
6 results
Stibiogoldfieldite, Cu12(Sb2Te2)S13, a new tetrahedrite-group mineral
-
- Journal:
- Mineralogical Magazine / Volume 86 / Issue 1 / February 2022
- Published online by Cambridge University Press:
- 07 January 2022, pp. 168-175
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
Stibiogoldfieldite, Cu12(Sb2Te2)S13, was approved as a new mineral species from the Mohawk mine, Goldfield mining district, Esmeralda County, Nevada, USA. It occurs as metallic anhedral grains, dark grey in colour. It is associated with quartz, pyrite and an Ag–Bi–(S,Se) phase (holotype material) and with quartz, pyrite, calaverite, bismuthinite, bohdanowiczite, and the Ag–Bi–(S,Se) phase (cotype material). In reflected light, stibiogoldfieldite is isotropic, grey in colour, with indistinct brownish shade. Reflectance data in air [R (%)] are: 31.1 at 470 nm, 30.9 at 546 nm, 30.8 at 589 nm and 31.0 at 650 nm. Electron microprobe analysis for holotype material gave (in wt.% – average of 60 spot analyses): Cu 45.03(60), Ag 0.26(7), Fe 0.02(3), Zn 0.13(15), Sn 0.02(4), Pb 0.05(6), Sb 8.02(62), As 2.80(65), Bi 2.77(87), Te 15.15(1.24), S 24.50(32), Se 0.52(11), total 99.27(69). On the basis of (As + Sb + Te + Bi) = 4 atoms per formula unit (apfu), the empirical formula of stibiogoldfieldite is (Cu12.05Ag0.04Zn0.03Fe0.01)Σ12.13(Sb1.12As0.63Bi0.23Te2.02)Σ4.00(S12.99Se0.11)Σ13.10. Chemical data on an additional sample from the same locality (cotype material) gave the following results (in wt.% – average of 181 spot analyses): Cu 43.84(63), Ag 0.21(7), Sb 5.92(78), As 2.63(45), Te 20.07(1.19), S 25.13(53), Se 0.97(35), total 99.47(66). On the basis of (As + Sb + Te + Bi) = 4 apfu, the empirical formula of cotype material is (Cu11.30Ag0.03)Σ11.33(Sb0.80As0.57Bi0.06Te2.57)Σ4.00(S12.83Se0.20)Σ13.03. Stibiogoldfieldite is cubic, I$\overline 4$
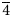 3m, with unit-cell parameters a = 10.3466(17) Å, V = 1107.6(5) Å3 and Z = 2 (holotype). Unit-cell parameters for the cotype sample are a = 10.3035(2) Å and V = 1093.83(7) Å3. The crystal structure of holotype stibiogoldfieldite was refined by single-crystal X-ray diffraction data to a final R1 = 0.032 on the basis of 285 reflections with Fo > 4σ(Fo) and 20 refined parameters. Stibiogoldfieldite is isotypic with other members of the tetrahedrite group.
3m, with unit-cell parameters a = 10.3466(17) Å, V = 1107.6(5) Å3 and Z = 2 (holotype). Unit-cell parameters for the cotype sample are a = 10.3035(2) Å and V = 1093.83(7) Å3. The crystal structure of holotype stibiogoldfieldite was refined by single-crystal X-ray diffraction data to a final R1 = 0.032 on the basis of 285 reflections with Fo > 4σ(Fo) and 20 refined parameters. Stibiogoldfieldite is isotypic with other members of the tetrahedrite group.
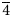
Ferro-ferri-hornblende from the Traversella mine (Ivrea, Italy): occurrence, mineral description and crystal-chemistry
-
- Journal:
- Mineralogical Magazine / Volume 80 / Issue 7 / December 2016
- Published online by Cambridge University Press:
- 02 January 2018, pp. 1233-1242
-
- Article
- Export citation
-
Ferro-ferri-hornblende is a new member of the amphibole supergroup (IMA-CNMNC 2015-054). It has been found in a rock specimen from the historical collection of Leandro De Magistris, which was collected at the Traversella mine (Val Chiusella, Ivrea, Piemonte, Italy). The specimen was catalogued as ‘speziaite', and contains a wide range of amphibole compositions from tremolite/actinolite to magnesio-hastingsite. The end-member formula of ferro-ferri-hornblende is A□BCa2c(Fe+Fe3+)T(Si7Al) O22W(OH)2 , which requires SiO2 43.41, Al2O3 5.26, FeO 29.66, Fe2O3 8.24 CaO 11.57, H2O 1.86, total 100.00 wt.%. The empirical formula derived from electron microprobe analysis and single-crystal structure refinement for the holotype crystal is A(Na0.10K0.13) Σ=0.23B(Ca 1.93Na0.07)Σ=2.00C(Mg1.16Fe2+3.21Mn0.O6Fe3+0.45 Al0.12Ti 0.01)Σ=5.01T(Si7.26Al0. 74)Σ=8.00 O22W(OH1.89F0.01C10.10)Σ=2.00- Ferro-ferri-hornblende is biaxial (-), with α = 1.697(2), P = 1 .722(5), γ = 1.726(5) and 2V (meas.) = 35.7(1.4)°, 2V (calc.) = 43.1°. The unit-cell parameters are a = 9.9307(5), b = 18.2232(10), c = 5.3190(3) Å, β = 104.857(1)°, V= 930.40 (9) Å3, Z= 2, space group C2/m. The a:b:c ratio is 0.545:1:0.292. The strongest eight reflections in the powder X-ray pattern [d values (in Å), I, (hkl)] are: 8.493, 100, (110); 2.728, 69, (151); 3.151, 47, (310); 2.555, 37, (
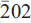 ); 2.615, 32, (061); 2.359, 28, (
); 2.615, 32, (061); 2.359, 28, (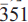 ); 3.406, 26, (131); 2.180, 25, (261). Type material is deposited in the collections of the Museo di Mineralogia, Dipartimento di Scienze della Terra e dell'Ambiente, Università di Pavia, under the catalogue number 2015-01. Sample M/U15285 from the historical collection of Luigi Colomba, presently at the Museo Regionale di Scienze Naturali di Torino, was also checked, and the presence of ferro-ferri-hornblende was confirmed.
); 3.406, 26, (131); 2.180, 25, (261). Type material is deposited in the collections of the Museo di Mineralogia, Dipartimento di Scienze della Terra e dell'Ambiente, Università di Pavia, under the catalogue number 2015-01. Sample M/U15285 from the historical collection of Luigi Colomba, presently at the Museo Regionale di Scienze Naturali di Torino, was also checked, and the presence of ferro-ferri-hornblende was confirmed.
 ); 2.615, 32, (061); 2.359, 28, (
); 2.615, 32, (061); 2.359, 28, (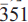 ); 3.406, 26, (131); 2.180, 25, (261). Type material is deposited in the collections of the Museo di Mineralogia, Dipartimento di Scienze della Terra e dell'Ambiente, Università di Pavia, under the catalogue number 2015-01. Sample M/U15285 from the historical collection of Luigi Colomba, presently at the Museo Regionale di Scienze Naturali di Torino, was also checked, and the presence of ferro-ferri-hornblende was confirmed.
); 3.406, 26, (131); 2.180, 25, (261). Type material is deposited in the collections of the Museo di Mineralogia, Dipartimento di Scienze della Terra e dell'Ambiente, Università di Pavia, under the catalogue number 2015-01. Sample M/U15285 from the historical collection of Luigi Colomba, presently at the Museo Regionale di Scienze Naturali di Torino, was also checked, and the presence of ferro-ferri-hornblende was confirmed.Morning versus afternoon cutting time of Berseem clover (Trifolium alexandrinum L.) affects feed intake, milk yield and composition in Girgentana goats
-
- Journal:
- Journal of Dairy Research / Volume 78 / Issue 4 / November 2011
- Published online by Cambridge University Press:
- 23 September 2011, pp. 500-504
- Print publication:
- November 2011
-
- Article
- Export citation
Effect of CSN1S1 genotype and its interaction with diet energy level on milk production and quality in Girgentana goats fed ad libitum
-
- Journal:
- Journal of Dairy Research / Volume 77 / Issue 2 / May 2010
- Published online by Cambridge University Press:
- 12 April 2010, pp. 245-251
- Print publication:
- May 2010
-
- Article
- Export citation
Diet selection and milk production and composition in Girgentana goats with different αs1-casein genotype
-
- Journal:
- Journal of Dairy Research / Volume 76 / Issue 2 / May 2009
- Published online by Cambridge University Press:
- 13 March 2009, pp. 202-209
- Print publication:
- May 2009
-
- Article
- Export citation
Milk quality as affected by grazing time of day in Mediterranean goats
-
- Journal:
- Journal of Dairy Research / Volume 75 / Issue 1 / February 2008
- Published online by Cambridge University Press:
- 29 January 2008, pp. 48-54
- Print publication:
- February 2008
-
- Article
- Export citation
