
14 results
Arazyme secreted by Serratia proteamaculans: current understanding in animal husbandry – CORRIGENDUM
-
- Journal:
- The Journal of Agricultural Science / Volume 161 / Issue 5 / October 2023
- Published online by Cambridge University Press:
- 31 January 2024, p. 703
-
- Article
-
- You have access
- HTML
- Export citation
Arazyme secreted by Serratia proteamaculans: current understanding in animal husbandry
-
- Journal:
- The Journal of Agricultural Science / Volume 161 / Issue 5 / October 2023
- Published online by Cambridge University Press:
- 07 August 2023, pp. 697-702
-
- Article
-
- You have access
- HTML
- Export citation
2.1
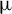 $\unicode{x3bc}$m, high-energy dissipative soliton resonance from a holmium-doped fiber laser system
$\unicode{x3bc}$m, high-energy dissipative soliton resonance from a holmium-doped fiber laser system
-
- Journal:
- High Power Laser Science and Engineering / Volume 11 / 2023
- Published online by Cambridge University Press:
- 17 January 2023, e12
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
NONPARAMETRIC ESTIMATION OF REGRESSION FUNCTIONS WITH DISCRETE REGRESSORS
-
- Journal:
- Econometric Theory / Volume 25 / Issue 1 / February 2009
- Published online by Cambridge University Press:
- 01 February 2009, pp. 1-42
-
- Article
- Export citation
Impact of epistasis and QTL×environment interaction on the accumulation of seed mass of soybean (Glycine max L. Merr.)
-
- Journal:
- Genetics Research / Volume 90 / Issue 6 / December 2008
- Published online by Cambridge University Press:
- 06 January 2009, pp. 481-491
-
- Article
-
- You have access
- HTML
- Export citation
15 - Medical management of captive adult and geriatric giant pandas
-
-
- Book:
- Giant Pandas
- Published online:
- 09 August 2009
- Print publication:
- 27 July 2006, pp 353-376
-
- Chapter
- Export citation
20 - Role and efficiency of artificial insemination and genome resource banking
-
-
- Book:
- Giant Pandas
- Published online:
- 09 August 2009
- Print publication:
- 27 July 2006, pp 469-494
-
- Chapter
- Export citation
13 - The neonatal giant panda: hand-rearing and medical management
-
-
- Book:
- Giant Pandas
- Published online:
- 09 August 2009
- Print publication:
- 27 July 2006, pp 315-333
-
- Chapter
- Export citation
9 - The value and significance of vaginal cytology
-
-
- Book:
- Giant Pandas
- Published online:
- 09 August 2009
- Print publication:
- 27 July 2006, pp 231-244
-
- Chapter
- Export citation
7 - Male reproductive biology in giant pandas in breeding programmes in China
-
-
- Book:
- Giant Pandas
- Published online:
- 09 August 2009
- Print publication:
- 27 July 2006, pp 159-197
-
- Chapter
- Export citation
4 - Significant medical issues and biological reference values for giant pandas from the Biomedical Survey
-
-
- Book:
- Giant Pandas
- Published online:
- 09 August 2009
- Print publication:
- 27 July 2006, pp 59-86
-
- Chapter
- Export citation
EQUI-ATTRACTION AND THE CONTINUOUS DEPENDENCE OF ATTRACTORS ON PARAMETERS
-
- Journal:
- Glasgow Mathematical Journal / Volume 46 / Issue 1 / January 2004
- Published online by Cambridge University Press:
- 15 January 2004, pp. 131-141
- Print publication:
- January 2004
-
- Article
-
- You have access
- Export citation
Dichotomies of solutions for a class of second-order nonlinear differential equations
-
- Journal:
- Glasgow Mathematical Journal / Volume 44 / Issue 2 / May 2002
- Published online by Cambridge University Press:
- 25 July 2002, pp. 339-348
- Print publication:
- May 2002
-
- Article
-
- You have access
- Export citation
Analysis of lectin receptors in normal nasal mucosa, nasal polyp, inverted papilloma and papillary adenocarcinoma
-
- Journal:
- The Journal of Laryngology & Otology / Volume 107 / Issue 7 / July 1993
- Published online by Cambridge University Press:
- 29 June 2007, pp. 600-602
- Print publication:
- July 1993
-
- Article
- Export citation
