



Introduction
Brittle stars (Ophiuroidea) are among the most common echinoderms in oceans worldwide, including the deep waters of the North Atlantic and Arctic basins (e.g. Piepenburg & von Juterzenka, Reference Piepenburg and von Juterzenka1994; Mayer & Piepenburg, Reference Mayer and Piepenburg1996; Piepenburg & Schmid, Reference Piepenburg and Schmid1996; Piepenburg, Reference Piepenburg2000; Smirnov et al., Reference Smirnov, Piepenburg, Ahearn and Juterzenka2014). They are often described to dominate megafaunal assemblages (Cusson et al., Reference Cusson, Archambault and Aitken2007; MacDonald et al., Reference MacDonald, Bluhm, Iken, Gagaev and Strong2010; Piepenburg et al., Reference Piepenburg, Archambault, Ambrose, Blanchard, Bluhm, Carroll, Conlan, Cusson, Feder and Grebmeier2011; Roy et al., Reference Roy, Iken and Archambault2015; Lacharité & Metaxas, Reference Lacharité and Metaxas2017) and they play important ecological roles in both inshore and offshore habitats (Geraldi et al., Reference Geraldi, Bertolini, Emmerson, Roberts, Sigwart and Connor2017; Volage et al., Reference Volage, Hamel and Mercier2021). Despite the growing number of exploratory surveys in northern Canada, brittle stars remain incompletely inventoried and poorly described.
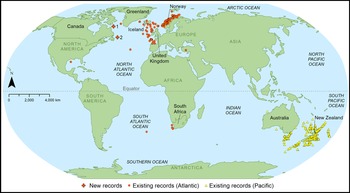
The brittle star Ophiactis abyssicola (M. Sars, 1861) is a morphologically variable species that has been reported from both hemispheres in the Pacific and Atlantic Oceans (Figure 1). However, records from the South Pacific around New Zealand and Australia (Stöhr et al., Reference Stöhr, O'Hara and Thuy2021) are most likely Ophiactis cuspidata Lyman, 1879, currently accepted as a subspecies of O. abyssicola, but representing a separate genetic clade (O'Hara et al., Reference O'Hara, England, Gunasekera and Naughton2014). In the Atlantic, O. abyssicola is reported from western Greenland (Smirnov et al., Reference Smirnov, Piepenburg, Ahearn and Juterzenka2014), Iceland and in other Scandinavian waters, around northern UK, western France, northern Spain, the Azores, Canaries, Cape Verde and the Gulf of Mexico (USA) (OBIS, 2021), as well as off South Africa (Mortensen, Reference Mortensen1933; Olbers et al., Reference Olbers, Griffiths, O'Hara and Samyn2019) and Tristan da Cunha (Lyman, Reference Lyman1882). Ophiactis abyssicola is known to occur on fine sand, gravel, shells and rocks, and among sponges, corals and other sessile organisms (Farran, Reference Farran1912; Stöhr & Segonzac, Reference Stöhr and Segonzac2005). In the North Atlantic, it has been found at depths ranging between 118–2107 m (Koehler, Reference Koehler1909; Stöhr & Segonzac, Reference Stöhr and Segonzac2005; Martynov & Litvinova, Reference Martynov and Litvinova2008).

Fig. 1. World map showing existing and new records of Ophiactis abyssicola in the Atlantic and existing records attributed to O. abyssicola in the Pacific, which are likely O. abyssicola cuspidata (O'Hara et al., Reference O'Hara, England, Gunasekera and Naughton2014). Sites 1 and 2 from eastern Canada represent new records off northern Labrador and off insular Newfoundland, respectively.
Despite this relatively broad distribution, O. abyssicola was conspicuously absent from records in Canadian waters (Figure 1). Two sampling campaigns conducted in deep waters off Newfoundland and Labrador confirmed its presence there, filling a major gap in the North Atlantic distribution of the species.
Methods
Specimens were collected along the continental shelf of eastern Canada in two regions. In Newfoundland, sampling occurred opportunistically during the annual multispecies bottom-trawl surveys operated by the Department of Fisheries and Oceans Canada (DFO) with the vessel CCGS Teleost in November 2013 inside the North Atlantic Fisheries Organization (NAFO) subdivision 3K. A Campelen 1800 shrimp trawl was used following a stratified-random design towed for 15 min for ~1.4 km (gear opened and closed at depth). Details of the type of gear and net can be found in Walsh & McCallum (Reference Walsh and McCallum1997). In Labrador, eight samplings (across seven stations) were carried out with the vessel CCGS Amundsen in September 2020 inside the NAFO subdivisions 2G and 2H. A rock dredge (mesh size of 7 mm) was deployed between 123–804 m depths while the ship moved at a maximum speed of 2 knots for a duration ranging between 5 and 10 min of bottom time.
Upon collection, brittle stars were sorted and preserved in 100% ethanol. The preserved specimens were examined under a dissecting microscope (M205A, Leica Microsystems, Singapore) and photographed (DFC7000T, Leica Microsystems, Wetzlar, Germany) with the software Leica Application Suite X, and identified morphologically using published keys and descriptions (Mortensen, Reference Mortensen1927; Paterson, Reference Paterson1985). A voucher specimen was deposited at the Canadian Museum of Nature (Ottawa) under catalog number CMNI 2021-0509.
For molecular identification, the mitochondrial cytochrome coxidase subunit I (COI) gene of one sample (September 2020) was sequenced using standard echinoderm primers (LCOech1aF1 TTTTTTCTACTAAACACAAGGATATTGG and HCO2198 TAAACTTCAGGGTGACCAAAAAATCA; Folmer et al., Reference Folmer, Black, Hoeh, Lutz and Vrijenhoek1994) and standard polymerase chain reaction (PCR) and DNA sequencing protocols (Ivanova et al., Reference Ivanova, Dewaard and Hebert2006; DeWaard et al., Reference DeWaard, Ivanova, Hajibabaei and Hebert2008). The sample was processed through the Canadian Centre for DNA Barcoding (University of Guelph) and compared with published sequences of specimens collected in the North Atlantic and around Norway on Barcode of Life Database (BOLD) and GenBank. We used BOLD ID engine, which assigns unknown sequences into BINS with other known sequences to obtain an identification (see Ratnasingham & Hebert, Reference Ratnasingham and Hebert2013 for details). The partial COI sequence with all meta-data were registered on BOLD, project AMLAB, and deposited in GenBank (accession number MZ364349).
Results
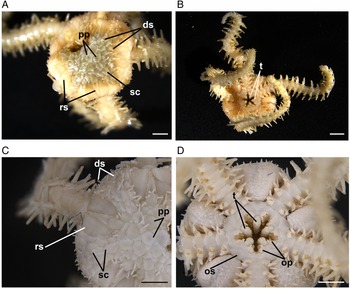
Two brittle star specimens were examined. The first was collected on 30 November 2013 along the northern coast of Newfoundland (49°58′00″N 49°45′24″W) at 1097 m depth and the other was collected on 2 September 2020 off northern Labrador (60°33′32″N 61°23′42″W) at 433 m depth (Figure 1). The specimens belong to the family Ophiactidae due to their tricuspid teeth (O'Hara et al., Reference O'Hara, Hugall, Stöhr, Thuy and Martynov2018). They concur with the descriptions of Ophiactis abyssicola by Mortensen (Reference Mortensen1927) and Paterson (Reference Paterson1985), and the COI results showed 100% similarity with records of O. abyssicola available in GenBank and BOLD. Main diagnostic characters observed in these specimens (~5 mm in diameter) are pentaradial symmetry, coarse dorsal disc scales with obvious primary plates, smaller ventral disc scales, conical spines on the dorsal and ventral disc, large (longer than half the disc radius), broad and separated radial shields, fan-shaped contiguous dorsal arm plates, three to four arm spines, rhombic oral shields, tricuspid teeth, and two lateral oral papillae, distal one larger than proximal one (Figure 2). Ophiactis abyssicola is a variable species, which is evidenced by the fact that six nominal species have been synonymized with it (Stöhr et al., Reference Stöhr, O'Hara and Thuy2021). Madeira et al. (Reference Madeira, Kroh, Cordeiro, Martins and Ávila2019) remarked that Ophiactis canotia (Lyman, 1879) from about 1830 m depth in the Azores is similar to O. abyssicola except for the outline of the dorsal arm plates. That species is only known from its two type specimens and has not been reassessed taxonomically, but it is highly likely that it is conspecific with O. abyssicola. The South Pacific subspecies O. abyssicola cuspidata that is currently unaccepted, cannot at present be separated morphologically from the Atlantic specimens, but molecular data have identified them as a distinct clade (O'Hara et al., Reference O'Hara, England, Gunasekera and Naughton2014). Ophiactis balli (Thompson, 1840) in the North Atlantic bears some resemblance to O. abyssicola, but its dorsal disc is covered by small scales and inconspicuous primary plates, it has a single lateral oral papilla, radial shields shorter than 1/4 disk radius, triangular oral shields and up to five arm spines (Paterson, Reference Paterson1985).
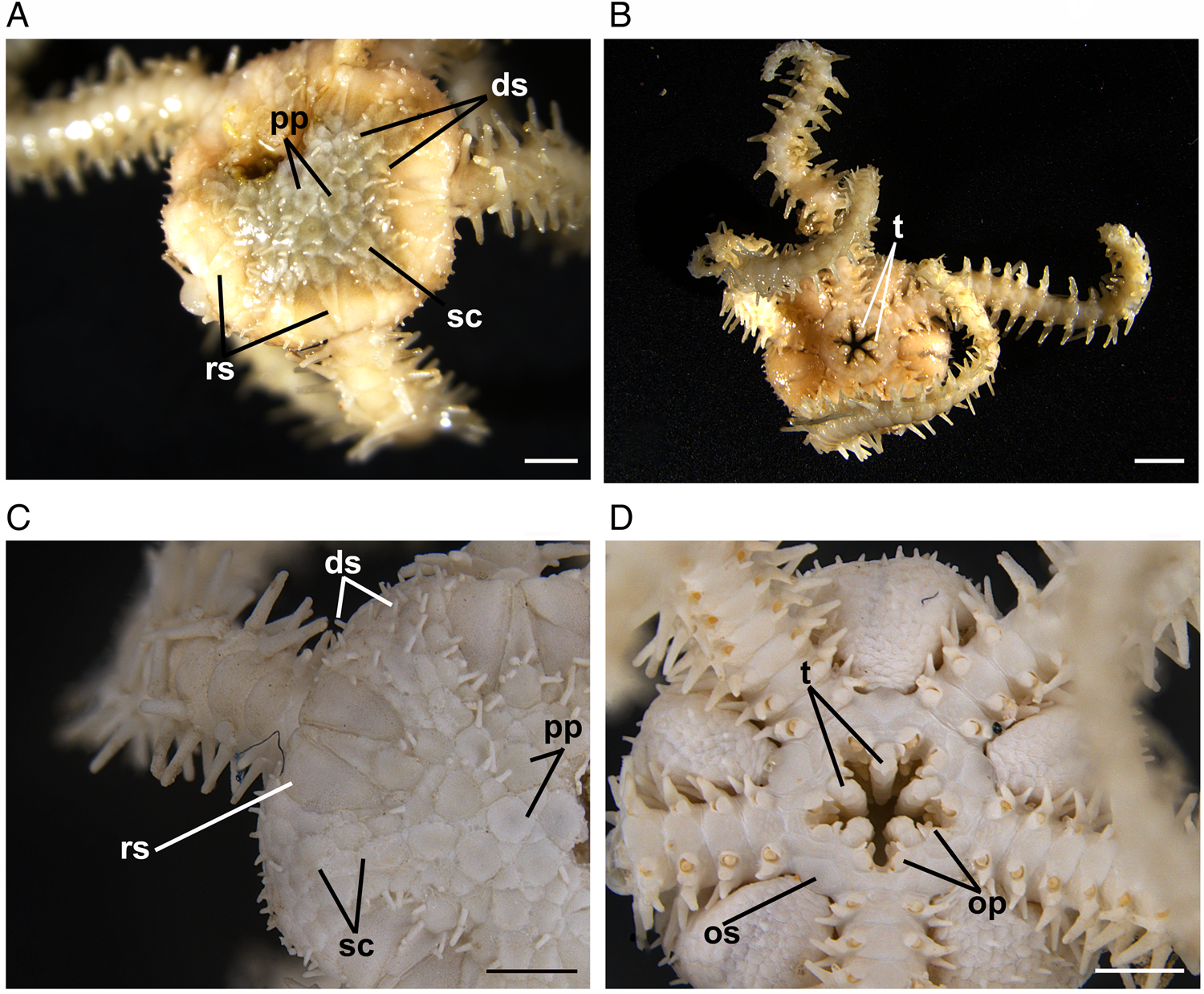
Fig. 2. Ophiactis abyssicola (from Labrador). (A) Dorsal view showing the primary plates (pp), radial shields (rs), disc scales (sc) and disc spines (ds). (B) Ventral view showing the teeth (t). (C) Dorsal view showing the disc spines (ds), disc scales (sc), primary plates (pp) and radial shields (rs). (D) Ventral view showing the oral shields (os), teeth (t) and oral papillae (op). Scale bars: (A, B) 1.5 mm; (C, D) 1 mm.
Discussion
The morphological characters of the specimens under study were consistent with previous descriptions of Ophiactis abyssicola, and the results of the COI analysis reinforced the morphological identification. The bathymetric distribution of O. abyssicola has been characterized as upper bathyal (shallower than 1800 m) by O'Hara et al. (Reference O'Hara, England, Gunasekera and Naughton2014), which corresponds to the collection depth of our specimens. However, the present records establish O. abyssicola for the first time in Canadian waters (Figure 1). Based on the closest site where the species was previously reported from (western Greenland; Figure 1), the present records extend its distribution range by about 500 km south-west in the Labrador Sea and about 1900 km south along the coast of northern Newfoundland (Figure 1). Due to sparse collecting effort on the continental slope in the study area (Coté et al., Reference Coté, Heggland, Roul, Fifield, Wareham, Colbourne, Maillet, Pilgrim, Pretty, Devine, Corre, Lawson, Yacos-Fuentes and Mercier2019), it is impossible to determine how common O. abyssicola is along the eastern Canadian seaboard or whether these specimens reflect a long-established distribution or recent range extension.
This newly recorded species of brittle star from the bathyal zone off Newfoundland and Labrador coasts adds to the marine biodiversity of Canada and to the general distribution knowledge of O. abyssicola.
Acknowledgements
We would like to thank the researchers and crew aboard CCGS Amundsen and CCGS Teleost for their help during the collections. We also warmly thank C.Pretty (DFO) for assistance with the map and two anonymous reviewers for their constructive comments.
Financial support
This research was partly funded by Discovery and Ship Time grants from the Natural Sciences and Engineering Research Council of Canada (NSERC) to A.M., with primary ship time on the CCGS Amundsen provided by DFO's ISECOLD program. Some of the data collected by the CCGS Amundsen were made available by Amundsen Science, which is supported through Université Laval by the Canada Foundation for Innovation (CFI) Major Science Initiatives (MSI) Fund.

 Open access
Open access