



Introduction
In most West African countries, including Mali, pearl millet is an important staple crop and is widely grown in the drought-prone Sahelian and Sudanian zones. The adoption of new varieties of pearl millet (Pennisetum glaucum L. R. Br.) in West Africa is low as it has been difficult to breed open-pollinated varieties that are clearly superior to well-adapted local landraces and improved varieties based on them. Instead of trying to breed better open-pollinated varieties, new hybrid varieties could be bred that farmers would adopt because they yield more while retaining other desirable traits. Single-cross hybrids (pure-line seed parent crossed with pure-line restorer parent) of Indian origin have been tested repeatedly in West Africa. They have proved to be highly vulnerable to downy mildew disease in Africa, where the disease originated. Breeding single-cross hybrids using African genotypes could give better results in the short term. However, because of the much stronger disease pressure in Africa compared with India, it is unlikely to give durable resistance, particularly as Indian single-cross hybrids have suffered from downy mildew epidemics (Singh et al., Reference Singh, King and Weraer1993). Breeding topcross hybrids rather than single-cross hybrids is a less conventional approach but is more likely to result in durable disease resistance. We produced hybrids by crossing a partially inbred male-sterile population (seed parent) and a partially inbred (open-pollinated) restorer population (pollinator) where both populations were based on locally adapted germplasm. Although a topcross hybrid is usually based on an inbred (pure-line) seed parent and a non-inbred pollinator, we describe these hybrids as topcrosses because, although the seed parent is not a pure line, it is more inbred than the pollinator. The partial inbreeding permits genetic heterogeneity for downy mildew resistance alleles that makes it, in theory and hence almost certainly in practice, possible to breed varieties with durable resistance across locations (Witcombe and Hash, Reference Witcombe and Hash2000). It is also practicable to preserve the resistance indefinitely through maintenance breeding because it is quite simple to introgress new sources of resistance into the parents of the topcross hybrids. It is straightforward to do so for the male parent and, with the additional effort of prior backcrossing into a sterile cytoplasm, it is also possible for the female parent.
Drought is one of the major yield-limiting factors in pearl millet, and terminal drought has a much greater impact than mid-season drought (Bidinger et al., Reference Bidinger, Mahalakshmi and Rao1987). Improved water uptake through better developed roots will reduce the impact of terminal drought, so we examined root development under well-watered and stressed conditions in a range of genotypes that included potential topcross parental lines.
The roots of pearl millet have been little studied. Chopart (Reference Chopart1983) found pearl millet root growth can be as much as 2 m in sandy soil with the roots growing rapidly, Azam-Ali et al. (Reference Azam-Ali1984) studied the roots of pearl millet at three row spacings, Brück et al. (Reference Brück, Sattelmacher and Payne2003) examined pearl millet roots under ridging and traditional sowing and Passot et al. (Reference Passot, Gnacko, Moukouanga, Lucas, Guyomarc’h, Ortega Beatriz, Atkinson, Belko Marème, Bennett, Gantet, Wells, Guédon, Vigouroux, Verdeil, Muller and Laplaze2016) examined the morphology of roots in early-stage development. Zegada-Lizarazu and Iijima (Reference Zegada-Lizarazu and Iijima2005) found that pearl millet was the best of six millet species in deep water uptake ability. The only authors that examined genotypic differences were Kusaka et al. (Reference Kusaka, Garcia Lalusin and Fujimura2005) but because the genotypes are only described as accessions from International Crops Research Institute for the Semi-Arid Tropics (ICRISAT) no conclusions can be drawn that relate to their adaptation or breeding history. In other crops, genetic variation in rooting ability has been found, for example, in sorghum (Salih et al., Reference Salih, Ali, Lux, Luxova, Cohen, Sugimato and Inanaga1999), in rice (O’Toole and Bland, Reference O’Toole and Bland1987), in groundnut (Azam-Ali, Reference Azam-Ali1984) and in wheat (Manschadi et al., Reference Manschadi, Christopher, de Voil and Hammer2006).
Topcross hybrids made by hand pollination were tested at Cinzana Experimental Research Station in Mali in the Sahelian zone to identify potential parental lines. Several, including selected lines from the root study, were converted to male-sterile populations and simultaneously restorer populations were bred. Finally, two topcross hybrids were tested by farmers in their fields in comparison to their local variety. We discuss current seed production efforts and the constraints to the commercialisation in Mali of topcross hybrids.
Materials and Methods
Root growth experiment
Plants of nine genotypes (Table 1) were grown in pots in a glasshouse at the ICRISAT, Patancheru, India from February to May 2010. Each pot contained 10 kg of soil in the ratio of five parts clay: three parts compost: one part sand. Three seeds were sown per pot and thinned to a single plant per pot after emergence. The pots were arranged in a split plot design with two water regimes as the main plot (with and without drought stress) and genotypes as the sub-plots. There were 5 replications (total of 90 pots: 2 water regimes × 9 genotypes × 5 replications).
Table 1. The genotypes in the root growth experiment with their pedigree and origin

* Station de Recherche Agronomique (SRA) Cinzana.
† Haryana Agricultural University, India.
†† International Crops Research Institute for the Semi-Arid Tropics.
For the first 30 days, all of the pots were well watered with the same quantity of water. In the evening of the 31st day (27th April), the pots were watered to saturation and they remained saturated until the next morning when the pots were sealed in polythene bags using paper clips at the base of the shoots and weighed to determine the initial weight. Subsequently, the 45 pots in the non-stressed regime were well watered every day. For the stress treatment, each pot was weighed to determine the water lost and no water was added until at least 20% of the water was lost. When the water loss was >20%, water was added sufficient to bring the pot to 80% of the initial weight. This applied a progressively increasing drought stress on the plants. All the plants in the pots were harvested on the 11th May, the day after the drought-stressed pots transpired less than 10% of the water in the well-watered regime.
At the end of the experiment, the roots were harvested. In a laboratory designed for the purpose, the roots were carefully washed with a water jet to remove all the soil and sealed in labelled plastic bags. They were then stored in a refrigerator at 4°C before evaluation. Thereafter, roots were rewashed, cut into large pieces and spread out, until completely separated, on labelled plastic trays containing water. The roots were scanned with a WinRHIZO (Regent Instruments Inc, Quebec, Canada) that determined the total root length and average root diameter, and which then calculated the total root surface and the total volume of the roots. After scanning, the roots were dried for at least 72 h at 120°C and then weighed on an electronic balance.
Statistical analyses
For the root traits, the differences between the environments, the genotypes and their interaction were determined by analysis of variance for all the traits using the aov function in R. From the analyses of variance, the entry mean repeatability, also known as entry mean heritability, was computed for each trait. It indicates the proportion of genetic variation over the total phenotypic variation. It is simply calculated as 1-(MSg-MSE), where MSg is the mean square for genotypes and MSE is the mean square for error (Fehr, Reference Fehr1991).
The performance of the genotypes across the well-watered (ww) and drought stress (ds) environments was determined by a regression analysis using environmental means.
A drought response for the nine lines was calculated as:
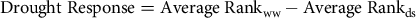 $$\rm {Drought \;Response = Average \;Rank_{ww} -Average \;Rank_{ds}}$$
$$\rm {Drought \;Response = Average \;Rank_{ww} -Average \;Rank_{ds}}$$
where the Average Rank was over five traits (diameter, length, area, volume and dry weight). The numerical values for these continuously variable traits were converted to ranks where 1 = best and 9 = worst. A positive Drought Response means a genotype is adapted to drought as it is better ranked in drought than under well-watered conditions. Ranking is necessary to remove large differences in the means of the traits across the two environments and the large differences in the means of the traits in a single environment.
The on-station and on-farm trials were also analysed using the aov function in R.
Conversion of the lines to male-sterile populations and the creation of restorer populations
Most of the lines in the root experiment were converted to female parents by a breeding programme at Cinzana Research Station, Mali. Five backcrosses were made into an A4 male-sterile cytoplasm with plant-by-plant crosses and the population continued only from pairs where the B (maintainer line) maintained sterility on the A (male-sterile line). At the end of the programme, the B lines were bulked to make a maintainer population. Simultaneously, with this programme, male populations were selected that restored fertility on the male-sterile lines. The programme was conducted from 2009 to 2012 and breeding work on the restorer populations continued until 2018 to select for improved flowering synchrony between the female and male parents, while also selecting for downy mildew resistance and restoration ability.
Performance of topcross hybrids on a research station
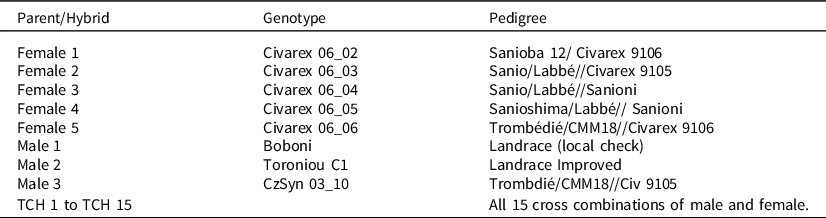
The experiment consisted of 5 female parents and 3 male parents and the 15 possible topcross hybrids on these parents (Table 2). The hybrid seed was produced by hand pollination as the experiment was conducted before the lines had been converted to male-sterile and restorer populations. The experiment was laid out in a Randomized Complete Block Design with four replications and with border plots that were discarded at the SRA Cinzana, Mali situated in the Sahelian zone of the country. Each plot consisted of two 5-m-long rows, with 0.5 m between rows and 50 cm between hills.
Table 2. The parents and topcross hybrids with their pedigree that were grown in SRA Cinzana, Mali in 2008
The experiment was sown on the 11 July 2010 in plots of 5 m row length with 0.75 m between rows and 0.5 m between plants. Plants were thinned to two plants per hill and the experiment was weeded twice at 15 and 25 days after sowing and earthing up was done 45 days after sowing. Diammonium phosphate (18-46-0) was applied at sowing at the rate of 100 kg ha−1 and 50 kg ha−1 of urea (46%) was applied after the first weeding. No fungicide was applied.
The downy mildew incidence was measured 60 days after emergence as the percentage of hills with a plot where there were symptoms of downy mildew on any plant. Although the results are only briefly referred to in this paper, stay green was also recorded as the percentage of leaves that remained green 7 weeks after flag leaf emergence.
All the mature panicles from each plot were harvested, threshed, dried in the sun until a constant weight and the weight of each panicle was measured. Data were analysed to estimate the best hybrid combinations.
Performance of topcross hybrids on farm
Two TCH (top cross hybrids), Toroniou TCH (CivarexA4 06_05/Toroniou C1) and Maiwa TCH (CivarexA4 06_05/Maiwa C1), were compared in farmers’ fields against a local variety (the farmers’ best variety) with 10 farmers in a cooperative in Koro in dispersed fields in 2016. Only data from seven farmers were available either because of late sowing or grazing damage. Koro is located in the Sahelian zone and is, incidentally, the region from where one of the trial entries, NKK (Niou Konio de Koro), originated. Each plot was 10 × 20 m and a standard dose of fertiliser was applied (diammonium phosphate at 100 kg ha−1 and 50 kg ha−1 urea) split equally between sowing time and the booting stage. Two sets of observations were made by interviewing farmers. The first was pre-harvest and farmers were asked about downy mildew incidence at maturity, grain yield potential and their overall appreciation of the variety compared with the local control. The second set, after harvest, asked about grain size, grain yield and again about losses caused by downy mildew. For all traits, the questions were asked relative to the performance of the local control. The yields of the harvested plots were weighed using a precision balance by researchers.
Results
Root trait experiment
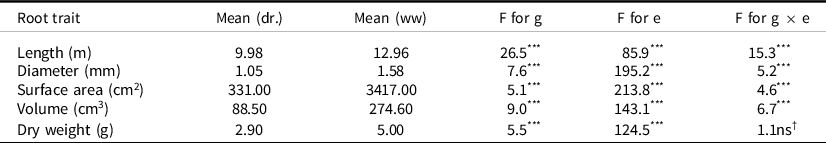
For all root traits, there were significant differences between genotypes and between the two watering regimes (Table 3). There were also significant genotype environment interactions for all of the traits other than for dry weight. The repeatability, also known as the entry mean heritability, was computed from the analyses of variance. Among the five parameters, root length, root surface area and root volume had the highest entry mean heritabilities (root length 0.96, surface area 0.80 and volume 0.89) so those characters could be selected for with a reliable predicted genetic gain. In single environments, the entry mean heritabilities for these three traits were higher in the droughted environment than in the well-watered.
Table 3. Mean length, diameter, surface area, volume and dry weight of roots under drought (dr.) and well-watered conditions (ww) in a glasshouse trial at ICRISAT, and the significance of genotype (g) effects, environment effect (e) and genotype × environment interactions (g × e) in an across treatment ANOVA
*** p ≤ 0.001.
† ns = non-significant.
Many of the traits had high correlations with each other so they cannot be independently selected for in a breeding programme (Table S1). In general, the correlations were higher within each of the watering regimes and lower across them. As expected, the highest correlations were for traits that were dependent on each other. Hence, root area and root volume were derived from diameter and root length and these traits showed generally high correlations – the lowest of these being 0.47 between diameter and surface area in drought stress. Across watering regimes, dry weight, a commonly measured root trait in the literature, had the highest correlations with diameter, area and volume, although not with length (Supplementary Table S1).
The degree of significance of g and g × e was highest for root length (Table 3). For this trait, the response of the genotypes to stress differed greatly (Figure 1) with one group, Civarex 06_05, H77 and NKK, having longer roots under drought conditions by 14% on average, whereas most of the entries had reduced root length by 58% on average (Figure 1). Similar trends were observed for root surface area and volume. Under drought, three genotypes, Civarex 06_05, H77 and NKK, had an increase in root surface area of 16% and root volume of 7% compared with well-watered conditions. In all other entries, on average, the root surface area was reduced by 65% and root volume by 109%.
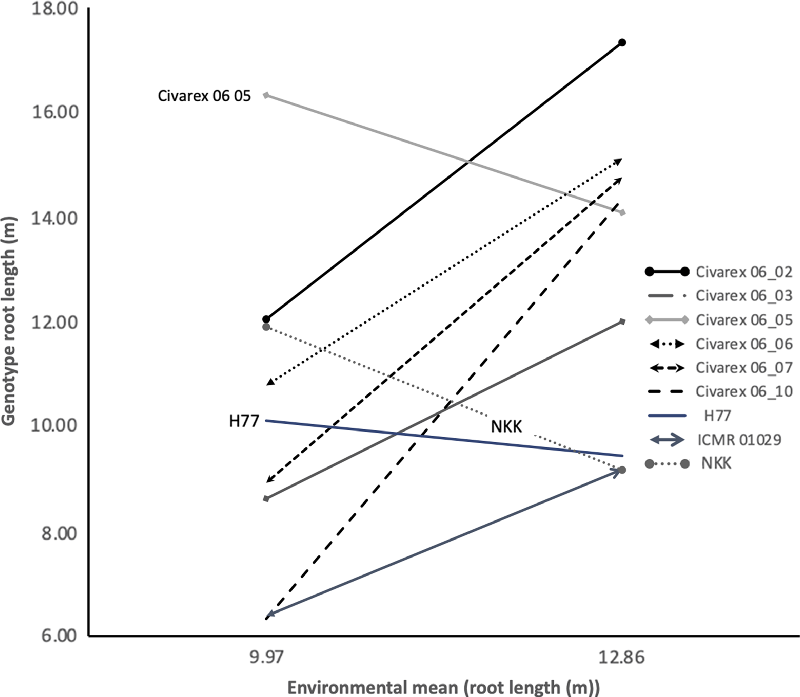
Figure 1. The response of the nine genotypes for root length from water stress environments (environmental mean 9.97 m) and well-watered environments (environmental mean 12.86 m). The three genotypes with positive drought responses are labelled.
Adaptation to drought conditions can be seen by the change in average rank for five root traits from well-watered to stress conditions (Table 4). Five genotypes showed a positive response (NKK, Civarex 06_05, H77, Civarex 06_07 and Civarex 06_02). Of these five, Civarex 06_05 and Civarex 06_02 were better ranked than NKK and H77 in both the water stress and well-watered environments. Four genotypes had negative drought responses as they had better developed roots in the well-watered environments than under stress (Table 4).
Table 4. Average ranking of the genotypes (1 = best and 9 = worst) of the nine genotypes for five root traits (length, diameter, area, volume and dry weight) in the droughted and well-watered environments and their drought response. The genotypes are ordered according to their drought response
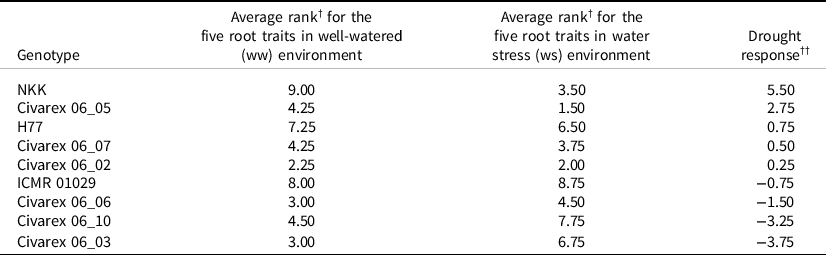
† Ranking done so that the genotype with the longest roots, largest diameter, surface area, volume and weight was ranked 1.
†† Drought Response = Average Rankww − Average Rankws (see Materials and Methods).
Performance of the topcross hybrids on-station
In the field trial, the topcross hybrids showed heterosis for yield and downy mildew resistance (Table 5). All the entries flowered within 1 or 2 days of each other. Although the mean heterosis of the hybrids was modest over the mean of the females, the best topcross hybrids were much higher yielding and had superior resistance to downy mildew. The two highest yielding topcross hybrids (Table 5) had Civarex 06_05, which had the most drought-adapted root traits, as the female parent.
Table 5. Yield, downy mildew incidence and flowering time in 15 topcross hybrids and their three male and five female parents, and the two highest yielding hybrids, SRA Cinzana, 2008, Mali
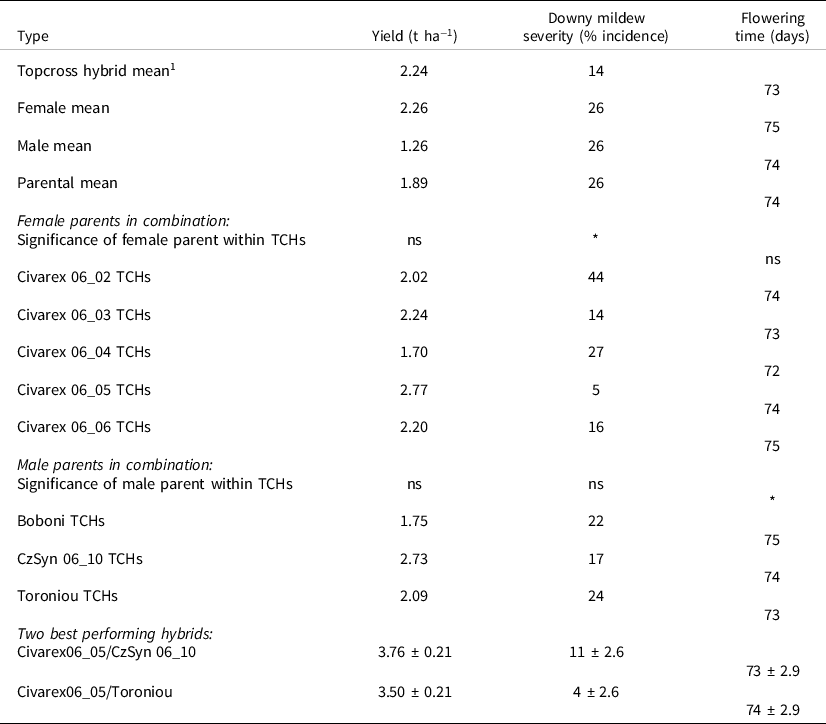
1 ***p ≤ 0.001 for differences among the 15 topcross hybrids for all three traits. ***p ≤ 0.001 for differences among all 23 genotypes for grain yield and downy mildew incidence, non-significant (ns) for flowering time.
The breeding and the performance of topcross hybrids at Koro
The on-station yield trial was based on topcross hybrids that were made by hand pollination as it was conducted before any lines were converted to male-sterile or restorer populations. This trial identified two high-yielding hybrids with male parents Toroniou and CzSyn 06_10 (both on female parent Civarex 06_05).
To make a restorer population, individual plant-by-plant crosses are made from the original population, for example, Toroniou, to male-sterile plants having the A4 cytoplasm. Only the plants that restored fertility are then included in the restorer population. For Toroniou, about 1000 crosses were made to identify 14 plants that were restorers and which were then recombined to make the initial restorer population (named as Toroniou R). However, it proved impossible to convert CzSyn 06_10 because no restorer plants were found in this population. However, population Maiwa from Nigeria was easy to convert as it was found to have a high frequency (about 20%) of plants that restored fertility. This resulted in two hybrids with commercial potential on seed parent Civarex 06_05 that had the restorer populations of Toroniou and Maiwa as the male parents.
Although there were only seven farmers that reported results, the results were highly significant as the topcross hybrid on Toroniou yielded 2.43 t ha−1 which was 72% more than the local check at 1.54 t ha−1 (p < 0.001, see Supplementary Table S2). The superiority of the hybrid on male-parent Maiwa was much less at 9%. The topcross hybrids were also considerably more resistant to downy mildew as the incidence on the Toroniou TCH was 7%, on the Maiwa TCH 10%, but on the local checks 33% (p < 0.001).
The farmers’ perceptions of the varieties confirmed the higher yield and disease resistance of the hybrids (Supplementary Table S3). Farmers also reported that the hybrids were more drought tolerant and stayed green at maturity compared with the local varieties. The local checks had the advantage of a larger grain size and longer panicles than both topcross hybrids, while the Toroniou TCH had the advantage of maturing earlier. Overall, despite its smaller grains and panicles, the farmers much preferred the Toroniou TCH to either the local variety of the Maiwa TCH which had the disadvantage of late maturity and a much smaller yield advantage.
Discussion
The effects of drought stress on root traits such as root length, surface area and volume separated the genotypes into two groups: a drought-adapted group comprised of Civarex 06_05, H77 and NKK, which responded positively to stress, and a drought intolerant group where the root traits under drought were greatly inferior (Table 4). Increased root growth under water stress has been reported in sunflower by Tahir et al. (Reference Tahir, Imran and Hussain2002) and in legumes by Yamauchi et al. (Reference Yamauchi, Paradales, Kono, Ito, Katayama, Johansen and Kumar Rao1996), and it allows the genotypes to exploit more of the residual moisture in the soil.
A finer root diameter was recognised by Bchini et al. (Reference Bchini, Daaloul and Saya2002) as being important for water stress adaptation in durum wheat as it enables better extraction of water and thus limits the consequences of severe drought during critical phases of plant development. NKK that originated from Koro, one of the driest prefectures in Mali, did have a below average root diameter under drought. However, Civarex 06_05 had a different adaptive mechanism as it had the largest root diameter of all genotypes under stress. However, both NKK and Civarex 06_05 had the lowest reduction in diameter from well-watered to drought conditions (14 and 21%, respectively, compared with an average reduction of 33%) and hence it is a trait worth considering for a marker-assisted selection programme.
Although data are not reported in this paper, the stay green trait was also assessed. Under two agro-climatic zones (Sahelian and Sudanese), Civarex 06_05, Civarex 06_06 and NKK were distinguished from all other genotypes that failed to stay green. More generally, genotypes with a better adaptation to drought had stay green trait at physiological maturity. Similar results with stay green have been reported by various authors: Vadez et al. (Reference Vadez, Krishnamurthy, Kashiwagi, Kholova, Devi, Sharma, Bhatnagar-Mathur, Hoisington, Hash, Bidinger and Keatinge2007) and Kamal et al. (Reference Kamal, Gorafi, Abdelrahman, Abdellatef and Tsujimoto2019) found a correlation between stay green trait and root depth in sorghum and Jordan et al. (Reference Jordan, Hunt, Cruickshank, Borrrell and Henzell2012) and Kamal et al. (Reference Kamal, Gorafi, Abdelrahman, Abdellatef and Tsujimoto2019) found a correlation between stay green trait and the proliferation of the root system in cereals.
The promise of Civarex 05_06 for hybrid breeding
The use of Civarex 06_05 as a male-sterile population for topcross hybrid breeding is promising as it has good root characteristics under drought, good general and specific combining ability, and a high and stable male sterility in the A4 cytoplasm. Civarex 06_05 also has the stay green trait. This was confirmed in experiments (data not presented in this paper) where chlorophyll a and b contents were measured under drought stress.
In India, single-cross hybrids have been popular for a long time. However, in Africa where downy mildew is much more of a problem, single-cross hybrids have either been found to be susceptible or when initially resistant found to rapidly become susceptible – once a new pathotype arises that can infect one plant then all of the plants of a single-cross hybrid are susceptible. However, the variation among the plants in a topcross hybrid overcomes this problem, particularly if variation for site-specific resistances have been incorporated into the population using data from multilocational disease screening. The topcross hybrids also displayed significant heterosis for downy mildew resistance as the hybrids had superior resistance to the parents. Apart from giving more durable resistance to downy mildew, variability can be disadvantageous in hybrids as it makes them less uniform phenotypically. Hence, strong selection pressure was applied for phenotypic traits that affect the appearance of uniformity in the hybrid such as plant height, panicle length and maturity. As a result, the topcross hybrids were more uniform than open-pollinated varieties or landraces but, of course, not as uniform as can be achieved in single-cross hybrids.
Having partially inbred parents is advantageous as they yield more than inbred lines and have more durable disease resistance because of their heterogeneity. Also, having non-inbred parents makes it possible to continuously improve important traits such as downy mildew resistance and synchrony of flowering between the parents. Combining ability can also be improved by reciprocal recurrent selection.
Conclusions
We have shown the superiority of topcross hybrids to the two current options available to African pearl millet farmers, namely improved varieties or landraces. However, to translate this advantage into higher yields in farmers’ fields seed needs to be commercially produced, but there are challenges in the large-scale production of topcross hybrid seed. Breeders have a set of challenges that can be overcome as long as the breeding programme has adequate resources. The non-inbred nature of the parents requires more breeding work than when inbred lines are used. To maintain sterility in the seed parent and maintain fertility restoration in the male, sib-mating (individual plant-by-plant crosses) is required. In addition, efforts had to be made to improve flowering synchrony between the parents because for efficient seed production they must flower either at the same time or the male parent has to commence flowering 1 or 2 days earlier. This was successfully done by selecting for earliness in the partially inbred restorer populations Toroniou and Maiwa from 2013 to 2018.
Unlike with the inbred lines used to make single-cross hybrids, it is simple to introgress new ‘donor’ material into the male parent when there is insufficient genetic variability to make progress for a particular trait such as downy mildew resistance or improved earliness. For effective introgression, individual crosses are made between the ‘donor’ line and a plant from the restorer population. This cross produces a full-sib progeny which is tested for good expression of the donor trait, a good match to the restorer population’s phenotype and a yield as high or greater than the restorer population. If these criteria are met, remnant full-sib seed is added to the restorer population.
The second set of challenges is to produce the seed commercially. Seed producers in Mali are, of course, unfamiliar with pearl millet hybrid seed production so, at least initially, training and subsidies are required for the producers to accept the risk of producing this novel cultivar type. The challenges include the need for isolated production plots in a region where pearl millet cultivation is widespread. Seed production could be done in the off season under irrigation, but this has not proved practicable: an irrigated seed production plot is the only pearl millet crop for kilometres so it will inevitably attract heavy predation by birds that cannot be economically prevented, and off-season differences in climate compared with the main season can decrease production and change flowering synchrony.
Seed production has been undertaken from 2014 to 2021 at research stations at Cinzana and N’Tarla and with agricultural cooperatives at Doumba and Banamba. Staff of the cooperatives were given training on seed production techniques including the roguing of off-type plants and any fertile plants in the male-sterile population. Two more cooperatives are interested to try production in 2022. The scale of production is around 0.5 ha in the cooperatives and 1 ha in the research stations. The two greatest problems have proved to be finding sufficient isolation distance in the case of the cooperatives and, for all, the problem of marketing. Farmers are not used to purchasing pearl millet seed as they normally use barter and the price of the seed is high. Farmers can buy seed at around €4 kg−1 so to sow a hectare they need to spend €20. If they gain 1 t ha−1 in yield, then at current grain prices (which vary according to whether it is sold immediately after harvest or in March when prices are highest) the farmer’s return on each euro spent on seed is returned 10–24 fold. The high price of seed is because the yields, excluding that of the pollinator, were low amounting to only about 350 kg ha−1 for the cooperatives and about 450 kg ha−1 on the research station. Apart from price considerations, marketing was difficult as farmers were generally unaware of these hybrids and the advantages they provide.
Outstanding seed parents such as Civarex 06_05 do produce topcross hybrids that are greatly superior for yield and downy mildew resistance over open-pollinated varieties and local landraces. If seed companies take up the commercial opportunities provided by these hybrids, they could usher in a new era of higher yielding pearl millet in Africa.
Supplementary material
For supplementary material for this article, please visit https://doi.org/10.1017/S0014479722000175
Acknowledgements
The authors thank Dr Bettina IG Haussmann, Principal Investigator of project ‘Tackling abiotic production constraints in pearl millet and sorghum-based agricultural systems of the West African Sahel’ and the German foreign assistance programme that financed the project, Vincent Vadez for welcoming the senior author in his laboratory at ICRISAT, Alliance for a Green Revolution for Africa (AGRA) for its support in the development of topcross hybrids in the projects ‘Participatory Development of Millet Parental lines and Hybrids production Adapted to the Sahelian and Sudanian Zone of Mali’ and ‘Testing and creation of top cross hybrids adapted to Sahelian and Sudanese zones of Mali’ and particularly Dr Joseph Devries and Dr Aboubacar Toure for having the belief that IER could develop the first pearl millet topcross hybrids for Africa and for their technical support.
Conflict of Interests
The authors declare none.









 Open access
Open access