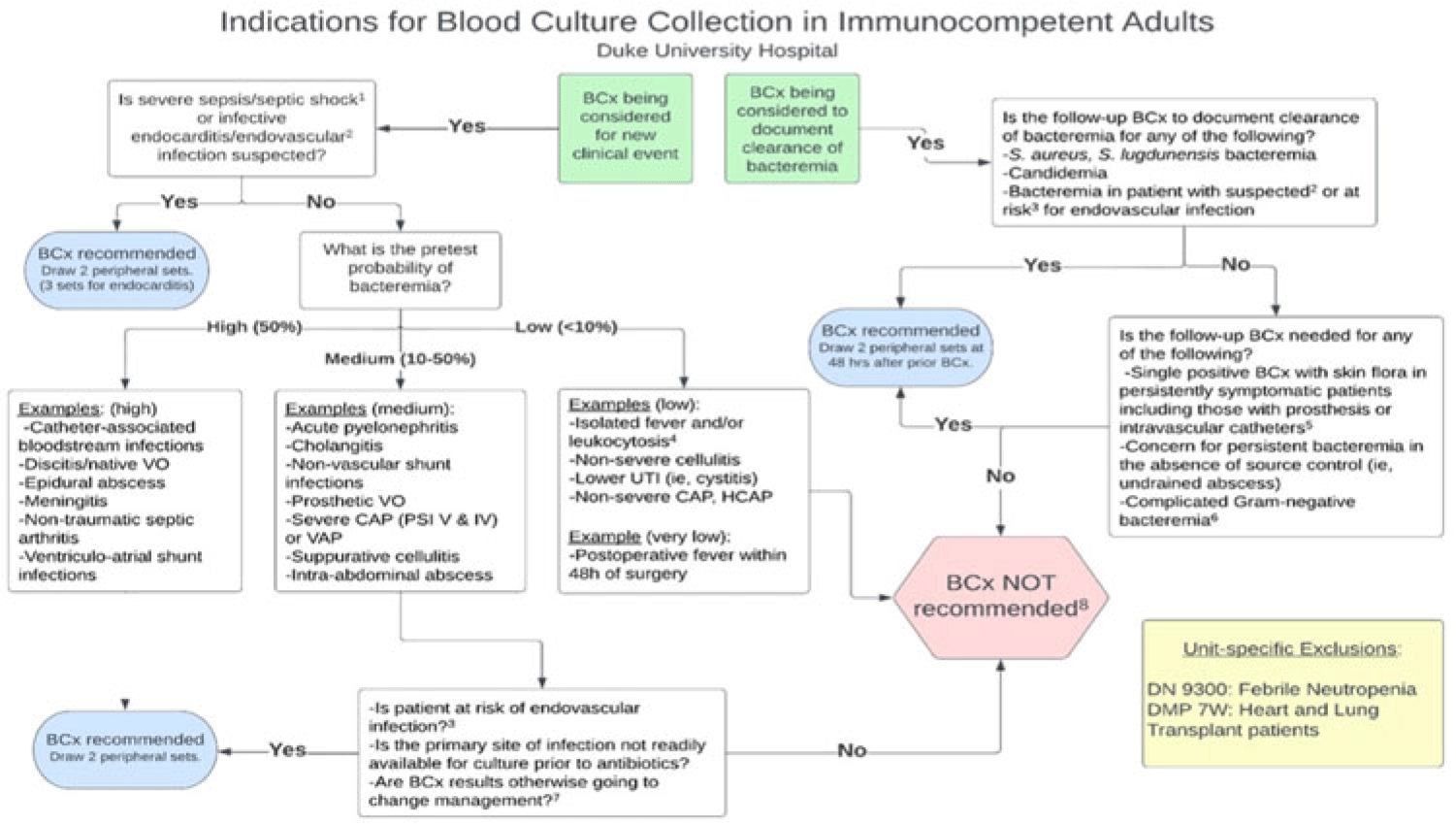
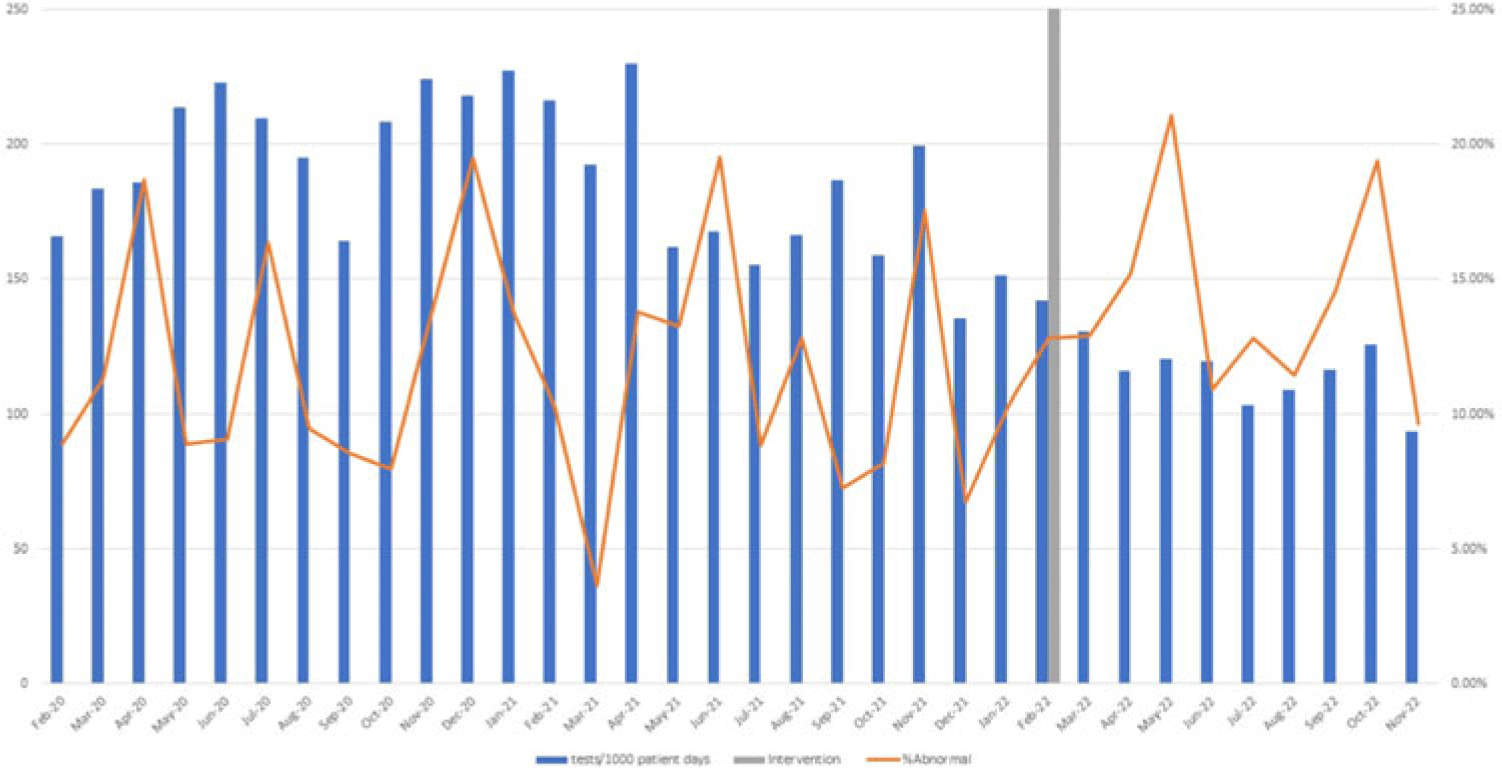
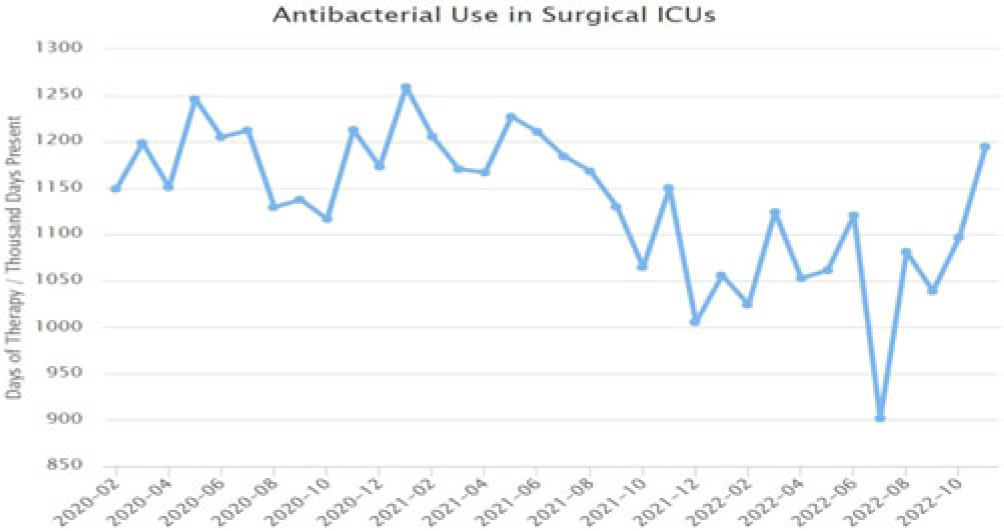
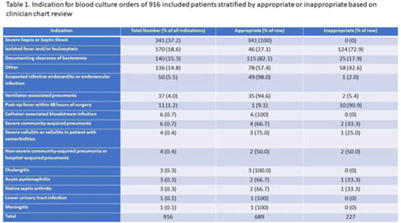
Disclosure: None




Epidemiology of carbapenem-resistant and extended-spectrum beta-lactamase-producing Enterobacterales in US children, 2016–2020
-
- Journal:
- Antimicrobial Stewardship & Healthcare Epidemiology / Volume 3 / Issue S2 / June 2023
- Published online by Cambridge University Press:
- 29 September 2023, p. s16
-
- Article
-
- You have access
- Open access
- Export citation
Investigation of the first cluster of Candida auris cases among pediatric patients in the United States―Nevada, May 2022
-
- Journal:
- Antimicrobial Stewardship & Healthcare Epidemiology / Volume 3 / Issue S2 / June 2023
- Published online by Cambridge University Press:
- 29 September 2023, pp. s118-s119
-
- Article
-
- You have access
- Open access
- Export citation
An approach for collaborative development of a federated biomedical knowledge graph-based question-answering system: Question-of-the-Month challenges
-
- Journal:
- Journal of Clinical and Translational Science / Volume 7 / Issue 1 / 2023
- Published online by Cambridge University Press:
- 14 September 2023, e214
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
The Rapid ASKAP Continuum Survey III: Spectra and Polarisation In Cutouts of Extragalactic Sources (SPICE-RACS) first data release
-
- Journal:
- Publications of the Astronomical Society of Australia / Volume 40 / 2023
- Published online by Cambridge University Press:
- 30 August 2023, e040
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
The Australian SKA Pathfinder (ASKAP) radio telescope has carried out a survey of the entire Southern Sky at 887.5 MHz. The wide area, high angular resolution, and broad bandwidth provided by the low-band Rapid ASKAP Continuum Survey (RACS-low) allow the production of a next-generation rotation measure (RM) grid across the entire Southern Sky. Here we introduce this project as Spectral and Polarisation in Cutouts of Extragalactic sources from RACS (SPICE-RACS). In our first data release, we image 30 RACS-low fields in Stokes I, Q, U at 25
 $^{\prime\prime}$ angular resolution, across 744–1032 MHz with 1 MHz spectral resolution. Using a bespoke, highly parallelised, software pipeline we are able to rapidly process wide-area spectro-polarimetric ASKAP observations. Notably, we use ‘postage stamp’ cutouts to assess the polarisation properties of 105912 radio components detected in total intensity. We find that our Stokes Q and U images have an rms noise of
$^{\prime\prime}$ angular resolution, across 744–1032 MHz with 1 MHz spectral resolution. Using a bespoke, highly parallelised, software pipeline we are able to rapidly process wide-area spectro-polarimetric ASKAP observations. Notably, we use ‘postage stamp’ cutouts to assess the polarisation properties of 105912 radio components detected in total intensity. We find that our Stokes Q and U images have an rms noise of 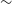 $\sim$80
$\sim$80 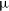 $\unicode{x03BC}$Jy PSF
$\unicode{x03BC}$Jy PSF $^{-1}$, and our correction for instrumental polarisation leakage allows us to characterise components with
$^{-1}$, and our correction for instrumental polarisation leakage allows us to characterise components with 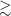 $\gtrsim$1% polarisation fraction over most of the field of view. We produce a broadband polarised radio component catalogue that contains 5818 RM measurements over an area of
$\gtrsim$1% polarisation fraction over most of the field of view. We produce a broadband polarised radio component catalogue that contains 5818 RM measurements over an area of 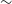 $\sim$1300 deg
$\sim$1300 deg $^{2}$ with an average error in RM of
$^{2}$ with an average error in RM of 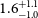 $1.6^{+1.1}_{-1.0}$ rad m
$1.6^{+1.1}_{-1.0}$ rad m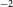 $^{-2}$, and an average linear polarisation fraction
$^{-2}$, and an average linear polarisation fraction 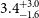 $3.4^{+3.0}_{-1.6}$ %. We determine this subset of components using the conditions that the polarised signal-to-noise ratio is
$3.4^{+3.0}_{-1.6}$ %. We determine this subset of components using the conditions that the polarised signal-to-noise ratio is 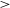 $>$8, the polarisation fraction is above our estimated polarised leakage, and the Stokes I spectrum has a reliable model. Our catalogue provides an areal density of
$>$8, the polarisation fraction is above our estimated polarised leakage, and the Stokes I spectrum has a reliable model. Our catalogue provides an areal density of 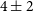 $4\pm2$ RMs deg
$4\pm2$ RMs deg $^{-2}$; an increase of
$^{-2}$; an increase of 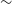 $\sim$4 times over the previous state-of-the-art (Taylor, Stil, Sunstrum 2009, ApJ, 702, 1230). Meaning that, having used just 3% of the RACS-low sky area, we have produced the 3rd largest RM catalogue to date. This catalogue has broad applications for studying astrophysical magnetic fields; notably revealing remarkable structure in the Galactic RM sky. We will explore this Galactic structure in a follow-up paper. We will also apply the techniques described here to produce an all-Southern-sky RM catalogue from RACS observations. Finally, we make our catalogue, spectra, images, and processing pipeline publicly available.
$\sim$4 times over the previous state-of-the-art (Taylor, Stil, Sunstrum 2009, ApJ, 702, 1230). Meaning that, having used just 3% of the RACS-low sky area, we have produced the 3rd largest RM catalogue to date. This catalogue has broad applications for studying astrophysical magnetic fields; notably revealing remarkable structure in the Galactic RM sky. We will explore this Galactic structure in a follow-up paper. We will also apply the techniques described here to produce an all-Southern-sky RM catalogue from RACS observations. Finally, we make our catalogue, spectra, images, and processing pipeline publicly available.


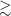
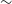

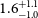
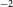
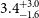
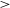

Advocacy at the Eighth World Congress of Pediatric Cardiology and Cardiac Surgery
-
- Journal:
- Cardiology in the Young / Volume 33 / Issue 8 / August 2023
- Published online by Cambridge University Press:
- 24 August 2023, pp. 1277-1287
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Disinfection efficacy of Oxivir TB wipe residue on severe acute respiratory coronavirus virus 2 (SARS-CoV-2)
-
- Journal:
- Infection Control & Hospital Epidemiology / Volume 44 / Issue 11 / November 2023
- Published online by Cambridge University Press:
- 19 July 2023, pp. 1891-1893
- Print publication:
- November 2023
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Assessing pig farm biosecurity measures for the control of Salmonella on European farms
-
- Journal:
- Epidemiology & Infection / Volume 151 / 2023
- Published online by Cambridge University Press:
- 13 July 2023, e130
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
12 - Becoming Political
- from Conclusion
-
-
- Book:
- Making the Middle Republic
- Published online:
- 20 April 2023
- Print publication:
- 27 April 2023, pp 253-269
-
- Chapter
- Export citation
Linear and nonlinear stability of Rayleigh–Bénard convection with zero-mean modulated heat flux
-
- Journal:
- Journal of Fluid Mechanics / Volume 961 / 25 April 2023
- Published online by Cambridge University Press:
- 11 April 2023, A1
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
Linear and nonlinear stability analyses are performed to determine critical Rayleigh numbers (
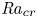 ${Ra}_{cr}$) for a Rayleigh–Bénard convection configuration with an imposed bottom boundary heat flux that varies harmonically in time with zero mean. The
${Ra}_{cr}$) for a Rayleigh–Bénard convection configuration with an imposed bottom boundary heat flux that varies harmonically in time with zero mean. The  ${Ra}_{cr}$ value depends on the non-dimensional frequency
${Ra}_{cr}$ value depends on the non-dimensional frequency 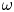 $\omega$ of the boundary heat-flux modulation. Floquet theory is used to find
$\omega$ of the boundary heat-flux modulation. Floquet theory is used to find 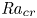 ${Ra}_{cr}$ for linear stability, and the energy method is used to find
${Ra}_{cr}$ for linear stability, and the energy method is used to find 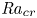 ${Ra}_{cr}$ for two different types of nonlinear stability: strong and asymptotic. The most unstable linear mode alternates between synchronous and subharmonic frequencies at low
${Ra}_{cr}$ for two different types of nonlinear stability: strong and asymptotic. The most unstable linear mode alternates between synchronous and subharmonic frequencies at low 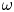 $\omega$, with only the latter at large
$\omega$, with only the latter at large 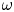 $\omega$. For a given frequency, the linear stability
$\omega$. For a given frequency, the linear stability 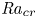 ${Ra}_{cr}$ is generally higher than the nonlinear stability
${Ra}_{cr}$ is generally higher than the nonlinear stability 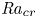 ${Ra}_{cr}$, as expected. For large
${Ra}_{cr}$, as expected. For large 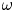 $\omega$,
$\omega$, 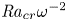 ${Ra}_{cr} \omega ^{-2}$ approaches an
${Ra}_{cr} \omega ^{-2}$ approaches an 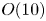 $O(10)$ constant for linear stability but zero for nonlinear stability. Hence the domain for subcritical instability becomes increasingly large with increasing
$O(10)$ constant for linear stability but zero for nonlinear stability. Hence the domain for subcritical instability becomes increasingly large with increasing 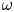 $\omega$. The same conclusion is reached for decreasing Prandtl number. Changing temperature and/or velocity boundary conditions at the modulated or non-modulated plate leads to the same conclusions. These stability results are confirmed by selected direct numerical simulations of the initial value problem.
$\omega$. The same conclusion is reached for decreasing Prandtl number. Changing temperature and/or velocity boundary conditions at the modulated or non-modulated plate leads to the same conclusions. These stability results are confirmed by selected direct numerical simulations of the initial value problem.
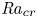

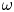
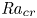
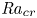
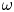
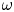
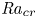
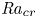
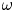
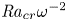
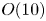
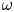
Using clinical decision support to improve urine testing and antibiotic utilization
-
- Journal:
- Infection Control & Hospital Epidemiology / Volume 44 / Issue 10 / October 2023
- Published online by Cambridge University Press:
- 29 March 2023, pp. 1582-1586
- Print publication:
- October 2023
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
