


INTRODUCTION
West Nile virus (WNV) is a mosquito-borne virus of the genus Flavivirus, family Flaviviridae. The geographical distribution of this virus is very large as its circulation has been reported in most of the continents (except Antarctica). Birds are the reservoir of the virus and its natural transmission cycle involves mosquitoes. Humans and horses show a short-term viraemia at low levels, and are thus considered dead-end hosts. WNV causes West Nile fever (WNF) in both species, characterized by a febrile illness, which may progress to a meningoencephalitic syndrome in a small proportion of cases. In Morocco, WNF was first reported in 1996, with 94 equine cases of which 42 died [Reference Murgue1]. WNF was again reported in equines in 2003 (nine cases of which five died) [Reference Schuffenecker2] and in 2010 (25 cases of which eight died) [Reference Benjelloun, El Harrak and Belkadi3] (Fig. 1). A serosurvey conducted in 2008 showed that WNV did circulate in Moroccan resident bird populations [Reference Figuerola4]. Recently, in the absence of clinical cases, significant seroprevalence rates for WNV (5–19%, seroneutralization assay) have been reported in humans in several Moroccan provinces [Reference El Rhaffouli5, Reference El Rhaffouli6]. Usutu virus (USUV) probably circulates in Morocco, as serological evidence of infection has been reported in wild birds, although at a lower level than for WNV [Reference Figuerola4]. Other flaviviruses circulate in Africa but none has ever been reported in Morocco. African areas with risk of yellow fever virus transmission are restricted to sub-Saharan countries [Reference Jentes7], and this is also the case for laboratory-confirmed cases of dengue virus infection [Reference Amarasinghe8]. Similarly, Koutango virus has only been isolated south of the Sahara, in Senegal and Somalia, and Bagaza virus has been isolated in Western and Central Africa. Wesselbron virus infection appears endemic in Southern Africa [Reference Mathengtheng and Burt9], and Zika virus has never been isolated in Maghreb (http://www.cdc.gov/zika/geo/index.html). The circulation of WNV is seasonal in Maghreb, with outbreaks in summer and autumn. It is also probably the case for USUV but the epidemiology of this virus in Africa is less well known [Reference Nikolay10].

Fig. 1. Map of the study area showing the location of the military sites where animals were sampled (•) or where they had been stationed during their military career (◦) (dark grey shading: areas where clinical cases were reported in horses in 1996, 2003 and/or 2010, lines: province boundaries; A, Agadir; B, Ain el Aouda; C, Benguerir; D, Ben Slimane; E, Casablanca; F, Tadla; G, Kenitra; H, Khenifra; I, Laâyoune; J, Marrakhech; K, Meknes; L, Oujda; M, Rabat; N, Safi; O, Salé; P, Sidi Slimane; Q, Temara).
Although some field studies have reported clinical cases of WNF in canids [Reference Buckweitz11–Reference Read, Rodriguez and Summers13], experimental studies failed to reproduce the disease in the dog model [Reference Austgen14, Reference Blackburn15] and the short-term and low-level viraemia [Reference Austgen14] makes it unlikely that dogs play a role in the WNV epidemiological cycle. Several studies have reported significant WNV seroprevalence rates in dogs in Africa [Reference Blackburn15, Reference Davoust16], Europe [Reference Davoust16, Reference Busani17], Turkey [Reference Ozkul18], the United States [Reference Kile19–Reference Lillibridge23] and China [Reference Lan24]. Moreover, in the United States, antibodies were detected in a population of young dogs several weeks before the first reported human cases [Reference Resnick25]. This suggests that dogs may be useful as sentinels providing early warning of virus circulation.
The serological study reported here had two objectives: (i) to evaluate the extent of past WNV circulation in Morocco, using horses as sentinels, and (ii) to analyse the relevance of dogs as sentinels of past WNV circulation, in comparison with horses. The study focused on military working animals (horses and dogs), for which service record books allowed a precise reconstruction of the military career of each animal. Serum samples were tested for antibodies against WNV and USUV.
MATERIALS AND METHODS
Animal samples
Moroccan military horses and/or dogs were stationed in 18 sites, among which 10 were selected for the study (Table 1, Fig. 1), based on the number of animals stationed (n > 20), and for practical reasons (geographical distance and accessibility). In order to compare dogs and horses as sentinels of past WNV circulation, five sites were selected in areas affected by previous epizootics of WNV (Kenitra, Khenifra, Benslimane, Sidi Slimane, Casablanca), and which housed both working horses and dogs. The five other sites were selected in areas where no WNF clinical case had ever been reported (Temara, Meknes, Marrakech, Agadir, Salé), in order to evaluate the extent of past WNV circulation in Morocco: in these sites, only equine samples were analysed. All the military working horses and dogs present at each of these ten sites were included in the study, except for Khenifra and Temara where the horse population was large (281 and 261 animals, respectively) and ~100 animals were randomly selected. The animals were blood-sampled in February 2012. Data collected for each animal of the study included the birth date, species and sex (stallion or male, vs. gelding or female). Service record books were screened for reconstructing the military career of the animal with, for each of its successive postings, the site and the arrival and departure dates. The dates of WNV vaccinations of horses were also collected. The whole study was designed and conducted in collaboration with the Moroccan Army. Data collection, field samples and protocols were performed by a veterinarian from the Royal Armed Forces, in accordance with the Moroccan regulations regarding animal welfare.
Table 1. Flavivirus seroprevalence, estimated by WNV cELISA, in military horses and dogs, Morocco, 2012

CI, Confidence interval.
* Provinces in which West Nile fever cases have been reported in horses in 1996, 2003 and/or 2010.
Serological tests
Competitive enzyme-linked immunosorbent assay (cELISA)
Serological screening of horse and dog sera was performed using a cELISA test (ID Screen® West Nile Competition ELISA kit, ID Vet, France). Assays were performed and interpreted according to the manufacturer's instructions. The threshold value for considering a serum as positive was %OD sample/negative control (S/N) <40% as recommended by the manufacturer. This test is often used for identifying anti-WNV immunoglobulins, but cross-reactions with other flaviviruses have been described [Reference Beck26–Reference Ziegler29]. Therefore a positive result was interpreted as indicating the presence of immunoglobulins directed against a flavivirus (either WNV or other flaviviruses).
WNV non-structural protein 1(NS1)-ELISA (DIVA test)
Equine serum samples positive by WNV cELISA were screened using an in-house epitope-blocking ELISA as developed by Blitvich et al. [Reference Blitvich30]. MaxiSorp 96-well ELISA plates (Nunc, Dutscher-Scientific, UK) were coated with WNV-inactivated antigens (kindly provided by Dr P. Desprès, Pasteur Institute) diluted 1:1000 in coating buffer (50 mm NaHCO3, 50 mm Na2CO3, pH 9·6) (VWR BDH-Prolabo, UK). A total of 100 µl diluted antigen was distributed in even columns, while 100 µl coating buffer alone was distributed in odd columns. Plates were incubated overnight at 4 °C as well as for 1 h at 37 °C on a plate shaker. Wells were washed four times with 0·1% Tween 20 in PBS (pH 7·4) (washing solution, Gibco, Life Technologies, USA). A total of 200 µl blocking buffer containing 0·2% bovine serum albumin (Sigma, USA) in washing buffer were added. Plates were incubated for 45 min at 37 °C under agitation and washed three times. Serum samples as well as positive and negative controls (NC), diluted 1:10 in blocking buffer, were added to the wells and incubated for 2 h at 37 °C under agitation. After four washing steps, 50 µl mouse anti-WNV NS1 monoclonal antibody (MAB8152, 3·1112 G clone, Merck Millipore, USA) diluted 1:4000 in blocking buffer was added to the wells, and incubated for 1 h at 37 °C under agitation, and for an additional 1 h at 4 °C. After four washing steps, F(ab′)2 rabbit anti-mouse IgG antibodies conjugated to horseradish peroxidase (Star 13B, Serotec, USA) were diluted 500-fold in blocking buffer with 10% of heat-inactivated horse serum (Gibco) and incubated for 1 h at 37 °C under agitation. After four washing steps, 200 µl TMB (Sigma) diluted 1:10 in phosphate citrate buffer was added to the wells. After 30 min incubation at 37 °C on a plate shaker, the reaction was stopped by adding 100 µl H2SO42N (Normapur Prolabo) in each well. The optical density (OD) was read at 450 nm with an ELISA microwell plate reader (Thermo Scientific Multiskan EX, USA). Blocking percentages were determined for each sample as
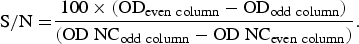 $${\rm S}/{\rm N =} \displaystyle{{100 \times ({\rm OD}_{\rm {even \ column}} - {\rm OD}_{\rm {odd \ column}} )} \over {({\rm OD}\; {\rm NC}_{\rm {odd \ column}} - {\rm OD}\;{\rm NC}_{\rm {even \ column}} )}}.$$
$${\rm S}/{\rm N =} \displaystyle{{100 \times ({\rm OD}_{\rm {even \ column}} - {\rm OD}_{\rm {odd \ column}} )} \over {({\rm OD}\; {\rm NC}_{\rm {odd \ column}} - {\rm OD}\;{\rm NC}_{\rm {even \ column}} )}}.$$
A sample was considered positive if S/N <30%.
Virus-specific microneutralization tests (MNTs)
To identify the circulating flaviviruses, WNV and USUV MNTs were performed on cELISA-positive sera: for each of the ten military sites and for both studied species, all of the cELISA-positive sera were tested, with a maximum of 20 randomly selected sera per site and per species. WNV and USUV neutralizing antibody titres were determined by MNT on Vero cells in 96-well cell culture plates, following the protocol described in Beck et al. [Reference Beck31], with WNV strain IS-98-ST1 and USUV strain SAAR-1776 (South Africa). The MNT results were interpreted according to the following rules: (i) if the WNV antibody titre was strictly greater than the USUV antibody titre, the serum was classified WNV positive, (ii) if the WNV antibody titre was strictly lower than the USUV antibody titre, the serum was classified USUV positive, and (iii) if both antibody titres were identical (and >0), the serum was classified positive for an undetermined flavivirus.
Statistical analyses
Site- and species-specific seroprevalence rates were computed, as well as their 95% confidence intervals (CIs). Horse and dog seroprevalence rates were compared in order to evaluate the relevance of dogs as sentinel species for the circulation of flaviviruses. However, because exposure to flaviviruses was likely to vary across military sites, the individual exposure of a given animal varied between animals, according to their successive postings during their military careers. Species-specific prevalence rates could thus not be directly compared and a multivariate logistic model was used to control this confounding factor when analysing the relationship between seropositivity and species. The dependent variable was the cELISA result (positive or negative). Explanatory variables were the species (horse or dog) and its interaction with the genus (as housing conditions differed according to the genus in horses but not in dogs), the vaccination status, the potential exposure to WNV circulation during reported epizootics (three binary variables indicating whether the animal was born before the reported epizootics of 1996, 2003 and 2010), and the time spent by the animal in the various military sites of Morocco (one quantitative variable for each of the various military sites, plus a supplementary variable for the time spent by the animal in Morocco before its integration into the Moroccan Army) (Fig. 1). The latter variables both represented variations across sites of the level of viral circulation, and differences of animal exposure to mosquito bites between sites (due to differences of housing conditions and/or to geographical variations of mosquito abundance).
RESULTS
Three hundred and forty-nine military horses and 231 dogs were included in the study.
Eighty-three percent (289/349) of the tested horses originated from Moroccan studs, the remainder from European studs. After their integration in the army, horses were stationed at up to three military sites, but the majority (88%, 307 animals) were stationed at a single location. The age of horses varied between 18 months and 32 years, with an average of 14·5 years. Most animals had been vaccinated (88%, 308 animals) against WNV with a canarypox-vectored vaccine expressing the premembrane (prM) and envelope (E) genes of WNV (Recombitek® rWNV vaccine, Merial, USA). A single vaccine injection was administered for most (n = 212) animals and the others received one boost 3 months later. The single or last vaccine injection was performed, on average, 16 months before the blood samples were taken (range 13–17 months).
Fifty-nine percent of dogs had been born in a military site (137/231), others originated from European breeders. Dogs had been successively stationed at 1–5 sites, but most had been stationed at one (67%, 155 animals) or two (23%, 54 animals) sites. The average age of dogs was 4·5 years (range 6 months to 13 years).
The overall cELISA seroprevalence rate was 60% in horses (95% CI 55–65) and 62% in dogs (95% CI 55–68) (Table 1). Among sites with ⩾20 tested sera (five sites for horses and two sites for dogs), the seroprevalence rate ranged between 29% (Khenifra) and 94% (Temara) in horses, and between 54% (Benslimane) and 96% (Kenitra) in dogs. Similar prevalence rates were obtained when restricting the dataset to animals which had been born in a military site (or which had been imported there from Europe), and which had remained there throughout their military career. For example, 95% of horses that had always lived at the Temara site were cELISA positive (42/44 animals), compared to an overall seroprevalence of 94%.
Antibodies directed against WNV NS1 protein were detected in 79% of tested horse sera (72/91, 95% CI 69–87) suggesting that most cELISA-positive results were due to WNV infection. No significant difference in the proportion of NS1-ELISA-positive sera was observed between vaccinated and non-vaccinated animals (Fisher's test, P > 0·05).
In horses, 74% (78/106) of cELISA-positive sera were found to be WNF positive by seroneutralization, 4% (4/106) USUV positive, and 2% (2/106) were classified positive for an undetermined flavivirus. Twenty-one percent (22/106) were negative for both viruses (Table 2). For dogs, 80% (48/60) of cELISA-positive sera were positive for WNV by seroneutralization, 12% (7/60) for USUV, and 8% (5/60) were classified positive for an undetermined flavivirus. None of the cELISA-positive sera were negative for both viruses (Table 2).
Table 2. Comparison of seroneutralization titres against WNV and USUV in military working horses and dogs, Morocco, 2012

WNV, West Nile virus; USUV, Usutu virus
a Number of sera classified WNV positive (WNV titre > USUV titre, numbers in plain font).
b Number of sera classified USUV positive (WNV titre < USUV titre, numbers in italics).
c Number of sera classified positive for an undetermined flavivirus (WNV titre = USUV titre and WNV titre > 0, numbers in bold). Underlined: sera classified negative for WNV and USUV.
The multivariate model did not show any significant effect of the species on cELISA seropositivity risk (Table 3). Conversely, a birth date before September 2010 (and having thus been potentially exposed to the 2010 WNV epizootic) was a seropositivity risk factor [odds ratio (OR) 7·4]. Being a stallion induced a protective effect (OR 0·3), probably because stallions are kept in housing conditions where they are less likely to be exposed to mosquito bites. Finally, having been stationed at Kenitra and Temara were seropositivity risk factors (OR 1·4 and 1·3, respectively, for a posting of 1 year).
Table 3. Multivariate model of flavivirus seroprevalence in military horses and dogs, Morocco, 2012

OR, Odds ratio; CI, confidence interval; n.s., not significant.
* Location before integration in the Moroccan Army.
DISCUSSION
The serological results reported here indicate a large geographical distribution of flaviviruses in Morocco. A seroprevalence rate of 60% has been observed in military working horses, which lies between values reported in sub-Saharan African countries such as Senegal (85% [Reference Chevalier32]) where WNV is considered endemic, and European countries such as Spain (7% in Andalusia [Reference Garcia-Bocanegra33] and 8% in Doñana [Reference Jimenez-Clavero34]) or France (8·5% in the Camargue area [Reference Durand35] and 34% in the Var department [Reference Durand36]).
High seroprevalence rates were observed in provinces where clinical cases had been previously reported in horses, such as Kenitra (82%). This result may be compared with the seroprevalence rate of 19% observed in sera collected 12 months earlier from humans living in the same area [Reference El Rhaffouli5]. The geographical distribution of flavivirus was, however, not limited to areas where clinical cases had been previously reported. Indeed, the highest seroprevalence rate observed in horses was obtained for the Temara site, located in an area from where no WNF case has ever been reported (although the province of Temara is close to areas affected in 1996, 2003 or 2010). For this site, the horse seroprevalence rate was 94%, and 95% when considering only animals that had always lived there. Similarly, whereas no WNF case has ever been reported in Meknes province, the horse seroprevalence rate was 34%, compared to the seroprevalence rate of 5% obtained in humans in the same area 12 months earlier [Reference El Rhaffouli5].
Most of the tested horses had been vaccinated against WNV and some cELISA-positive results may be attributed to vaccine-induced antibodies. However, vaccine-induced antibodies have a shorter half-life than WNV-induced antibodies, as they are usually detected up to 300 days post-injection [Reference Khatibzadeh37]. All of the tested horses had received the last vaccine injection >400 days before blood samples were taken, and most had received a single vaccine injection, in 2010. The majority of the cELISA-positive results are thus probably attributable to natural antibodies, which is confirmed by the 79% of cELISA-positive sera that tested positive using a WNV-DIVA test (detection of antibodies directed against WNV NS1 protein, which was not included in the WNV vaccine used). Discordant cELISA-positive and NS1-ELISA negative results could be due to prior and confounding vaccination, but also to other factors. Weakly WNV-positive sera may give NS1-ELISA-negative results: four NS1-ELISA-negative sera had a cELISA S/N value between 30% and 40%, and seven had a cELISA S/N value between 20% and 30% (data not shown). Infection with a flavivirus different from WNV may also explain discordant results, as the NS1-ELISA is far more specific for WNV antibodies than WNV cELISA: indeed, two NS1-ELISA-negative horses were found to have antibodies to USUV (data not shown). Seroneutralization test results indicate that most of the cELISA-positive results observed in horses were due to WNV (74%), few sera being classified USUV positive (4%). Twenty-two samples were MNT negative and corresponded to samples with low anti-flavivirus response (low positives by cELISA), confirming the previously lower sensitivity of MNT [Reference Weingartl38].
The overall cELISA seroprevalence rate obtained in dogs (62%) was remarkably close to that obtained in horses (60%), and site-specific seroprevalence rates observed in dogs were close to those observed in horses. As for horses, seroneutralization results indicate a predominant exposure of dogs to WNV (80%); however, a significant proportion of sera were classified USUV positive (12%). The multivariate model did not show any significant effect of the species on cELISA status, despite several expected effects on the seropositivity risk. For example, two variables describing an exposure to previous epizootics were seropositivity risk factors: being born before the last WNF epizootic of 2010, and having been stationed at Kenitra (i.e. in a province where WNF cases had been reported in 1996, 2003 and 2010). Similarly, a variable linked to housing conditions with a lower exposure to mosquito bites (i.e. being a stallion) was a protective factor. The fact that military working dogs moved on average more often than horses between military sites complicated the comparison of seroprevalence between the species, but the multivariate model allowed this confounding factor to be controlled for. Therefore our results suggest that, under similar exposure conditions to mosquito bites, the WNV seropositivity risk in dogs and horses is similar. Considering the shorter lifespan of dogs, this may indicate a stronger exposure of dogs than horses to mosquito bites, which is in line with results obtained in Europe (Barcelona, Spain), where bloodmeal analysis of Culex pipiens has shown that this vector of WNV had often acquired blood from dogs [Reference Munnoz39], and in the United States where similar results have been obtained for Culex quinquefasciatus and Aedes albopictus [Reference Dennett40, Reference Molaei41].
In conclusion, our study reveals a large geographical distribution of WNV circulation in Morocco, and the first evidence of USUV circulation in Morocco in horses and dogs. The similar seroprevalence rates observed in military working dogs and horses suggest that dogs may be a relevant choice for serological surveillance of WNV or USUV circulation, especially in areas where equine vaccination precludes passive surveillance (based on the detection of WNF cases) in horses. However, whether a dog-based sentinel surveillance system would enable early detection of WNV circulation remains to be investigated.
ACKNOWLEDGEMENTS
The authors thank Philippe Desprès (Pasteur Institute, Paris) for kindly providing the viral strains and antigens used in the study.
DECLARATION OF INTEREST
None.




