



Introduction
The mammalian ovary is composed of a follicular pool that contains thousands of preantral follicles, ∼99.9% of which never reach ovulation due to follicular atresia (Markström et al., Reference Markström, Svensson, Shao, Svanberg and Billig2002). In most species, these follicles are located in the outer portion of the ovarian tissue, called the cortical region.
Counting preantral follicles in mammalian ovaries provides an estimate of the follicular population. This type of study is laborious because all follicles present in the extension of the cortical region must be counted. Therefore, most studies use representative samples of ovarian tissue for this analysis and overlook the boundaries of the ovarian layers (cortical and medullary region; Schmidt et al., Reference Schmidt, Byskov, Nyboe Andersen, Müller and Yding Andersen2003; Stansfield et al., Reference Stansfield, Nöthling and Ansari2011).
The location and distribution of ovarian follicular populations have been studied in most domestic species. However, some authors found peculiarities and distinctions in relation to preantral follicles in mammals, such as in the canine species, in which a difference was observed in the number of preantral follicles between the right and left ovaries (Lunardon et al., Reference Lunardon, Silva-Santos, Justino, Dessunti, Seneda and Martins2015). In equine species, a new concept exists regarding the location of preantral follicles, which exhibited a greater quantity and integrity in the internal region, close to the ovulatory fossa (González et al., Reference González, da Silva, Lindquist, Búfalo, Morotti, Lisboa and Seneda2017). Furthermore, Alves et al. (Reference Alves, Alves, Gastal, Gastal, Figueiredo and Gastal2018) reported the influence of mare age, ovarian region, and portion on the spatial distribution of ovarian follicles, indicating that the follicular concentration was greater in the centre of the ovary of younger animals. In bovine species, a study showed a distinction in the population of preantral follicles between Bos taurus and Bos taurus indicus females (Silva-Santos et al., Reference Silva-Santos, Santos, Siloto, Hertel, Andrade, Rubin, Sturion, Melo-Sterza and Seneda2011) and a correlation between the preantral follicle population and antral follicle count (AFC); Bos taurus indicus cows with high AFC presented a lower population of preantral follicles (Silva-Santos et al., Reference Silva-Santos, Santos, Koetz Júnior, Morotti, Siloto, Marcantonio, Urbano, Oliveira, Lima and Seneda2014). Additionally, this study, conducted by Silva-Santos et al. (Reference Silva-Santos, Santos, Siloto, Hertel, Andrade, Rubin, Sturion, Melo-Sterza and Seneda2011), reported that multi-oocyte follicles were present in 41% of the evaluated ovaries of Nelore and Aberdeen Angus heifers. Multi-oocyte follicles are characterized by the presence of two or more oocytes within a single follicle without a basement membrane separating them (Tingen et al., Reference Tingen, Kim and Woodruff2009).
Nevertheless, in bovine species, despite the knowledge regarding the location of preantral follicles in the cortical portion of the ovary, little is known regarding the distribution of these follicles in this region (Cushman et al., Reference Cushman, Soares, Yake, Patterson, Rosasco, Beard, Northrop, Rich, Miles, Chase, Gonda, Perry, Mcneel and Summers2019; Tenley et al., Reference Tenley, Gomes, Rosasco, Northrop, Rich, Mcneel, Summers, Miles, Chase, Lents, Perry, Wood, Cupp and Cushman2019). Compared with species of similar size, such as horses, donkeys, and buffaloes, bovine species have ovaries with smaller dimensions and/or a greater number of follicles (Driancourt et al., Reference Driancourt, Paris, Roux, Mariana and Palmer1982; Mondadori et al., Reference Mondadori, Santin, Fidelis, Porfírio and Báo2010; Lopes et al., Reference Lopes, Lima, Bezerra, Barreto-Junior, Oliveira and Silva2017; Teh et al., Reference Teh, Izzati, Mori, Fuke, Hirai, Kitahara and Yamaguchi2018). Recent studies have shown that small and highly variable follicles can be collected from the cortical portion of the ovary (Bergamo et al., Reference Bergamo, Bonato, Bizarro-Silva, Bonato, González, Rossaneis, Verri, Morotti and Seneda2022; Cerezetti et al., Reference Cerezetti, González, Ferraz, Verri, Rabelo, Seneda and Morotti2021). Therefore, studies need to be conducted on the distribution of preantral follicles in bovine ovarian fragments.
A study on the distribution of ovarian preantral follicles in bovine species may benefit the biotechnique of manipulation of oocytes included in preantral ovarian follicles (MOIFOPA), as it will allow the location of the region with the greatest quantity and integrity of the follicles. The results from the MOIFOPA experiments will enable the elucidation of the mechanisms involved in the activation of primordial follicles and in the growth of primary and secondary follicles. In the future, the isolation of thousands of preantral follicles from a single ovary and subsequent in vitro culture (IVC) of the constituent oocytes until the maturation stage could contribute to the multiplication of animals of high zootechnical value or those threatened with extinction (Langbeen et al., Reference Langbeen, De Porte, Bartholomeus, Leroy and Bols2015; de Vasconcelos et al., Reference de Vasconcelos, Maculan, Da Cunha, Silva, Batista, Donato, Peixoto, Silva and De Souza2020).
These reports portray the growing interest of the scientific community in mapping bovine follicles in different ovarian areas so that the gaps in the subject are filled. In this study, we investigated the distribution of preantral follicles in the ovaries of bovines, considering the presence and integrity of follicles in the regions of the greater curvature of the ovary (GCO) and the region close to the ovarian pedicle (OP).
Materials and methods
Obtaining the ovaries
Ovaries (n = 12) were collected from Bos taurus indicus heifers, Nelore breed, with a body condition score (BCS) between 3 and 4 (scale 0–5; Ayres et al., Reference Ayres, Ferreira, de Souza Torres-Júnior, Demétrio, de Lima and Baruselli2009). The ovaries were obtained from a slaughterhouse 48 km away (40 min) from the Animal Reproduction Biotechnology Laboratory (ReproA-UEL). Ovaries were transported and stored in a thermal plastic container at 21ºC, in the absence of a buffer solution (González et al., Reference González, da Silva, Lindquist, Búfalo, Morotti, Lisboa and Seneda2017). Each ovary was carefully dissected with a scalpel to remove the surrounding connective tissue.
Experimental design
Ovaries obtained from slaughterhouses (n = 12) were used for investigating the regional distribution of the preantral follicles. These were individually identified and washed in three saline baths (0.9%; JP Farma, São Paulo, Brazil). Fragmentation of the ovaries was performed using a sterile and disposable dermatological punch (6 mm; Kolplast, São Paulo, Brazil), yielding fragments of ∼9 mm2. These fragments were defined in two regions that included the outer regions of the ovary: the region of the GCO and the region close to the OP, the insertion site of the utero-ovarian ligament. Two fragments were obtained from each region of the ovary (Figure 1). The weight of each ovary was measured (analytical balance, Bel®, Monza, Italy). The macroscopic count of all visible antral follicles (AFC) was determined for 12 ovaries of six heifers by the same person from the previously trained team. All antral follicles visible on the surfaces of both ovaries of each female were counted (Cushman et al., Reference Cushman, Allan, Kuehn, Snelling, Cupp and Freetly2009; Stojsin-Carter et al., Reference Stojsin-Carter, Mahboubi, Costa, Gillis, Carter, Neal, Miranda, Ohashi, Favetta and King2016; Rosa et al., Reference Rosa, Marinho, da Rosa, De Cesaro, Lunardelli, Silva-Santos, Basso, Bordignon and Seneda2018). The mean number of antral follicles and standard deviation were determined for the six pairs of ovaries, indicating the minimum and maximum variation in these follicles.
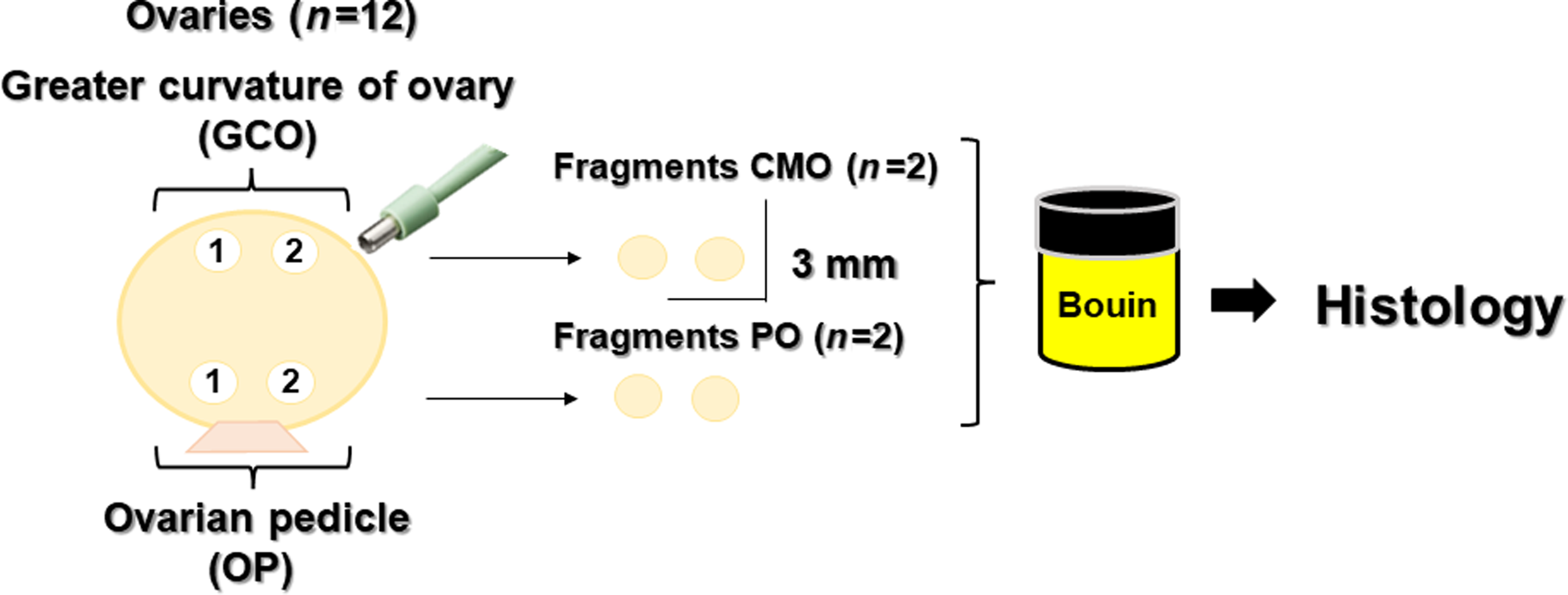
Figure 1. Experimental protocol describing the fragmentation of bovine ovaries according to the region: the greater curvature of ovary (GCO) and the region close to the ovarian pedicle (OP).
Histological process
The fragments were immediately fixed in Bouin’s solution for 24 h, kept in 70% ethanol for 24 h, dehydrated in increasing concentrations of ethanol, subjected to deparaffinization in xylene, embedded in paraffin and sequentially cut to a thickness of 5 µm using a microtome (Leica, Wetzlar, Germany) for mounting the blades. Five slides with three histological sections were prepared for each ovarian fragment. The sections were stained using periodic acid-Schiff (PAS) stain and haematoxylin. Histological slides were observed under an optical microscope (Nikon, Tokyo, Japan) at magnifications of ×20 and ×40.
Follicle classification
Follicles were classified according to their developmental stage as follows: primordial follicles (oocytes surrounded by a layer of flattened granular cells); developing follicles, comprising primary follicles (oocytes surrounded by a single layer of cuboid granulosa cells); and secondary follicles (oocytes surrounded by more than a complete layer of cuboid granulosa cells; Gomes et al., Reference Gomes, Lisboa, Silva, Max, Marino, Oliveira, González, Barreiros, Marinho and Seneda2015). They were also classified as intact or degenerated, according to the morphology of the oocyte and granulosa cells. Morphologically intact follicles had intact oocytes and granulosa cells arranged in discrete layers without a pycnotic core. Degenerated follicles were defined as follicles containing an oocyte with a pycnotic core and/or oocytes surrounded by disorganized granulosa cells detached from the membrane and with a retracted cytoplasm (Andrade et al., Reference Andrade, Marcondes Seneda, Alfieri, de Oliveira, Frederico Rodrigues Loureiro Bracarense, Figueiredo and Toniolli2005; Haag et al., Reference Haag, Magalhães-Padilha, Fonseca, Wischral, Gastal, King, Jones, Figueiredo and Gastal2013; Gomes et al., Reference Gomes, Lisboa, Silva, Max, Marino, Oliveira, González, Barreiros, Marinho and Seneda2015).
A multi-oocyte follicle is characterized by the presence of two or more oocytes within a single follicle, with granulosa cells around them. Multi-oocytes were classified according to their developmental stage into primordial, primary, and secondary oocytes, similar to follicles with a single oocyte (Tingen et al., Reference Tingen, Kim and Woodruff2009).
Statistical analysis
The weight and AFC of the ovaries were determined using the mean and standard deviation, indicating the minimum and maximum variation. The proportion of intact follicles was established from the total number of evaluated follicles (intact and degenerated) in each ovarian region studied, that is, GCO versus OP. The proportions of primordial, primary, and secondary follicles in the GCO and OP regions were analyzed using the chi-squared test and/or Fisher’s exact test. For descriptive analysis, data are presented as percentages (%). All analyses were performed using the Minitab® 18.1.1 statistical program, adopting a 5% probability level.
Results
The average weight of ovaries was 4.04 ± 0.32 g, with a minimum and maximum variation of 2.65 and 5.63 g, respectively. The mean AFC was 54.58 ± 3.55 follicles, with a minimum and maximum variation of 30 and 71 follicles, respectively.
Histological analysis enabled the comparison of the number of follicles in each region studied and the evaluation of their integrity. In total, 1123 follicles were visualized in the region of the GCO; 949 (84.5%) were primordial, and 174 (15.5%) were developing. The region close to the OP presented 1454 follicles, 1266 (87%) primordial follicles and 44 (12.9%) developing follicles. The region close to the OP showed the highest proportion of intact follicles in the primordial (P < 0.0001) and primary (P = 0.042; Figure 2) stages. However, for secondary follicles, the follicular proportion was similar between the regions close to the OP and GCO.
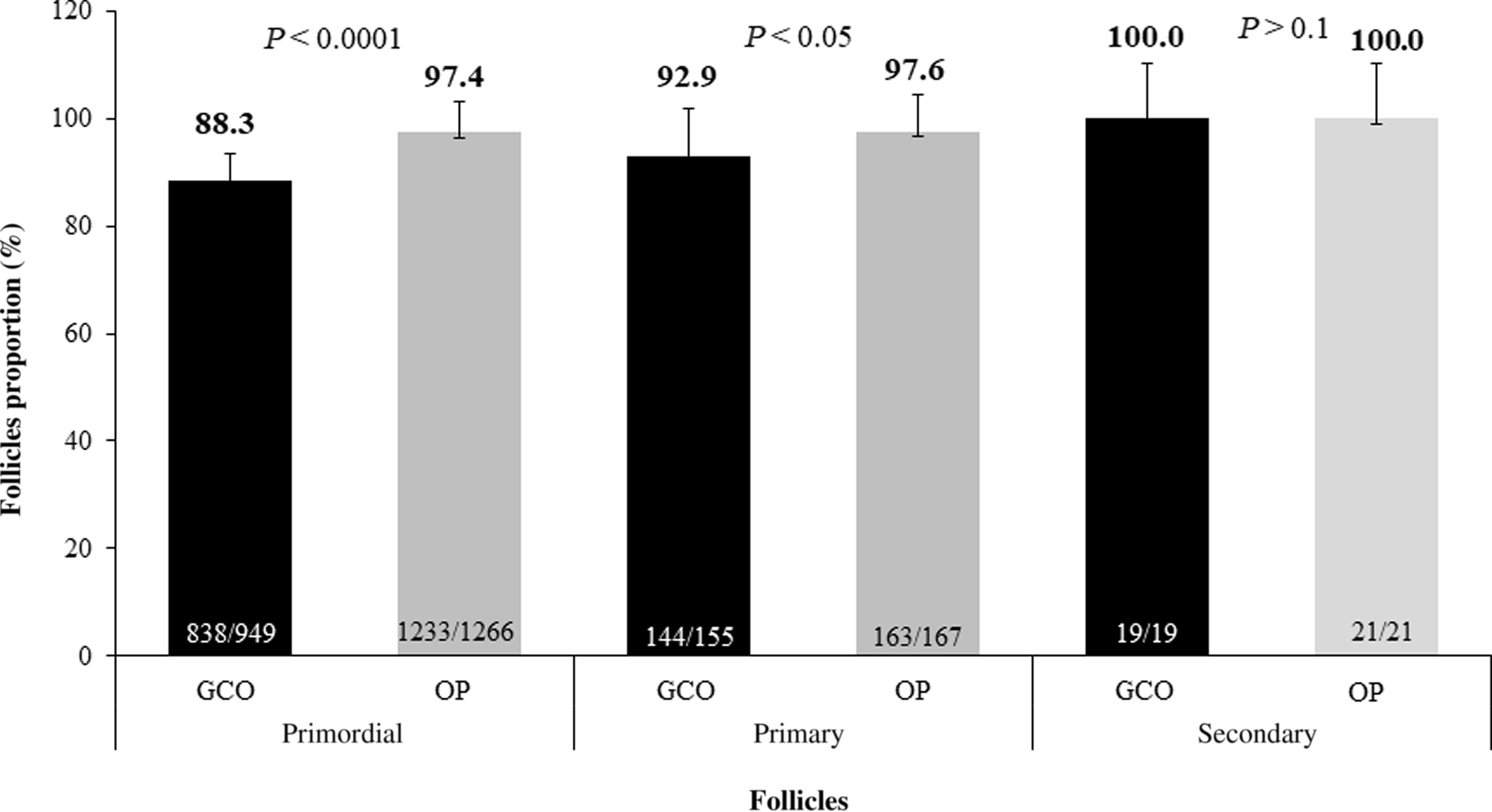
Figure 2. Total proportion of intact preantral follicles obtained from different regions of the bovine ovary, close to the greater curvature of ovary (GCO) or ovarian pedicle (OP).
Evaluation of the histological slides indicated the presence of multi-oocyte follicles in the ovaries of two bovine females (16%; 2/12). These were counted as a single follicle. All multi-oocyte follicles had organized granulosa cells and intact oocytes. Each multi-oocyte contained three oocytes, which were surrounded by only a layer of cuboidal granulosa cells, characterizing the primary developmental stage (Figure 3).

Figure 3. Bovine preantral follicles. (A) Primordial preantral follicle activated. (B) Primary multi-oocyte follicle with three oocytes. Periodic acid-Schiff (PAS) and haematoxylin staining, ×40 magnification.
Discussion
This study found that the primordial and primary follicles of bovines were distributed in greater quantities in the region close to the OP. Our results may be related to the topography of the ovarian surface formed during fetal development. The germinal epithelium, which is concentrated in the outer portion of the ovary, produces a reserve of female follicles. In adult animals, the ovary surface is continually altered by the expansion and regression of follicles and the corpus luteum, in addition to cell death and repair of the tunica of the superficial epithelium, which occurs at the point of rupture of the follicle wall during ovulation (Hartanti et al., Reference Hartanti, Hummitzsch, Bonner, Bastian, Irving-Rodgers and Rodgers2020). Therefore, we suggest that the greater number of follicles found in the OP may be related to the physiological events of the oestrous cycle, which occur repeatedly on the surface of this epithelium, causing a smaller number of preantral follicles to remain in the GCO.
After verifying the highest concentration of follicles in the OP region, we verified that the regional distribution of ovarian follicles in the bovine species was little known, with no similar publications available. This fact may be because the investigation is considered laborious, as all follicles in each investigated ovarian fragment must be counted. Therefore, few studies have been conducted on follicular distribution in mammalian species (Schmidt et al., Reference Schmidt, Byskov, Nyboe Andersen, Müller and Yding Andersen2003; Stansfield et al., Reference Stansfield, Nöthling and Ansari2011; González et al., Reference González, da Silva, Lindquist, Búfalo, Morotti, Lisboa and Seneda2017, Alves et al., Reference Alves, Alves, Gastal, Gastal, Figueiredo and Gastal2018). Studies on bovine preantral follicles have already determined the relationship between the maternal age of the female and the reserve of primordial follicles of her daughters (Tenley et al., Reference Tenley, Gomes, Rosasco, Northrop, Rich, Mcneel, Summers, Miles, Chase, Lents, Perry, Wood, Cupp and Cushman2019) and demonstrated the difference in ovarian reserve among various breeds (Angus, Brahma, and Brangus: Cushman et al., Reference Cushman, Soares, Yake, Patterson, Rosasco, Beard, Northrop, Rich, Miles, Chase, Gonda, Perry, Mcneel and Summers2019; Angus and Nelore: Silva-Santos et al., Reference Silva-Santos, Santos, Siloto, Hertel, Andrade, Rubin, Sturion, Melo-Sterza and Seneda2011). These investigations did not determine the distribution of preantral follicles and the region with the greatest presence of preantral follicles in the ovary of bovine females. In our study, we used ovarian fragments from a slaughterhouse as experimental units to obtain reliable results for the distribution of ovarian follicles and the region with the greatest presence of ovarian follicles in cattle. Despite the global importance of the bovine species, studies of follicular distribution in cattle are not present in the published literature.
The most recent studies on the distribution and location of preantral follicles were conducted on the ovaries of equine and elephant females. In these studies, the highest concentration of follicles was found in the inner portion and close to the ovulatory fossa (González et al., Reference González, da Silva, Lindquist, Búfalo, Morotti, Lisboa and Seneda2017; Alves et al., Reference Alves, Alves, Gastal, Gastal, Figueiredo and Gastal2018) or only in the outer portion of the ovary (cortical; Stansfield et al., Reference Stansfield, Nöthling and Ansari2011). Similarly, in our study, we found that the cortical region, close to the OP, contained a greater number of follicles compared with the GCO. Additionally, our study identified a difference in follicle integrity between the GCO and OP. The latter showed higher proportions of intact primordial and primary follicles [97.4% (1233/1266) and 97.6% (163/167), respectively]. This may be because, compared with the GCO region, the OP region had a greater interaction with neighbouring follicles and the production of paracrine factors such as hormones, growth factors, and gases that can affect follicular degeneration. (Araújo et al., Reference Araújo, Gastal, Figueiredo and Gastal2014).
This is an important finding for our data relative to the classic concept of follicular distribution only in the cortical region. We identified an irregular or heterogeneous distribution of bovine ovarian follicles, and this result contradicts the concept that these follicles can only be found in the cortical region. In addition, among mammalian species (equine, donkey, and buffalo), bovine species have smaller ovaries with a greater number of follicles (Driancourt et al., Reference Driancourt, Paris, Roux, Mariana and Palmer1982; Mondadori et al., Reference Mondadori, Santin, Fidelis, Porfírio and Báo2010; Lopes et al., Reference Lopes, Lima, Bezerra, Barreto-Junior, Oliveira and Silva2017; Teh et al., Reference Teh, Izzati, Mori, Fuke, Hirai, Kitahara and Yamaguchi2018). A high percentage of follicular integrity was observed when bovine ovaries were stored in a container at 21ºC and without buffer solutions for transport (48 km away from the laboratory). Histological processing was performed using the Bouin fixation protocol previously reported by our group; this procedure effectively maintained the integrity of preantral follicles in the bovine ovarian tissue (Santos et al., Reference Santos, Silva-Santos, Andrade, Lisboa, Schneider, Ciquini, Ferreira, Nóbrega Junior and Seneda2012).
After verifying the distribution of preantral follicles in two different regions of the bovine ovary, during the identification and follicular classification, we observed the presence of multi-oocyte follicles in the ovarian fragments of two bovine females. Due to the few published reports on multi-oocyte follicles in adult bovine females, in a complementary way to our study, the authors thought it pertinent to emphasize this fact. This finding corroborates the findings of Oliveira et al. (Reference Oliveira, Silva-Santos, González, Bizarro-Silva, Machado, Bracarense and Seneda2017), who observed multi-oocyte follicles at the early stages of ovarian development. They additionally reported that oocytes might show dominance among themselves and remain in different phases of the cell cycle.
Multi-oocyte follicles have been described in all phases of folliculogenesis, from primordial to pre-ovulatory, in both bitches and cows (Ireland et al., Reference Ireland, Scheetz, Jimenez-Krassel, Themmen, Ward, Lonergan, Smith, Perez, Evans and Ireland2008; Silva-Santos et al., Reference Silva-Santos, Santos, Siloto, Hertel, Andrade, Rubin, Sturion, Melo-Sterza and Seneda2011). The frequency of these follicles varies among species, within species, and among individuals. The proportion of multi-oocyte follicles in Bos taurus females ranges from 0.3 to 5.4% (Ireland et al., Reference Ireland, Scheetz, Jimenez-Krassel, Themmen, Ward, Lonergan, Smith, Perez, Evans and Ireland2008), for Bos taurus indicus females it is 41% (Silva-Santos et al., Reference Silva-Santos, Santos, Siloto, Hertel, Andrade, Rubin, Sturion, Melo-Sterza and Seneda2011). In our study on Bos taurus indicus females, the proportion of multi-oocyte follicles was 16%, which is lower than that reported in previous studies. However, the presence of these structures in adults and their involvement in folliculogenesis remain unclear.
Another pertinent data collection carried out in the study described the weight of the ovaries of bovine females, allowing an understanding of the proportions of the ovaries used to determine the distribution of preantral follicles. Information on the weight of the ovaries of adult bovine females indicated that the average weight was 10–20 g and there may still be variation in ovarian weight according to the phase of the oestrous cycle, age and breed. In our study, the mean ovary weight of Nelore heifers was lower (4.04 ± 0.32 g) compared with the above value. According to Sartorelli et al. (Reference Sartorelli, Carvalho, Bergfelt, Ginther and Barros2005), the ovaries of Nelore animals were smaller and lighter than that of the Angus breed. Woodruff and Shea (Reference Woodruff and Shea2011) reported that older females were more predisposed to ovarian plasticity because of the longer exposure of the ovarian parenchyma to follicular growth and formation and regression of the corpus luteum, resulting in changes in size and weight.
Our study used slaughterhouse ovaries, making it possible to observe and account for the AFC, generating data to contextualize the reproductive condition of bovine females. According to our results, the average AFC in the ovaries of Nelore heifers from the slaughterhouse was 54.58 ± 3.55 follicles (30 and 71 follicles, minimum and maximum variation, respectively). The results for females with low AFC (<31 follicles) corroborated the results obtained by Rosa et al. (Reference Rosa, Marinho, da Rosa, De Cesaro, Lunardelli, Silva-Santos, Basso, Bordignon and Seneda2018) for animals with high AFC (>92 follicles). The correlation of follicular reserve with the antral population has already been described by Silva-Santos et al. (Reference Silva-Santos, Santos, Koetz Júnior, Morotti, Siloto, Marcantonio, Urbano, Oliveira, Lima and Seneda2014), who reported that Bos taurus indicus females showed lower ovarian reserve but higher AFC than Bos indicus taurus animals.
In recent biotechnology research on in vitro culture of bovine preantral follicles, these follicles were found to be limited in ovarian fragments (Cerezetti et al., Reference Cerezetti, González, Ferraz, Verri, Rabelo, Seneda and Morotti2021; Bergamo et al., Reference Bergamo, Bonato, Bizarro-Silva, Bonato, González, Rossaneis, Verri, Morotti and Seneda2022). The broad scope of ovarian fragment collections may restrict the success of the in vitro growth of preantral follicles. Therefore, our study, which is the first on the distribution of bovine preantral follicles, presents relevant information to the scientific community. Additionally, the location of follicles has practical applicability, as it helps guide the removal of ovarian fragments for biotechnologies that use in vitro cultures of follicles, indicating that the ovarian region has the highest concentration and integrity of preantral follicles.
This study showed that most intact primordial and primary follicles were located close to the OP. Therefore, the distribution of preantral follicles in the bovine ovary directed the removal of ovarian fragments for MOIFOPA biotechnology, thereby contributing to the advancement of this technique.
Financial support
The authors thank the National Council for Scientific and Technological Development (CNPq) for their support in this study, grant 308460/2018-9.
Conflict of interest
The authors declare none.
Ethical standards
This study was conducted according to the standards of the Ethics Committee for Animal Experimentation of the University of Londrina, following the rules established by Law 1283 for ovaries obtained from slaughterhouses.



