



Introduction
Cretaceous lacustrine oil shales are widely distributed in rift basins near Mongolia's eastern and southeastern borders with China, forming a well-known petroleum system of eastern Asia (e.g., Yang et al., Reference Yang, Li and Gao1985; Graham et al., Reference Graham, Hendrix, Johnson, Badamgarav, Badarch, Amory, Porter, Barsbold, Webb and Hacker2001; Johnson et al., Reference Johnson, Greene, Zinniker, Moldowan, Hendrix and Carroll2003; Prost, Reference Prost2004; Hasegawa et al., Reference Hasegawa2018). Due to the high organic carbon oil shale, many petroleum explorations have been conducted. In particular, Johnson et al. (Reference Johnson, Constenius, Graham, Mackey, Menotti, Payton and Tully2015) stated that this petroleum system extends west into the Gobi–Altai region of Mongolia according to seismic data, cores, and sedimentological outcrop studies. However, the age of oil-shale-bearing formations in the Gobi–Altai region is debated because of a lack of well-defined regional stratigraphic nomenclature and age determination (e.g., Khand et al., Reference Khand, Badamgarav, Ariunchimeg, Barsbold, Okada and Mateer2000; Johnson et al., Reference Johnson, Constenius, Graham, Mackey, Menotti, Payton and Tully2015), which has hindered understanding of the formation of this big petroleum system in eastern Asia.
Ostracoda, a group of microcrustaceans with an excellent fossil record, are widely applied to biostratigraphy as well as paleoenvironmental and paleoclimate reconstruction (e.g., Horne, Reference Horne, Whittaker and Hart2009; Sames, Reference Sames and Sames2011a, Reference Sames and Samesb; Sames and Horne, Reference Sames and Horne2012; Wang et al., Reference Wang, Sha, Pan and Zhang2015; Choi et al., Reference Choi, Jia, Huh, Jung and Wang2021; Wang and Zhong, Reference Wang and Zhong2022). In particular, the well-known ostracod faunas from the so-called Purbeck–Wealden interval (latest Tithonian to earliest Aptian; see Sames and Horne, Reference Sames and Horne2012 for details) deposits were notably expedited by oil and gas exploration, including the marginal marine to nonmarine basins in Europe, Africa, and South America as well as the nonmarine rift basins of China (e.g., Erlian and Hailaer basins of China) (e.g., Gou et al., Reference Gou, Wang, Yang and Wang1986; Ye et al., Reference Ye, Huang, Liu, Zhang and Chen2003; Liu et al., Reference Liu, Huang and Ye2004). However, it should be noted that the taxonomic composition of Purbeck–Wealden type ostracod faunas from different basins in several continents can be quite different, as can the stratigraphic intervals represented. For example, the genus Theriosynoecum Branson, Reference Branson1936 is widely reported from the Purbeck–Wealden type ostracod faunas of Europe, Africa, South America, and North America, but it is rarely found in the Purbeck–Wealden type ostracod faunas of China (e.g., Hou et al., Reference Hou, Gou and Chen2002; Hou and Gou, Reference Hou and Gou2007; Sames, Reference Sames and Sames2011b).
In this study, we describe a moderately diverse Purbeck–Wealden type nonmarine ostracod fauna from the Shinekhudag Formation in the Gobi–Altai region of Mongolia. Our ostracod biostratigraphic data provide a new age constraint for the oil-shale-bearing Shinekhudag Formation in this area. In addition, our results provide insights into potential microfaunal interchange between distant nonmarine settings around eastern Asia. Moreover, paleoenvironmental interpretations based on ostracods provide new insights into the formation mechanism of source rocks in Gobi–Altai region.
Geological setting
The Gobi–Altai region of southern Mongolia comprises intramontane basins that contain a succession of Jurassic–Cretaceous sediments and lavas unconformably overlain by lesser volumes of Neogene–Recent alluvial fan, fluvial, and lacustrine deposits (Cunningham et al., Reference Cunninghan, Davies and Mclean2009) (Fig. 1). In this region, the Lower Cretaceous strata consist of the Tsagaantsav, Shinekhudag, Khukhteeg, and Zuunbayan formations in ascending order (Fig. 2) and their equivalents (Khand et al., Reference Khand, Badamgarav, Ariunchimeg, Barsbold, Okada and Mateer2000; Johnson et al., Reference Johnson, Constenius, Graham, Mackey, Menotti, Payton and Tully2015).

Figure 1. Locality map of the studied sections of the Lower Cretaceous Shinekhudag Formation, Gobi–Altai area, southwestern Mongolia (a: Tsagaan Tsuvarga; b: Oshih Hollow East).

Figure 2. Stratigraphic distribution and occurrence of ostracod species in the Shinekhudag Formation, southwestern Mongolia (stratigraphic chart modified from Johnson et al., Reference Johnson, Constenius, Graham, Mackey, Menotti, Payton and Tully2015).
The Shinekhudag Formation is also transcribed as “Shinhudag Formation” in several papers (see Graham et al., Reference Graham, Hendrix, Johnson, Badamgarav, Badarch, Amory, Porter, Barsbold, Webb and Hacker2001; Johnson et al., Reference Johnson, Constenius, Graham, Mackey, Menotti, Payton and Tully2015). It should be noted that Mongolian place and stratigraphic names have been spelled in many different ways. To minimize confusion, the translation standards suggested by Benton et al. (Reference Benton, Shishkin, Unwin, Kurochkin, Benton, Shishkin, Unwin and Kurochkin2000) are used in this study. The Shinekhudag Formation is well exposed in the type section of the Shine Khudag locality in the Shaazangiin Gobi area of the Darnogobi Province (Ando et al., Reference Ando, Hasegawa, Hasegawa, Ohta, Yamamoto, Hasebe and Ichinnorov2011; Hasegawa et al., Reference Hasegawa2018). In the Gobi–Altai region, Johnson et al. (Reference Johnson, Constenius, Graham, Mackey, Menotti, Payton and Tully2015) assigned the rock unit locally mapped and identified as Manlai Formation previously to the Shinekhudag Formation (= Shinhudag Formation) because of the age equivalence of the formations. The Shinekhudag Formation (~ 50–1,600 m thick in different regions; see Khand et al., Reference Khand, Badamgarav, Ariunchimeg, Barsbold, Okada and Mateer2000; Johnson et al., Reference Johnson, Constenius, Graham, Mackey, Menotti, Payton and Tully2015; Adiya et al., Reference Adiya, Johnson, Loewen, Ritterbush, Constenius and Dinter2017) is composed mainly of sandstones, silty mudstones, silty claystones, and oil shales; it overlies the Tsagaantsav Formation (also spelled “Tsagantsav Formation” and equals locally mapped “Undur Formation”) and underlies the Khukhteeg Formation (also spelled “Huhteeg Formation”) in the Gobi–Altai region (Khand et al., Reference Khand, Badamgarav, Ariunchimeg, Barsbold, Okada and Mateer2000; Johnson et al., Reference Johnson, Constenius, Graham, Mackey, Menotti, Payton and Tully2015; Hasegawa et al., Reference Hasegawa2018). To date, the age of the Shinekhudag Formation is still controversial. A Hauterivian–Barremian age for the Shinekhudag Formation had been proposed by several authors on the basis of ostracod fossils and 40Ar/39Ar ages (e.g., Khand et al., Reference Khand, Badamgarav, Ariunchimeg, Barsbold, Okada and Mateer2000; Johnson et al., Reference Johnson, Constenius, Graham, Mackey, Menotti, Payton and Tully2015). Hasegawa et al. (Reference Hasegawa2018) assigned an Aptian age to this formation on the basis of biostratigraphic correlations and U–Pb dating of zircons in intercalated tuff samples.
Materials and methods
The studied ostracod specimens were recovered from the dark silt claystones and mudstones of the Shinekhudag Formation in Tsagaan Tsuvarga (45°11′21″N, 99°02′55″E) and Oshih Hollow East (44°96′16″N, 103°13′94″E) areas of the petroleum blocks IV and V (Fig. 1; for Mongolia petroleum blocks, also see online map: https://www.petromatadgroup.com/mongolia/oil-in-mongolia/). The rock samples from the outcrops were collected by an American–Mongolian petroleum investigation group during July 2011. However, because only the sample numbers and height were given by the field geologists, the detailed information (e.g., lithology and sedimentary structures) of the sections is not represented in this study. Since the ostracods are associated with bivalves that shall remain intact, we could not use the “standard method” of chemical rock disintegration. Thus, we use fine needles and brushes to clean the surrounded sediments of each ostracod carapaces/valves and expose specimens. Selected ostracod specimens were coated with gold and photographed using a scanning electronic microscope TESCAN Maia3 at the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (Nanjing, China).
Morphologic terminology of this study follows Sames (Reference Sames and Sames2011c). Ostracod size ranges are as follows: very small: <0.40 mm; small: 0.40–0.50 mm; medium: 0.51–0.70 mm; large: 0.71–1.00 mm; very large: 1.01–2.00 mm; gigantic: >2.00 mm (Ayress and Whatley, Reference Ayress and Whatley2014). We use the following abbreviations: L = length; H = height; W = width; LV = left valve; RV = right valve.
Repository and institutional abbreviation
The illustrated specimens in this study are deposited in the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (NIGPAS) under the collection numbers NIGP 180012–180052.
Systematic paleontology
The classification above family level used herein follows Danielopol et al. (Reference Danielopol2018) and Meisch et al. (Reference Meisch, Smith and Martens2019). The taxonomy of lower ranks is based on Hou et al. (Reference Hou, Gou and Chen2002), Neustrueva et al. (Reference Neustrueva, Sinitsa, Khand and Melnikova2005), Sames (Reference Sames and Sames2011a, Reference Sames and Samesb), and Choi et al. (Reference Choi, Wang, Hu and Huh2020), although with revision that will be further discussed in this paper.
Class Ostracoda Latreille, Reference Latreille1802
Order Podocopida Sars, Reference Sars1866
Suborder Cypridocopina Baird, Reference Baird1845
Superfamily Cypridoidea Baird, Reference Baird1845
Family Cyprideidae Martin, Reference Martin1940
Genus Cypridea Bosquet, Reference Bosquet1852
Type species
Cypris granulosa Sowerby, Reference Sowerby and Fitton1836.
Cypridea verrucata Neustrueva, Reference Neustrueva, Kramarenko, Luvsandansan and Rozhdestvensky1974 emended
Figure 3
- Reference Neustrueva, Kramarenko, Luvsandansan and Rozhdestvensky1974
Cypridea verrucata n. sp., Neustrueva, p. 258, pl. 2, fig. 4a, b.
- Reference Neustrueva, Sinitsa, Khand and Melnikova2005
Cypridea verrucata Neustrueva in Neustrueva et al., pl. 4, fig. 3.
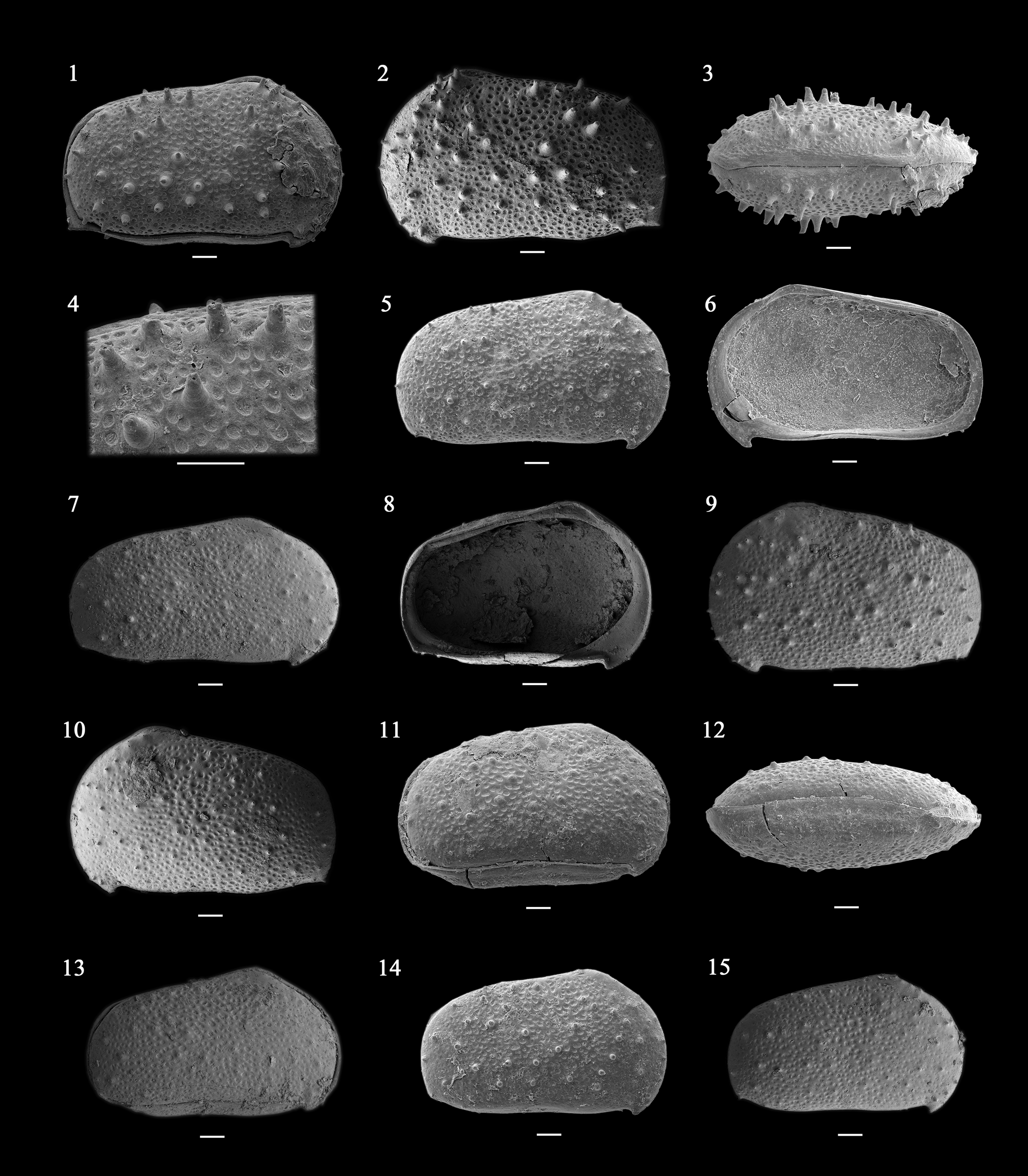
Figure 3. Cypridea verrucata (Neustrueva, Reference Neustrueva, Kramarenko, Luvsandansan and Rozhdestvensky1974). (1–4) Adult carapace, NIGP 180012, sample 5KC062111-2: (1) right view; (2) left view; (3) dorsal view (anterior end to the right); (4) detail of carapace surface. (5, 6) Adult right valve, NIGP 180013, sample 5KC062111-2: (5) lateral view; (6) internal view. (7) Lateral view of adult right valve, NIGP 180014, sample 5MT062111-9. (8, 9) Adult left valve, NIGP 180015, sample 5KC062111-2: (8) internal view; (9) lateral view. (10) Lateral view of adult left valve, NIGP 180016, sample 5MT062111-3. (11, 12) Adult carapace, NIGP 180017, sample 5MT062111-9: (11) right view; (12) ventral view (anterior end to the right). (13) Right view of A-1 instar carapace, NIGP 180018, sample 5MT062111-9. (14) Lateral view of A-1 instar right valve, NIGP 180019, sample 5MT062111-9. (15) Lateral view of A-1 instar right valve, NIGP 180020, sample 5MT062111-3. Scale bars = 100 μm.
Holotype
Specimen no. 2945/20 (deposited in the Institute of Precambrian Geology and Geochronology, Russian Academy of Sciences [IGGD], St. Petersburg, Russia) from the Lower Cretaceous Khukhteeg Formation, Mongolia (Neustrueva, Reference Neustrueva, Kramarenko, Luvsandansan and Rozhdestvensky1974, p. 258–259, pl. 2, fig. 4a, b).
Diagnosis (emended)
Very large Cypridea, valves somewhat asymmetrical in lateral view due to different shapes; subrectangular for LV and subovate for RV; LV larger than RV. Rostrum short but well defined. Alveolus (notch and furrow) short and indistinct. Selvage of larger LV overlapping RV along entire margin. Cyathus acute and distinct in LV. Surface covered by puncta and spine-like tubercles with four-forked ends.
Description
Carapace very large, subrectangular in LV and subovate in RV (in lateral view). Maximum length at mid-height, maximum height at one-third of length (anterior cardinal angle), and maximum width near mid-length. LV larger than RV and overlapping RV along entire margin. Anterior margin broad and slightly infracurvate; anterior cardinal angle (~140°) somewhat angular and protruding in both valves, and a node-like swelling developed in LV. Rostrum short but well defined, its point overreaching the ventral margin. Alveolar notch very shortly incised. Alveolar furrow shallow and faint (reaching up to a maximum of one-eighth of the height) in both valves. Selvage of larger LV overlapping RV along entire margin (Fig. 3.8). Posterior margin nearly equicurvate to slightly infracurvate and narrower than anterior one; posterior cardinal angle somewhat rounded to angular. Dorsal margin of both valves straight and declined toward the posterior end. In dorsal view, a slightly incised hinge margin forms a shallow dorsal furrow. Ventral margin almost straight but weakly concave; posteroventral area protruding with acute triangular cyathus in LV, but angular shape in RV. Surface covered by distinct puncta, spines, and spine-like tubercles; tip of spines displaying three- or four-forked ends. Normal pore canals form in muri and center of spines and spine-like tubercles.
Internal characters.—Hinge lophodont, a straight/smooth hinge bar with anterior and posterior sockets (elongate for anterior one and short for posterior one) in RV, with corresponding groove and anterior and posterior teeth in LV. Calcified inner lamella moderately broad along the anterior area with maximum width at anteroventral area and narrow in posterior area with maximum width at cyathus area. Striae identified along the free inner lamella near anterior inner margin. Selvage distinct in both anterior and posterior margins, but more strongly developed in anterior one, and interrupted selvage not identified (continuous selvage developed in posterior side of rostrum). Attached area narrow, and its point not protruding.
Morphologic variation.—The development of tubercles varies. Strongly developed tubercles show elongate, spine-like shapes with distinct four-forked ends with a normal pore (Fig. 3.4),whereas the centers of small or blunt-shaped tubercles have a smooth end with a normal pore (Fig. 3.15). These characters are local ornamentation elements related to ecophenotypic variation and/or ontogenetic character (see Sames, Reference Sames and Sames2011c). Sexual dimorphism is not observed in this study.
Ontogenetic variation.—Adults and A-1 instar are identified. Carapace length of an adult ranges from 0.9 to 1.11 mm, while the A-1 instar is about 0.71 mm (Fig. 3.15) long. Besides the development of tubercles (also possibly ecophenotypic variation) and size differences, no obvious morphologic differences have been observed between adult and A-1 instar stages.
Material
Four carapaces and 11 valves from samples 5MT062111-9, 5MT062111-3, and 5KC062111-2 of the Oshih Hollow East Section.
Dimensions
L: 0.71–1.11 mm; H: 0.58–0.68 mm; W: 0.41–0.62 mm.
Remarks
In general, the selvage is discontinuous and interrupted along the rostrum's posterior side of the representatives of the genus Cypridea (see Horne and Colin, Reference Horne and Colin2005, pl. 1, figs. 2, 4; Sames, Reference Sames and Sames2011b, fig. 7f). Although the species Cypridea verrucata has a continuous selvage in the anteroventral region (see Fig. 3.8), it still perfectly matches the diagnostic characters of the genus Cypridea. Therefore, this species is assigned to the genus Cypridea.
Cypridea concina Hou, Reference Hou1958 from the Lower Cretaceous Xiagou Formation of Gansu Province, China, is similar to C. verrucata, but the latter has a stronger valve asymmetry and a more distinct cyathus. In addition, Cypridea yanjiensis Gou, Reference Gou1983 from the Lower Cretaceous Tongfosi Formation of the Yanji Basin is similar in carapace outline to C. verrucata, but the former has a very rounded posterior margin and cyathus area.
Cypridea ihsienensis Hou, Reference Hou1958 emended
Figure 4.1, 4.2
- Reference Hou1958
Cypridea (Ulwellia) ihsienensis n. sp., Hou, p. 101, pl. 10, figs. 11–14.
- Reference Zhang, Zhang, Wu and Wu1985
Cypridea (Ulwellia) ihsienensis; Zhang, pl. 1, fig. 14.
- Reference Yang1985
Cypridea (Ulwellia) ihsienensis; Yang, p. 217, pl. 3, fig. 12.
- Reference Qi and Zhang1990
Cypridea (Ulwellia) ihsienensis; Qi and Zhang, pl. 3, figs. 1, 2.
- Reference Hou, Gou and Chen2002
Cypridea ihsienensis; Hou et al., p. 502, pl. 178, figs. 11–16.
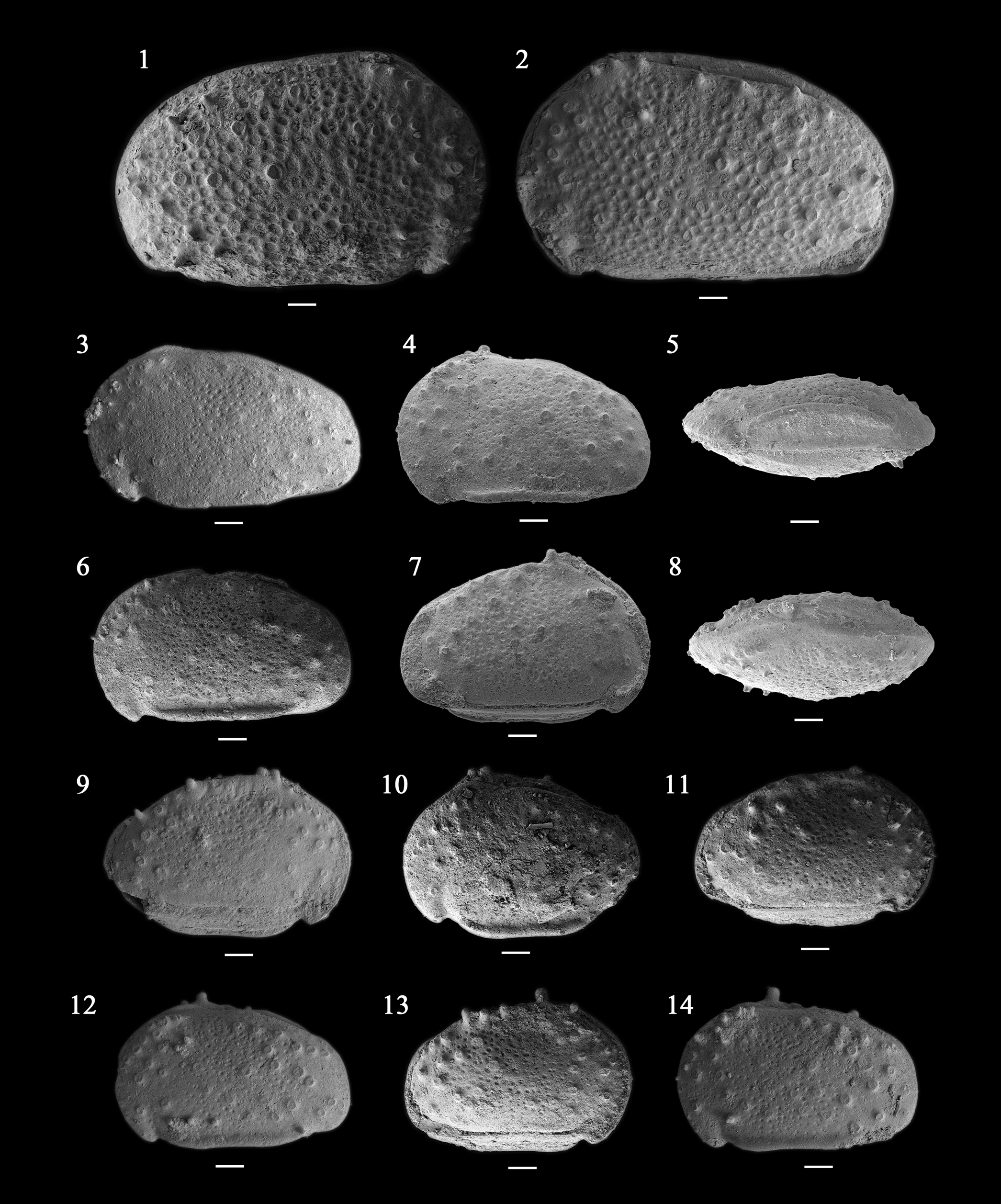
Figure 4. (1, 2) Cypridea ihsienensis (Hou, Reference Hou1958): (1) right view of carapace, NIGP 180021, sample 4TM062411-1; (2) left view of carapace, NIGP 180022, sample 4TM062411-1. (3–14) Cypridea unicostata (Galeeva, Reference Galeeva1955): (3) left view of carapace, NIGP 180023, sample 4TM062411-1; (4, 5) carapace, NIGP 180024, sample 4TM062411-1: (4) left view, (5) ventral view (anterior end to the left); (6) left view of carapace, NIGP 180025, sample 4TM062411-1; (7, 8) carapace, NIGP 180026, sample 4TM062411-1: (7) right view, (8) dorsal view (anterior end to the left); (9, 10) carapace, NIGP 180027, sample 4TM062411-1; (9) right view, (10) left view; (11) right view of carapace, NIGP 180028, sample 4TM062411-1; (12) left view of carapace, NIGP 180029, sample 4TM062411-1; (13, 14) carapace, NIGP 180030, sample 4TM062411-1: (13) right view, (14) left view. Scale bars = 100 μm.
Holotype
Specimen no. 9252 (deposited in the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences [NIGPAS], Nanjing, China) from the Lower Cretaceous Fuxin Formation, Liaoning Province, China (Hou, Reference Hou1958, p. 101–102, pl. 10, fig. 14).
Diagnosis (emended)
Very large Cypridea, subovate in lateral view; RV larger than LV, valve overlapping distinct in dorsal area of RV due to the dorsolateral extension. Rostrum short but broad in RV, and smaller with acute shape in LV. Alveolar notch short but well defined. Alveolar furrow elongated in RV, small and triangular in LV. Cyathus triangular in RV. Surface covered by puncta and conic tubercles.
Description
Carapace very large, subovate in lateral view. Maximum length at mid-height and maximum height at nearly one-third of length (anterior cardinal angle). RV larger than LV and overlapping LV along entire margin, especially along the dorsal margin (due to the dorsolateral extension). Anterior margin rounded and slightly infracurvate; anterior cardinal angle (~140°) somewhat rounded but distinct, with a lateral node-like swelling developed in RV. Rostrum short but broadly developed in RV, small and very acute in LV; rostrum point bending backward and slightly overreaching the ventral margin. Alveolar notch short but well defined. Alveolar furrow elongated and narrow in RV (reaching up to a maximum of one-fifth of the height), but much smaller and triangular in LV. Posterior margin slightly infracurvate and narrower than anterior margin; posterior cardinal angle obtuse rounded in both valves. Dorsal margin of both valves slightly declined toward posterior end; dorsal outline slightly convex to straight in RV, nearly straight in the LV. Ventral margin straight in both valves, but ventral outline of RV slightly convex by weakly developed ventral ridge; posteroventral area of RV somewhat protruded by triangular cyathus, but indistinct and rounded in LV. Surface covered by puncta tending toward reticulation, with conic tubercles particularly in anterolateral, posterocentral, posterolateral, and dorsolateral areas of carapace.
Material
Two carapaces from sample 4TM062411-1 of the Oshih Hollow East Section.
Dimensions
L: 1.39 mm; H: 0.88–0.89 mm; W: 0.58–0.68 mm.
Remarks
Sames (Reference Sames and Sames2011b, p. 381, and references therein) rejected the subgenus Ulwellia Anderson, Reference Anderson1939, and this opinion is followed here. Cypridea ihsienensis is similar to C. muriculata Zhang, Reference Zhang, Zhang, Wu and Wu1985 from the Lower Cretaceous Yixian Formation, but the latter has a much broader rostrum in LV. Cypridea justa Lübimova, Reference Lübimova1956 from the Zuunbayan Formation of Mongolia and the Hailaer Basin of China is similar to C. ihsienensis in rostrum shape, subovate outline, and ornamentation pattern. However, C. justa has more distinct anterior cardinal angle and a rounded cyathus area. According to Wang et al. (Reference Wang, Sames, Liao, Xi and Pan2017a), the specimens of C. justa in Lübimova (Reference Lübimova1956) are probably juveniles of C. beipiaoensis Cao, Reference Cao1999. Cypridea beipiaoensis from the Yixian Formation, northeast China, differs from C. ihsienensis by a strong overlap of the larger valve along the entire margin and angular cardinal angles. Cypridea subrectangular obtusovata Li and Cao, Reference Li and Cao2002 from the Chaoshui Basin, northwest China, resembles the outline of C. ihsienensis at first glance, but the former has a much-inflated ventral outline with a rounded cyathus area. Cypridea copulenta Lübimova, Reference Lübimova1956 from the Barunbayan Formation, Mongolia, has much larger rostrum in the RV than C. ihsienensis, and its point is strongly overreaching the ventral margin.
Cypridea unicostata Galeeva, Reference Galeeva1955 emended
Figure 4.3–4.14
- Reference Galeeva1955
Cypridea unicostata n. sp., Galeeva, p. 34, pl. 4, fig. 2a–d.
- Reference Lübimova1956
Cypridea unicostata; Lübimova, pl. 7, fig. 2a–c.
- Reference Hao, Su, Li, Ruan and Yuan1974
Cypridea unicostata; Hao et al., p. 34, pl. 9, fig. 1a–g.
- non 1976
Cypridea unicostata; DOFEAD, p. 33, pl. 3, figs. 2a–4b.
- Reference Zhang, Zhang, Wu and Wu1985
Cypridea (Cypridea) unicostata; Zhang, pl. 1, fig. 6.
- Reference Yang1985
Cypridea unicostata; Yang, p. 217, pl. 3, fig. 11.
- non Reference Shou1985
Cypridea unicostata; Shou, p. 59, pl. 2, fig. 9.
- Reference Qi1988
Cypridea unicostata; Qi, p. 130, pl. 3, figs. 1–3.
- Reference Sinitsa1993
Cypridea unicostata; Sinitsa, p. 201, pl. 1, figs. 3, 4.
- Reference Nikolaeva and Neustrueva1999
Cypridea unicostata; Nikolaeva and Neustrueva, pl. 8, fig. 2.
- non Reference Ye, Huang, Zhang and Chen2002
Cypridea unicostata; Ye et al., p. 148, pl. 2, fig. 3a–c.
- Reference Hou, Gou and Chen2002
Cypridea unicostata; Hou et al., p. 452, pl. 166, figs. 20, 21.
- Reference Neustrueva, Sinitsa, Khand and Melnikova2005
Cypridea unicostata; Neustrueva et al., pl. 4, figs. 4–6, pl. 5, figs. 1–4.
Holotype
Specimen no. 200-16 (deposited in the All-Russia Petroleum Research Exploration Institute [VNIGRI], St. Petersburg, Russia) from the Lower Cretaceous Zuunbayan Formation, Mongolia (Galeeva, Reference Galeeva1955, p. 34–35, pl. 4, fig. 2a–d.).
Diagnosis (emended)
Large Cypridea, subovate to subtriangular in lateral view; LV overlaps RV along all margins except the hinge margin; a well-developed ventrolateral ridge presents in the ventral margin area of LV. Rostrum short but broad and stout. Alveolus moderately to well developed; alveolar notch broad, alveolar furrow crescent shaped in LV and indistinct in RV. Cyathus rounded but distinctly developed in LV but indistinct in RV. Surface covered by fine puncta and cylindric tubercles.
Description
Carapace large, subovate to subtriangular in lateral view. Maximum length at one-fifth of height near the cyathus area, maximum height at nearly one-third of length (anterior cardinal angle), and maximum width at mid-length. LV larger than RV and overlapping RV along entire margin. Anterior margin broad, weakly equicurvate to infracurvate; anterior cardinal angle (~150°) rounded but protruding in LV with an indistinct lateral node-like swelling, but nearly rounded in RV. Rostrum short but broad and stout, its point somewhat blunt and not reaching the ventral outline. Alveolar notch short but moderately broad. Alveolar furrow slightly crescent and moderately incised in LV, reaching up to one-fourth of height, but smaller in RV and more indistinct than the former (reaching up to a maximum of one-fifth of the height). Posterior margin strongly infracurvate and tapering in LV but equicurvate to slightly infracurvate with narrow margin in RV; posterior cardinal angle area somewhat angular in LV and rounded in RV. Dorsal margin convex and declined toward the posterior end; dorsal outline prominent and more distinctive in LV due to the dorsal ridge. In dorsal view, hinge margin incised between anterior and posterior cardinal angle areas, forming a dorsal furrow. Ventral margin and outline nearly straight in RV, inflated ventral outline in LV with very well-developed ventrolateral ridge; posteroventral area narrowly rounded by cyathus in LV, but moderately rounded in RV. Surface covered by fine puncta and cylindrical tubercles of variable size.
Morphologic variation.—Some specimens show very inflated ventral outline. However, this feature is identified only in LV. Thus, specific characters for male/female distinction in Cypridea unicostata are not observed.
Ontogenetic variation.—It is unclear due to the narrow range of sizes between specimens and because internal characters are not observed.
Material
Eleven carapaces from sample 4TM062411-1 of the Oshih Hollow East Section.
Dimensions
L: 0.86–0.96 mm; H: 0.52–0.58 mm; W: 0.37–0.43 mm.
Remarks
The strong valve asymmetry combined with the remarkably inflated ventral outline and ventrolateral ridge of LV in C. unicostata are very distinguishable from the morphology of other Cypridea species. Cypridea figurata Neustrueva, Reference Neustrueva, Kramarenko, Luvsandansan and Rozhdestvensky1974 from the Shinekhudag Formation (Neustrueva et al., Reference Neustrueva, Sinitsa, Khand and Melnikova2005) is similar to C. unicostata, but the former has a narrower alveolus (notch and furrow), a distinct and well-developed dorsal ridge between anterior and posterior cardinal angles, as well as development of spine-like tubercles. Cypridea bicostata Wang (in Qi and Wang, Reference Qi and Wang1981) from the Lower Cretaceous Xinmingpu Group resembles C. unicostata in outline and shape of the ventrolateral ridge, but the valve overlap of the former is more distinctive, and the cyathus area is smooth and more rounded than in C. unicostata. In addition, Cypridea liaoningensis Zhang, Reference Zhang, Zhang, Wu and Wu1985 from the Yixian Formation has a similar carapace outline and some features of C. unicostata, but C. liaoningensis has ventrolateral ridges and anterior swellings in each valve, and the presumed functional significances of these features are upside-down swimming and eye tubercles, respectively (see Wang et al., Reference Wang, Sha and Pan2013, p. 106).
Cypridea tumefacta Neustrueva, Reference Neustrueva, Kramarenko, Luvsandansan and Rozhdestvensky1974
Figure 5.1
- Reference Neustrueva, Kramarenko, Luvsandansan and Rozhdestvensky1974
Cypridea tumefacta n. sp., Neustrueva, p. 260, pl. 3, fig. 1a–c.
- Reference Neustrueva1989
Langtonia tumefacta; Neustrueva, pl. 1, figs. 1, 2.
- Reference Nikolaeva and Neustrueva1999
Langtonia tumefacta; Nikolaeva and Neustrueva, pl. 9, figs. 1, 2.
- Reference Neustrueva, Sinitsa, Khand and Melnikova2005
Langtonia tumefacta; Neustrueva et al., pl. 13, figs. 1–3.

Figure 5. (1) Cypridea tumefacta (Neustrueva, Reference Neustrueva, Kramarenko, Luvsandansan and Rozhdestvensky1974), left view of valve, NIGP 180031, sample 5MT062111-9. (2, 3) Yumenia cf. Y. oriformis (Hou, Reference Hou1958): (2) right view of adult valve, NIGP 180032, sample 4UK070411-2; (3) right view of ?A-2 instar valve, NIGP 180033, sample 4UK070411-1. (4, 5) Scabriculocypris subscalara (Zhang and Chen in Ye et al., Reference Ye, Huang, Liu, Zhang and Chen2003), adult carapace, NIGP 180034, sample 4UK070411-1: (4) left view; (5) right view. (6, 7) ?Trapezoidella sp.: (6) left view of valve, NIGP 180035, sample 5MT062111-3; (7) left view of internal mold, NIGP 180036, sample 5KC062111-2. (8) Candona sp., lateral view of right valve, NIGP 180037, sample 5MT062111-9. Scale bars = 100 μm.
Holotype
Specimen no. 2945/27 (deposited in the Institute of Precambrian Geology and Geochronology, Russian Academy of Sciences [IGGD], St. Petersburg, Russia) from the Lower Cretaceous Zuunbayan Formation, Mongolia (Neustrueva, Reference Neustrueva, Kramarenko, Luvsandansan and Rozhdestvensky1974, p. 260–261, pl. 3, fig. 1a–c).
Material
One partial left valve from sample 5MT062111-9 of the Oshih Hollow East Section.
Remarks
Despite many missing and crushed parts (partial anterior, dorsal, and posterior areas), the low L/H ratio and rostrum/alveolus shapes of the studied specimen fit well into Cypridea tumefacta Neustrueva, Reference Neustrueva, Kramarenko, Luvsandansan and Rozhdestvensky1974 (also known as Langtonia tumefacta in some literature, e.g., Nikolaeva and Neustrueva, Reference Nikolaeva and Neustrueva1999). However, this species shows typical features of the representatives of the Cypridea laevigata/setina group (see more details in Sames, Reference Sames and Sames2011b, p. 396–397). Although their affinities are controversial and unsolved so far (Schudack and Schudack, Reference Schudack and Schudack2009; Sames, Reference Sames and Sames2011b; Trabelsi et al., Reference Trabelsi, Colin and Touir2011, Reference Trabelsi, Sames, Salmouna, Piovesan, Ben Rouina, Houla, Touir and Soussi2015), the representatives of the C. laevigata/setina group have a completely smooth carapace surface. Rostrum/alveolus size ranges and shapes are variable in this species group (e.g., well-developed rostrum/alveolus of C. ex gr. laevigata in Trabelsi et al., Reference Trabelsi, Sames, Salmouna, Piovesan, Ben Rouina, Houla, Touir and Soussi2015); however, many Early Cretaceous species have a small rostrum and small/inconspicuous alveolus (see Anderson, Reference Anderson1985; Schudack and Schudack, Reference Schudack and Schudack2009; Sames, Reference Sames and Sames2011b). Cypridea tumefacta herein also has a totally smooth surface and small rostrum/alveolar notch; therefore, this species is considered a representative of the C. laevigata/setina group. Among this species group, some European species with a low L/H ratio (C. laevigata var. camelodes and C. laevigata var. dotica in Anderson, Reference Anderson1962) are similar to C. tumefacta. However, European variants of C. laevigata have a much smaller carapace size and rostrum (see also Anderson, Reference Anderson1985).
Family Alloiocyprideidae Ayress and Whatley, Reference Ayress and Whatley2014
Genus Yumenia Hou, Reference Hou1958
Type species
Cypridea (Yumenia) oriformis Hou, Reference Hou1958.
Remarks
Hou (Reference Hou1958) established the new subgenus Cypridea (Yumenia). However, because it lacks a rostrum and alveolar notch, this subgenus has been excluded from the genus Cypridea as well as from the family Cyprideidae (e.g., Horne and Colin, Reference Horne and Colin2005; Sames, Reference Sames and Sames2011b). Yumenia has been placed into the Trapezoidellidae Sohn, Reference Sohn1979 (Nikolaeva and Neustrueva, Reference Nikolaeva and Neustrueva1999) or the Alloiocyprideidae Ayress and Whatley, Reference Ayress and Whatley2014 (Wang et al., Reference Wang, Yang, Pan and Liao2017b). Many representatives of the family Trapezoidellidae are, however, very different from Yumenia in carapace shape and local and area-wide ornamentation elements. Here we follow the view of Wang et al. (Reference Wang, Yang, Pan and Liao2017b) that Yumenia is assigned to the family Alloiocyprideidae.
The representatives of Yumenia are quite similar to those of the genus Scabriculocypris Anderson, Reference Anderson1941 in general morphology, such as typical Cypridea-like carapace shapes (subrectangular to subtrapezoidal in lateral view) but lacking rostrum/alveolus and cyathus. The maximum heights of both genera are usually at the anterior cardinal angle. In addition, except for some taxa, the area-wide ornamentation elements in many representatives of both genera are very similar (reticulation or punctuation). Therefore, Yumenia is potentially synonymous with Scabriculocypris. However, some Yumenia species (including questionable species) in previous studies show more various morphology and ornamentation types (e.g., smooth surface of ?Y. acutiuscula (Zhang, Reference Zhang, Zhang, Wu and Wu1985) in Wang et al., Reference Wang, Sha, Pan and Zuo2016 and large spines of ?Y. ordinata (Ye in Hao et al., Reference Hao, Su, Li, Ruan and Yuan1974) in Wang et al., Reference Wang, Yang, Pan and Liao2017b). The ornamentation trends in Scabriculocypris species consist of reticulation/punctation and may or may not have local ornamentation elements. Many of these characters are insignificant to divide two genera; hence, the phylogenetic relationship between Yumenia and Scabriculocypris needs further investigation.
Yumenia cf. Y. oriformis Hou, Reference Hou1958
Figure 5.2, 5.3
- Reference Hou1958
Cypridea (Yumenia) oriformis n. sp., Hou, p. 94, pl. 9, figs. 1–5.
- Reference Hou1958
Cypridea (Yumenia) suboriformis n. sp., Hou, p. 95, pl. 9, figs. 6–9, 11.
- Reference Hou1958
Cypridea (Yumenia) pentanoda n. sp., Hou, p. 95, pl. 9, figs. 10, 12–14.
- Reference Hou, Gou and Chen2002
Yumenia oriformis; Hou et al., p. 611, pl. 254, figs. 1, 2.
Holotype
Specimen no. 9224 (deposited in the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences [NIGPAS], Nanjing, China) from the Lower Huihuipu Series, China (Hou, Reference Hou1958, p. 94–95, pl. 9, fig. 1).
Description
Valve large, subtriangular in lateral view. Maximum length at one-third of height and maximum height at near one-third of length (anterior cardinal angle). Valve overlapping unknown (in Hou, Reference Hou1958, LV larger than RV). Anterior margin broadly rounded and weakly infracurvate to equicurvate; anterior cardinal angle (~140°) angular and distinct. Posterior margin much narrower than anterior one and infracurvate; posterior cardinal angle nearly rounded and inconspicuous. Dorsal margin straight and deeply declined toward the posterior end. Ventral margin almost straight but weakly concave in near mid-length. Surface covered by fine puncta, with small tubercles particularly in anterolateral and posterolateral areas or totally absent.
Ontogenetic variation.—Presumed A-2 instar (0.80 mm long) is smaller than the adult (1.02 mm long). The morphologic characters in a presumed A-2 instar are the same as in the adult.
Material
Three valves from samples 4UK070311-3, 4UK070411-1, and 4UK070411-2 of the Tsagaan Tsuvarga Section.
Dimensions
L: 0.80–1.02 mm; H: 0.52–0.71 mm; W: unknown.
Remarks
The classification of this species is based only on several valves. Thus, the species assignment is questionable. However, on the basis of general shape and local/area-wide ornamentation pattern, we presume that the studied specimens are related to or same taxon of Yumenia oriformis Hou, Reference Hou1958.
Some representatives of the genus Yumenia in Hou (Reference Hou1958) are likely synonymous with Y. oriformis as follows: Yumenia suboriformis Hou, Reference Hou1958 has concave ventral margin, and Y. pentanoda Hou, Reference Hou1958 has a more slender carapace than Y. oriformis, but other features fit the features of the latter. Thus, these species are considered synonyms in this study.
Yumenia remota (Lübimova, Reference Lübimova1956) in Nikolaeva and Neustrueva (Reference Nikolaeva and Neustrueva1999) shows strong similarities to Y. oriformis in carapace shape and ornamentation. The original illustrations and photos of Y. remota in Lübimova (Reference Lübimova1956) and Neustrueva et al. (Reference Neustrueva, Sinitsa, Khand and Melnikova2005), however, show a more trapezoidal shape with convex ventral margin in both valves. Janinella tsaganensis Neustrueva, Reference Neustrueva1977 from the Shinekhudag Formation is similar to Yumenia cf. Y. oriformis in carapace shape and ornamentation, but the former exhibits a small rostrum-like structure and slightly incised alveolar notch in both valves.
Genus Scabriculocypris Anderson, Reference Anderson1941
Type species
Scabriculocypris trapezoides Anderson, Reference Anderson1941.
Scabriculocypris subscalara Zhang and Chen in Ye et al., Reference Ye, Huang, Liu, Zhang and Chen2003
Figure 5.4, 5.5
- Reference Ye, Huang, Liu, Zhang and Chen2003
Limnocypridea subscalara n. sp., Zhang and Chen in Ye et al., p. 95, pl. 18, fig. 5a–c; pl. 20, figs. 1, 2, 4.
- Reference Ye, Huang, Liu, Zhang and Chen2003
Limnocypridea huangqimiaoensis n. sp., Huang in Ye et al., p. 95, pl. 20, fig. 3a, b; pl. 21, fig. 1a–c.
- Reference Liu, Huang and Ye2004
Limnocypridea subscalara; Liu et al., pl. 2, fig. 3a, b.
Holotype
Specimen no. DQH191 (deposited in the Institute of Petroleum Exploration and Exploitation, Division Company of Corporation Ltd., Daqing, China) from the Nantun Formation of the Hailaer Basin, China (Ye et al., Reference Ye, Huang, Liu, Zhang and Chen2003, p. 95).
Description
Carapace very large, subtrapezoidal in lateral view. Maximum length at mid-height and maximum height at one-third of length (anterior cardinal angle). LV larger than RV and overlapping RV entire margin, especially along the dorsal margin by the dorsal ridge. Anterior margin equicurvate; anterior cardinal angle (~140°) rounded and indistinct in both valves. Posterior margin rounded and infracurvate; posterior cardinal angle somewhat more angular than anterior one. Dorsal margin straight and slightly declined toward the posterior end. Ventral margin straight in LV but concave in mid-length of RV. Surface covered by puncta tending toward reticulation. Local ornamentation elements absent.
Material
One carapace from sample 4UK070411-1 from the Tsagaan Tsuvarga Section.
Dimensions
L: 1.33 mm; H: 0.86 mm; W: Unknown (due to the compressed carapace).
Remarks
This species was formerly assigned to the genus Limnocypridea. However, it differs from trapezoidellids by strongly inclined dorsal margin, a somewhat weaker dorsal overlap, and a reticulation-like punctation. Therefore, we transfer this species to the genus Scabriculocypris. L. huangqimiaoensis Huang (in Ye et al., Reference Ye, Huang, Liu, Zhang and Chen2003) differs from S. subscalara by only the elongated carapace shape, but other features fit the diagnostic characters of the latter. Furthermore, these two species have been reported from the same strata and basin. Thus, L. huangqimiaoensis is considered a junior synonym of S. subscalara.
Genus Trapezoidella Sohn, Reference Sohn1979
Type species
Bythocypris (Bairdiocypris) trapezoidalis Roth, Reference Roth1933.
?Trapezoidella sp.
Figure 5.6, 5.7
Material
One partial valve and one internal mold from samples 5MT062111-3 and 5KC062111-2 of the Oshih Hollow East Section.
Dimensions
L: 1.90 mm; H: 1.20 mm; W: unknown.
Remarks
This species is distinguished from other species by (presumed) gigantic size and thick/elongate lateral ridge, but the details of the species are very limited due to the fragmentary material. However, a distinct lateral ridge and smooth carapace are relatively common in the genus Trapezoidella (see Sohn, Reference Sohn1979). Therefore, here we tentatively assign this taxon to the species of Trapezoidella. Some species are partially similar to ?Trapezoidella sp. in this study but differ in the size and other characters; Trapezoidella hornei Khand, Sames, and Schudack, Reference Khand, Sames and Schudack2007 has also been recorded from the Shinekhudag Formation, but this species shows a thin lateral ridge, punctate area-wide ornamentation elements, and a much smaller carapace size (maximum 1.13 mm long). Thus the specimens of T. hornei in Khand et al. (Reference Khand, Sames and Schudack2007) could be juvenile forms of ?Trapezoidella sp. in this study. However, this view needs further investigation (due to sparse data). Trapezoidella rothi Sohn, Reference Sohn1979 from the Lower Cretaceous Lakota Formation, USA, shows a smooth carapace, but the lateral ridges are more displaced toward the ventral margin than in ?Trapezoidella sp. The elongate and thick lateral ridge is also confirmed in several trapezoidellid species, such as Falklandicypris petrasaltata Ayress and Whatley, Reference Ayress and Whatley2014 from the North Falkland Basin, South Atlantic, and Limnocypridea bitumulosa Lübimova, Reference Lübimova1956 from Mongolia. However, except for the existence of a lateral ridge, they are totally different in shape.
Family Candonidae Kaufmann, Reference Kaufmann1900
Genus Candona Baird, Reference Baird1845
Description
Valve large, elongated and ovate in lateral view. Maximum length at slightly below mid-height and maximum height near three-fifths of length (posterior cardinal angle). Anterior margin rounded and nearly equicurvate; anterior cardinal angle rounded and very inconspicuous. Posterior margin much narrower than anterior one and equicurvate to slightly supracurvate. Dorsal margin somewhat convex and rounded. Ventral margin almost straight. Surface smooth.
Material
One valve from sample 5MT062111-9 of the Oshih Hollow East Section.
Dimensions
L: 0.71 mm; H: 0.40 mm; W: unknown.
Remarks
The studied material includes only a single valve; therefore, more specimens are required for more detailed investigation and comparison. However, the small valve and elongate/ovate shape with convex dorsal margin of this specimen are typical of the representatives of the genus Candona in much of the literature. The general outline of this species is somewhat to that of Candona sp. 1 from the Tongfosi Formation of the Yanji Basin (Choi et al., Reference Choi, Wang, Hu and Huh2020). However, the latter has a broad posterior area (beyond the posterior cardinal angle) and a concave ventral margin.
Superfamily Cytheroidea Baird, Reference Baird1850
Family Limnocytheridae Sars, Reference Sars and Sars1928
Subfamily Limnocytherinae Sars, Reference Sars and Sars1928
Genus Vlakomia Gramm, Reference Gramm1966 emended
- Reference Gramm1966
Vlakomia, Gramm, p. 83.
- Reference Sinitsa1993
Tsetsenia, Sinitsa, p. 223.
Type species
Vlakomia ustinovskii Gramm, Reference Gramm1966.
Included species
Vlakomia ustinovskii Gramm, Reference Gramm1966; Vlakomia jilinensis Gou, Reference Gou1983; Vlakomia ulanense Neustrueva, Reference Neustrueva1977 (= Theriosynoecum ulanense Neustrueva, Reference Neustrueva1977) (wrongly described as Tsetsenia ulanensis by Neustrueva et al., Reference Neustrueva, Sinitsa, Khand and Melnikova2005); Vlakomia mira Sinitsa, Reference Sinitsa1993 (= Tsetsenia mira Sinitsa, Reference Sinitsa1993); ?Vlakomia temperata Sinitsa, Reference Sinitsa1993 (= Tsetsenia temperata Sinitsa, Reference Sinitsa1993).
Diagnosis (emended)
Small to large carapace (typically smaller than 0.80 mm), oblong and rhomboidal to subrhomboidal in lateral view. Anterior margin broad with a remarkable anterior cardinal angle. LV larger than RV and weakly overlapping RV along entire margin. Two sulci developed in anterodorsal area, one of them barely to weakly developed in some species. Lateral pore canals with simple and type C sieve pores. Hinge lophodont with strong anterior and posterior teeth/sockets, posterior tooth roundish, anterior tooth roundish or divided into three tiny and shallow teeth, hinge line straight. Surface reticulation. Common presence of large hollow tubercles with constant position and smaller tubercles with no fixed location. Sexual dimorphism visible; precocious sexual dimorphism also seen. Male elliptical in dorsal view and oblong in lateral view. Female rhomboidal in dorsal view with more rounded and broader posterior margin. Male always larger than female. Female with stronger ornamentation.
Occurrence
‘Bon-Tsagaan Series’ deposits and Shinekhudag Formation of Central and Gobi–Altai regions of Mongolia, Barremian–Aptian; Tongfosi Formation of the Yanji Basin in Northeast China, Albian (Sinitsa, Reference Sinitsa1993; Choi et al., Reference Choi, Wang, Hu and Huh2020; Zhong et al., Reference Zhong, Wang, Jia, Wang, Hu and Pan2021, and this study).
Paleoecology
Nonmarine, fresh to (hyper-)saline water environment (Choi et al., Reference Choi, Wang, Hu and Huh2020).
Remarks
The genus Tsetsenia Sinitsa, Reference Sinitsa1993 is considered a junior synonym of Vlakomia because the type species of the former genus fits the diagnostic features of Vlakomia (general shape, two dorsolateral sulci, and reticulation) very well. Although Sinitsa (Reference Sinitsa1993) tentatively assigned her genus Tsetsenia to the family Limnocytheridae, Nikolaeva and Neustrueva (Reference Nikolaeva and Neustrueva1999) and Neustrueva et al. (Reference Neustrueva, Sinitsa, Khand and Melnikova2005) clearly stated that the genus Tsetsenia belongs to the family Limnocytheridae.
The family Limnocytheridae comprises two subfamilies, the subfamily Limnocytherinae and the subfamily Timiriaseviinae (Sames, Reference Sames and Sames2011a; Danielopol et al., Reference Danielopol2018). The taxonomy above species level of this family has been comprehensively discussed in several papers (e.g., Colin and Danielopol, Reference Colin and Danielopol1978; Sames, Reference Sames and Sames2011a; Danielopol et al., Reference Danielopol2018 and references therein). Table 1 lists important carapace features to compare and differentiate several Late Mesozoic members of the family Limnocytheridae, including Vlakomia Gramm, Reference Gramm1966, Theriosynoecum Branson, Reference Branson1936, Timiriasevia Mandelstam, Reference Mandelstam1947, and Metacypris Brady and Robertson, Reference Brady and Robertson1870. Among them, Vlakomia belongs to the subfamily Limnocytherinae while the other three genera are considered members of the subfamily Timiriaseviinae (Danielopol et al., Reference Danielopol2018; Choi et al., Reference Choi, Wang, Hu and Huh2020). In fossil representatives of the Timiriaseviinae, the female brood pouches are always present and mostly strongly developed (showing visible sexual dimorphism) (Sames, Reference Sames and Sames2011a). The members of Limnocytherinae do not develop female brood pouches, and the male carapace is often larger than the female one (Meisch, Reference Meisch, Schwoerbel and Zwick2000). In summary, the genus Vlakomia can be well determined on the basis of carapace characters and can be clearly distinguished from the genera Theriosynoecum, Timiriasevia, and Metacypris as well (see Table 1 for details).
Table 1. Comparison of taxonomically significant carapace features (taxonomic key) of selected genera from the family Limnocytheridae and their stratigraphic distribution (modified from Sames, Reference Sames and Sames2011a, table 1).

V. pristina Zhang, Reference Zhang1982 is invalid since this species was established on the basis of one broken specimen without a shell (Choi et al., Reference Choi, Wang, Hu and Huh2020). ?Vlakomia temperata (Sinitsa, Reference Sinitsa1993) is questionable here because the figured type specimen of this species seems to be deformed by compression (see Sinitsa, Reference Sinitsa1993, pl. 16, fig. 5).
Vlakomia ulanense Neustrueva, Reference Neustrueva1977
Figure 6
- Reference Neustrueva1977
Theriosynoecum ulanense, Neustrueva, p. 140, pl. 2, figs. 14, 15.
- Reference Neustrueva, Sinitsa, Khand and Melnikova2005
Tsetsenia ulanensis, Neustrueva et al., p. 55, pl. 26, figs. 7, 8.

Figure 6. Vlakomia ulanense (Neustrueva, Reference Neustrueva1977). (1) Lateral view of adult male right valve, NIGP 180038, sample 4UK070311-1. (2) Lateral view of adult male right valve, NIGP 180039, sample 4UK070311-2. (3) Lateral view of A-1 instar male left valve, NIGP 180040, sample 4UK070311-1. (4) Lateral view of adult female right valve, NIGP 180041, sample 4UK070311-3. (5) Internal view of female adult right valve (part), NIGP 180042, sample 4UK070311-2. (6) Lateral view of adult female left valve, NIGP 180043, sample 4UK070411-2. (7) Lateral view of adult female right valve with anterolateral part missing, NIGP 180044, sample 4UK070311-2. (8) Lateral view of adult female left valve, NIGP 180045, sample 4UK070411-2. (9) Lateral view of adult female right valve, NIGP 180046, sample 4UK070411-2. (10) Lateral view of A-1 instar female left valve, NIGP 180047, sample 4UK070411-2. (11) Lateral view of A-2 instar left valve, NIGP 180048, sample 4UK070311-2. (12) Lateral view of A-2 instar right valve, NIGP 180049, sample 4UK070311-2. (13) Lateral view of A-2 instar left valve, NIGP 180050 (broken), sample 4UK070311-2. (14) Lateral view of A-3 instar left valve, NIGP 180051, sample 4UK070311-2. (15) Lateral view of A-3 instar right valve, NIGP 180052, sample 4UK070411-2. Scale bars = 100 μm.
Holotype
Specimen no. 3589/12 (deposited in the Paleontological Institute, Russian Academy of Sciences [PIN], Moscow, Russia) from the Andaikhudag Formation (= Shinekhudag Formation; see Khand et al., Reference Khand, Badamgarav, Ariunchimeg, Barsbold, Okada and Mateer2000), Mongolia (Neustrueva, Reference Neustrueva1977, p. 140–141, pl. 2, figs. 14, 15).
Description
Carapace medium to large, suboblong to subrhomboidal in lateral view. Maximum length near mid-height, and maximum height at one-fourth of length near anterior cardinal angle. LV larger than RV and slightly overlapping RV along all free margins. Anterior margin broadly rounded and infracurvate to slightly equicurvate; anterior cardinal angle remarkable (145° to 150° in male; 130° to 150° in female). Posterior margin in male broadly rounded but somewhat narrower than anterior one, equicurvate to slightly supracurvate; supracurvate in female. Hinge margin straight and slightly or deeply declined toward the posterior end. Dorsolateral sulcus weakly incised behind anterior cardinal angle area. Ventral margin straight to slightly convex in females and moderately to strongly concave near mid-length in males. Surface covered by reticulation-like punctation, except on the surface of nodes and in the central area of both valves where coincident with the muscle-scar field area in internal view. The pattern of puncta consists of round (globular to elongate) to polygonal shape. Two nodes: a round node in the ventrolateral area near the posteroventral region and more visible in the female; a ridge-like node in the dorsolateral area near the posterior cardinal angle, with broadly inflated (male) to somewhat convex (female) shape.
Internal characters.—Adductor muscle scars (AMS) consisting of a vertical row of four closely placed elongate scars, two scars (frontal and mandibular scars) situated just in front of AMS (Fig. 6.5). Hinge lophodont. Calcified inner lamella moderately broad; selvage and fused zone well visible along the anterolateral margin.
Sexual dimorphism.—Sexual dimorphism presents in A-1 instars and adults. Males have larger and more elongated carapaces/valves. The ridge-like node is more visible in males and somewhat overreaches the dorsal outline. Female is smaller than male, with broader anterior margin and the more visible ventrolateral node, the ridge-like node not or slightly overreaching the dorsal outline.
Ontogenetic variation.—The adult specimens are more than 0.60 mm long, but males are much larger (0.68 to 0.71 mm long) than females (0.60 to 0.62 mm long). A-1 instar (about 0.58 mm long) shares similar carapace features with the adult. The carapace shape of A-2 and A-3 instars (0.40 to 0.52 mm long) is nearly subrhomboidal to subtriangular due to narrow posterior margin and deeply declined hinge margin. Ridge-like nodes are less developed or absent in A-2 and A-3 instars. The separation of instars is based on size and general shape (Choi et al., Reference Choi, Wang, Hu and Huh2020).
Material
Nineteen valves and one broken carapace from samples 4UK070311-3, 4UK070311-2, 4UK070311-1, and 4UK070411-2 of the Tsagaan Tsuvarga Section.
Dimensions
Adult L: 0.60–0.71 mm; H: 0.37–0.41 mm; W: 0.31 mm. Juvenile L: 0.40–0.58 mm; H: 0.23–0.34 mm; W: Unknown.
Remarks
Although Neustrueva et al. (Reference Neustrueva, Sinitsa, Khand and Melnikova2005) transferred the species Theriosynoecum ulanense of Neustrueva (Reference Neustrueva1977) to the genus Tsetsenia without any explanation, they (Neustrueva et al., Reference Neustrueva, Sinitsa, Khand and Melnikova2005) reillustrated type material of this species with much better-quality SEM photographs (wrongly described as Tsetsenia “ulanensis” therein). Our specimens fit all diagnostic features of “Tsetsenia ulanensis” (= Theriosynoecum ulanense). Meanwhile, Tsetsenia is considered a junior synonym of Vlakomia in this study (see the preceding discussion). Thus, we identify our specimens as Vlakomia ulanense.
Vlakomia ulanense is similar to V. jilinensis from the Tongfosi Formation of the Yanji Basin of northeast China (Gou, Reference Gou1983; Choi et al., Reference Choi, Wang, Hu and Huh2020), but the former has a different node distribution pattern and lesser degree of ornamentation development. The general pattern of nodes, spines, and punctation is a useful characteristic in specific levels, although some ontogenetic, sexual, and ecophenotypic variability can be found. Despite some details in the shape, V. jilinensis is relatively similar to V. ulanense, by trend younger than the latter, and thus considered to be closely related and possibly deriving from the Mongolian lineage designated as V. ulanense. Vlakomia ustinovskii differs from V. ulanense and V. jilinensis in having developed large “horn-like” anterocentral nodes and having a visible anterior cardinal angle in both sexes. V. jilinensis and V. ustinovskii both derive from the Albian Tongfosi Formation; however, the horizon from which V. jilinensis came is lower than that from which V. ustinovskii came. Thus, we propose a possible phylogenetic lineage: V. ulanense–V. jilinensis–V. ustinovskii. This lineage is endemic to East Asia to date.
Discussion
Ostracod biostratigraphy and paleobiogeography
The “Purbeck–Wealden type” ostracod faunas from Mongolia had been reviewed by Nikolaeva and Neustrueva (Reference Nikolaeva and Neustrueva1999), Khand et al. (Reference Khand, Badamgarav, Ariunchimeg, Barsbold, Okada and Mateer2000), and Neustrueva et al. (Reference Neustrueva, Sinitsa, Khand and Melnikova2005). Khand et al. (Reference Khand, Badamgarav, Ariunchimeg, Barsbold, Okada and Mateer2000) proposed a “Cypridea fasciculata Zone” for the Shinekhudag Formation in the Eastern Gobi and Choir–Nyalga basins, which includes ostracod species of the genera Cypridea, Lycopterocypris, Rhinocypris, and Darwinula. In addition, Neustrueva et al. (Reference Neustrueva, Sinitsa, Khand and Melnikova2005) established two assemblages for the lower and upper parts of the Shinekhudag Formation in southeast Mongolia: the Cypridea unicostata–Cypridea figurata assemblage and the Janinella tsaganensis–Cypridea selenginensis–Langtonia tumefacta (= Cypridea tumefacta in this study) assemblage, respectively. On the basis of biostratigraphic correlation, Khand et al. (Reference Khand, Badamgarav, Ariunchimeg, Barsbold, Okada and Mateer2000) and Neustrueva et al. (Reference Neustrueva, Sinitsa, Khand and Melnikova2005) concluded that the age of the Shinekhudag Formation in southeastern and eastern Mongolia is Hauterivian to Barremian. However, in contrast to the well-established ostracod biostratigraphic framework in southeastern, eastern, and central Mongolia, there is limited research on the ostracod taxonomy in the Gobi–Altai area thus far, limiting its potential for biostratigraphic application.
Here we identify nine species of six genera from two sections of the Shinekhudag Formation in the Gobi–Altai region. The ostracod fauna from the Tsagaan Tsuvarga Section consists of three species of three genera. Among them, Yumenia oriformis was reported from the Lower Huihuipu Series (Hou, Reference Hou1958), which is equal to the Xiagou Formation of China and known as early Aptian in age (Xi et al., Reference Xi, Wan, Li and Li2019; Li et al., Reference Li, Wang and Zhang2020). Scabriculocypris subscalara was found in the Barremian Nantun Formation of the Hailaer Basin, China (Ye et al., Reference Ye, Huang, Liu, Zhang and Chen2003; Xi et al., Reference Xi, Wan, Li and Li2019). Vlakomia ulanense was found in the Andaikhudag Formation in the Altai region (Neustrueva et al., Reference Neustrueva, Sinitsa, Khand and Melnikova2005) (which stratigraphically equals the Shinekhudag Formation; see Khand et al., Reference Khand, Badamgarav, Ariunchimeg, Barsbold, Okada and Mateer2000). In the Oshih Hollow East Section, six species of three genera are identified. Among them, Cypridea ihsienensis was reported from the Aptian–Albian Fuxin Formation (Hou, Reference Hou1958; Wang et al., Reference Wang, Sha, Pan and Zhang2015). Cypridea unicostata is widely distributed in Hauterivian to Albian strata in China and Mongolia (Sinitsa, Reference Sinitsa1993; Neustrueva et al., Reference Neustrueva, Sinitsa, Khand and Melnikova2005; Wang et al., Reference Wang, Sha, Pan and Zhang2015). Cypridea verrucata is distributed in the Barremian to Aptian strata of Mongolia (Neustrueva, Reference Neustrueva, Kramarenko, Luvsandansan and Rozhdestvensky1974; Neustrueva et al., Reference Neustrueva, Sinitsa, Khand and Melnikova2005; this study). Species of Candona and Trapezoidella are commonly reported from the Lower Cretaceous Strata of East Asia (Hou et al., Reference Hou, Gou and Chen2002; Neustrueva et al., Reference Neustrueva, Sinitsa, Khand and Melnikova2005; Wang et al., Reference Wang, Sha, Pan and Zhang2015, Reference Wang, Sha, Pan and Zuo2016; Choi et al., Reference Choi, Jugdernamjil, Huh and Khand2017, Reference Choi, Wang, Hu and Huh2020, Reference Choi, Jia, Huh, Jung and Wang2021). Consequently, we assign a Barremian–Aptian age for the Shinekhudag Formation in Gobi–Altai area (Fig. 7).

Figure 7. Stratigraphic ranges of the ostracod species in this study from the Shinekhudag Formation.
The high ratio of nonendemic ostracod species from the Shinekhudag Formation provides the evidence of the microfaunal exchanges and paleobiogeographic relationships between the Cretaceous basins of East Asian regions. Three species (Cypridea ihsienensis, C. unicostata, and Scabriculocypris subscalara) have been reported from the Lower Cretaceous deposits in northeast China, such as the Liaoning area and the Hailaer Basin (Hou, Reference Hou1958; Ye et al., Reference Ye, Huang, Liu, Zhang and Chen2003). Yumenia oriformis has been recorded from northwest China (Hou, Reference Hou1958). Cypridea tumefacta is a member of the C. laevigata/setina group, and the representatives of this species group have been reported from the pre-Aptian strata (mostly Berriasian to Valanginian in Europe) of Eurasia (see species list in Schudack and Schudack, Reference Schudack and Schudack2009). The post-Aptian members were reported from Europe (Portugal) and North Africa (Tunisia) (Cabral, Reference Cabral, Crasquin-Soleau, Braccini and Lethiers1998; Trabelsi et al., Reference Trabelsi, Colin and Touir2011). Thus, the members of this species group in Asia probably derived from immigrated representatives of the pre-Aptian European lineage, but many scenarios are possible. Therefore, it should be considered that origin and evolutionary trends in Asian species include C. tumefacta. Therefore, the ostracod fauna of the Shinekhudag Formation records relatively frequent faunal exchanges between southwest Mongolia and northeast China during the Early Cretaceous. However, it seems that there are only a few, single faunal relationships with other regions (i.e., northwest China).
Paleoecology
In Mongolia, the paleoecology of Early Cretaceous ostracod faunas including species of the Shinekhudag Formation has not been studied thus far. Therefore, we here present first insights into paleoenvironmental interpretations of the Early Cretaceous nonmarine ostracod fauna in Mongolia. The taphonomic analyses are very limited or impossible since all specimens in this study had to be picked from the surfaces of the sedimentary rock samples (see “Materials and methods”). Because the studied sections (Tsagaan Tsuvarga and Oshih Hollow East) of the Shinekhudag Formation in this study are separated, the ostracod fauna can be divided into two assemblages, which show very different species composition and diversity. The first paleoenvironmental interpretations of each section are as follows.
Samples of the Tsagaan Tsuvarga Section are characterized by low diversity (three species) and the dominance of Vlakomia ulanense. Despite the small quantity of ostracod specimens available, the assemblage with various ontogenetic stages (A-3 to adult) of Vlakomia ulanense in the samples (4UK070311-3, 4UK070311-2, 4UK070311-1, and 4UK070411-2) is considered an in situ assemblage. The lack of V. ulanense in the sample 4UK070411-1 may indicate that the paleoenvironment of that time interval was unfavorable for this taxon, but the specific reason is uncertain and needs to be investigated (or Vlakomia specimens are just not discovered).
The genus Vlakomia is considered to have a wide range of paleoecological tolerance (Choi et al., Reference Choi, Wang, Hu and Huh2020, p. 21) because their extant and fossil relatives of the subfamily Limnocytherinae of the family Limnocytheridae occur in both ephemeral and permanent water bodies, including fresh to saline water environments, and they are able to lay desiccation-resistant eggs (Horne and Martens, Reference Horne and Martens1998; Meisch, Reference Meisch, Schwoerbel and Zwick2000; Karanovic, Reference Karanovic2012; Santos Filho et al., Reference Santos Filho, Fauth, Sames, Wolfgring and Villegas-Martin2021). In recent studies (Wang and Choi, Reference Wang and Choi2019; Choi et al., Reference Choi, Wang, Hu and Huh2020), representatives of the genus Vlakomia are interpreted to have tolerated high salinities in nonmarine water bodies (e.g., saline lakes) and in nonmarine to transitional environments with the influence of seawater. The two species of Yumenia and Scabriculocypris in the Tsagaan Tsuvarga Section are assigned to the superfamily Cypridoidea (family Alloiocyprideidae). Almost all cypridoidean taxa (both living and fossil representatives) also certainly possess(-ed) desiccation-resistant eggs, which enable them to dwell in ephemeral water bodies and survive desiccation during dry periods (Horne and Martens, Reference Horne and Martens1998; Horne, Reference Horne2002; Sames, Reference Sames and Sames2011b; Sames and Horne, Reference Sames and Horne2012). Thus, the absence of permanent water-body indicators, such as the representatives of the subfamily Timiriaseviinae (family Limnocytheridae, superfamily Cytheroidea) and the superfamily Darwinuloidea (Horne and Martens, Reference Horne and Martens1998; Horne, Reference Horne2002; Sames, Reference Sames and Sames2011a, Reference Sames and Samesb), could be explained by an increased aridity in the study area (Horne, Reference Horne2002). In addition, the low diversity and high salinity tolerance of Vlakomia species may indicate an increased salinity. This kind of setting is comparable to the nonmarine saline to transitional settings with seawater influence of the Albian Tongfosi Formation of northeast China, where species of Vlakomia and Scabriculocypris dominate the ostracod assemblages (Wang and Choi, Reference Wang and Choi2019; Choi et al., Reference Choi, Wang, Hu and Huh2020). The saline conditions in the Tsagaan Tsuvarga area of the Shinekhudag Formation are supported by the presence of carbonates, gypsum, and microbial–caddisfly bioherms in a shallow-littoral setting and have been interpreted as relatively alkaline, saline to hypersaline, and rich in Ca/Mg and bicarbonate ions in a paleolake (Adiya et al., Reference Adiya, Johnson, Loewen, Ritterbush, Constenius and Dinter2017). Because the areas of Central Asia, including southwest Mongolia, were far from the sea during the Cretaceous, nonmarine ostracod taxa and other proxies in the Tsagaan Tsuvarga Section are indicative of the existence of an athalassic (salinity of inland waters not deriving from seawater) saline lake setting (see Horne, Reference Horne2002). Therefore, we confirm that the saline lake conditions (possibly hypersaline, see Adiya et al., Reference Adiya, Johnson, Loewen, Ritterbush, Constenius and Dinter2017, p. 27) under arid climate have persisted during the deposition periods of the sampling layers in the Tsagaan Tsuvarga Section.
The samples (4TM062411-1, 5MT062111-9, 5MT062111-3, and 5KC062111-2) of the Oshih Hollow East Section (Fig. 2) yielded a cypridoidean ostracod fauna of relatively high diversity (six species), including the genera Cypridea, Candona, and ?Trapezoidella. A common feature of these samples is lack of younger juveniles (under A-1 instar), which may suggest that smaller juveniles have been removed from the assemblage by postmortem current and deposited in much deeper areas of water bodies (Whatley, Reference Whatley and Maddocks1983, Reference Whatley, De Decker, Colin and Peypouquet1988; Boomer et al., Reference Boomer, Horne, Slipper, Park and Smith2003; Nye et al., Reference Nye, Feist-Burkhardt, Horne, Ross and Whittaker2008). However, due to the small number of ostracod specimens in the samples, the younger juveniles may have been overlooked. Thus, it is very difficult to interpret the taphonomic processes of the Oshih Hollow East Section.
The species composition of this section is characterized by only representatives of superfamily Cypridoidea. Especially, four species in this study belong to the genus Cypridea (extinct family Cyprideidae). The representatives of the genus Cypridea are the most common Late Mesozoic ostracod faunal elements of nonmarine settings worldwide (except Antarctica and Australia; Horne and Martens, Reference Horne and Martens1998; Sames, Reference Sames and Sames2011b). As many other cypridoideans, the genus Cypridea is adapted to both temporary and permanent water bodies due to their desiccation-resistant eggs and reproductive strategies (sexual or mixed reproduction; see Horne and Martens, Reference Horne and Martens1998; Sames, Reference Sames and Sames2011b), and therefore, the dominance of Cypridea species in deposits with a lack of Timiriaseviinae and darwinuloidean ostracods has been considered indicative of ephemeral water bodies or unstable lake environments (see Horne, Reference Horne2002; Wang et al., Reference Wang, Sha and Pan2013; Trabelsi et al., Reference Trabelsi, Sames, Salmouna, Piovesan, Ben Rouina, Houla, Touir and Soussi2015; Choi et al., Reference Choi, Wang, Hu and Huh2020, Reference Choi, Jia, Huh, Jung and Wang2021; Santos Filho et al., Reference Santos Filho, Fauth, Sames, Wolfgring and Villegas-Martin2021, and references therein). In addition, some of them possibly inhabited saline lakes, but mainly lived in freshwater environments (Horne, Reference Horne2002; Sames, Reference Sames and Sames2011b). Extant representatives of Candona are living in various aquatic environments, such as lakes/ponds and springs (Löffler and Danielopol, Reference Löffler, Danielopol and Illies1978; Carbonel et al., Reference Carbonel, Colin, Danielopol, Löffler and Neustrueva1988; Meisch, Reference Meisch, Schwoerbel and Zwick2000; Karanovic, Reference Karanovic2012). Trapezoidella also belongs to superfamily Cypridoidea, and the representatives of the genus have been discovered from the typical Cretaceous nonmarine deposits in East Asia (e.g., Lübimova, Reference Lübimova1956; Khand et al., Reference Khand, Sames and Schudack2007). Consequently, the dominance of cypridoideans and the lack of cytheroidean/darwinuloidean ostracods of the Oshih Hollow East Section are indicative of unstable and ephemeral water bodies (e.g., ephemeral lakes, ponds, and seasonal pools).
In summary, two studied sections of the Shinekhudag Formation show very different ostracod diversity and paleoecological information; we interpret that the ostracod-bearing samples of the Tsagaan Tsuvarga Section have been deposited in a shallow-littoral zone of an athalassic saline lake and its ephemeral marginal areas, which might be influenced by chemical effects and fluctuation of paleolake under arid climate. The paleoenvironment of the Oshih Hollow East Section is interpreted to represent ephemeral water bodies.
Conclusions
A total of nine species belonging to six genera of an ostracod fauna were recovered from the Lower Cretaceous lacustrine sediments from two sections (Tsagaan Tsuvarga and Oshih Hollow East) of the Shinekhudag Formation, southwest Mongolia (Gobi–Altai area): Cypridea verrucata, C. ihsienensis, C. unicostata, C. tumefacta, Yumenia cf. Y. oriformis, Scabriculocypris subscalara, ?Trapezoidella sp., Candona sp., and Vlakomia ulanense.
This ostracod fauna suggests that the age of the Shinekhudag Formation is Barremian–Aptian, and there is evidence of active faunal exchange and migration between northeast China and the Gobi–Altai area of Mongolia, and potentially several other areas (e.g., northwest China). Among the taxa, Vlakomia ulanense is considered an ancestor group of the Far East group, such as species in the Tongfosi Formation (Albian), China.
Two different faunal compositions in the sections provide new information about paleoenvironment and paleoecology of ostracod fauna of the Shinekhudag Formation. In particular, the low diversity and paleoecological tolerance of Vlakomia in the Tsagaan Tsuvarga Section are interpreted to indicate a shallow-littoral zone in the athalassic saline lake (possibly hypersaline conditions) under arid climate. By contrast, the dominance of cypridoideans (especially the genus Cypridea) in the Oshih Hollow East Section indicates ephemeral water bodies. The absence of the subfamily Timiriaseviinae (family Limnocytheridae, superfamily Cytheroidea) and superfamily Darwinuloidea is also indicative of an arid climate in the Tsagaan Tsuvarga and Oshih Hollow East areas of the Shinekhudag Formation.
Our results suggest that the application of ostracod fauna in the Shinekhudag Formation can be a very useful tool for an interpretation of the Lower Cretaceous deposits of southwest Mongolia. Therefore, this investigation provides a new insight and better understanding for the biostratigraphy, paleobiogeography, paleoenvironment, and resource exploration in the Gobi–Altai area of Mongolia.
Acknowledgments
We thank Y. Fang (NIGPAS), H. Zhang (NIGPAS), and Z. Ge (CDUT) for their technical assistance. Many thanks go to J. Sha (NIGPAS) for providing rock samples. We are very grateful to A. de Jesus e Sousa and an anonymous reviewer for their constructive suggestions and comments. This study was supported by the National Natural Science Foundation of China (grant no. 42288201, 41873018), Youth Innovation Promotion Association, CAS (2019309) (YQW), “Fossil excavation and research for the Natural History exhibition” of Daegu National Science Museum, South Korea, and the National Research Foundation of Korea (NRF), funded by the Korean government (Ministry of Science and ICT) (grant no. 2022M3H9A1097213) (B.-D.C.).
Declaration of competing interests
The authors declare that they have no conflicts of interest.








