


Introduction
From an ecological perspective, dispersal affects species distribution and abundance, population dynamics and persistence, and community structure (Dieckmann et al. Reference Dieckmann, O′Hara and Weisser1999). Within a biogeographical context, dispersal is not only important for geographical spread, but also seems to have genetic consequences (e.g. inbreeding reduction and gene exchange between populations; Janes Reference Janes and Cody1985). Often during the dispersal period, floater individuals of some long-lived raptors, such as Bonelli’s Eagle Hieraaetus fasciatus and Golden Eagle Aquila chrysaetos, tend to stay in settlement areas outside of breeding territories where they remain for a variable period before joining breeding populations (Ferrer Reference Ferrer1993, Mañosa et al. Reference Mañosa, Real and Codina1998). The dispersal period is relatively long and critical for these raptors (Newton Reference Newton1979, Watson Reference Watson1997, Mañosa et al. Reference Mañosa, Real and Codina1998), as it accounts for approximately 80% of floater mortality among large raptor species in Mediterranean areas (Real et al. Reference Real, Grande, Mañosa and Sánchez-Zapata2001, Díaz Reference Díaz2004, Carrete et al. Reference Carrete, Sánchez-Zapata, Calvo and Lande2005). The factors affecting survival during the non-breeding stage have major consequences for population stability in these species (Arroyo Reference Arroyo, Madroño, González and Atienza2004, Ontiveros et al. Reference Ontiveros, Real, Balbontin, Carrete, Ferrero, Ferrer, Mañosa, Pleguezuelos and Sánchez-Zapata2004, Margalida et al. Reference Moreno, Villafuerte and Delibes2008, Soutullo et al. Reference Soutullo, López-López and Urios2008a).
Bonelli’s Eagle and Golden Eagle are long-lived birds of prey that nest mainly on cliffs and have a modal clutch size of two eggs (range 1–3), with young maturing at about 4–5 years (Cramp and Simmons Reference Cramp and Simmons1980). These raptors could coexist in settlement areas in the southern Iberian Peninsula (Ferrer Reference Ferrer1993), where the lack of appropriate settlement areas or reduction in habitat quality may decrease floater survival and seriously threaten population stability (Mañosa et al. Reference Mañosa, Real and Codina1998, Arroyo Reference Arroyo, Madroño, González and Atienza2004, Ontiveros et al. Reference Ontiveros, Real, Balbontin, Carrete, Ferrero, Ferrer, Mañosa, Pleguezuelos and Sánchez-Zapata2004). In recent decades, Bonelli’s and Golden Eagles have shown a marked decline throughout most of their distribution and thus are currently considered endangered species in Western Europe (BirdLife International/European Bird Census Council 2000, Arroyo Reference Arroyo, Madroño, González and Atienza2004, Real Reference Real, Madroño, González and Atienza2004). Declines have been attributed to habitat change, direct persecution, electrocution by electric power lines, decrease in prey abundance, and disturbance (Watson Reference Watson1997, Arroyo Reference Arroyo, Madroño, González and Atienza2004, Ontiveros et al. Reference Ontiveros, Real, Balbontin, Carrete, Ferrero, Ferrer, Mañosa, Pleguezuelos and Sánchez-Zapata2004). The development of effective conservation programmes for threatened species such as these requires a clear understanding of the factors determining their distribution and abundance as well as their ecological requirements (Soulé and Wilcox Reference Soulé and Wilcox1980).
Until now, most studies on the conservation of raptors have been focussed on their breeding biology, habitat selection, diet composition, survival, and interspecific interactions (e.g. Real et al. Reference Real, Grande, Mañosa and Sánchez-Zapata2001, Borgo Reference Borgo2003, Penteriani et al. Reference Penteriani, Balbontin and Ferrer2003, Ontiveros et al. Reference Ontiveros, Real, Balbontin, Carrete, Ferrero, Ferrer, Mañosa, Pleguezuelos and Sánchez-Zapata2004, Reference Ontiveros, Pleguezuelos and Caro2005, Carrete et al. Reference Carrete, Sánchez-Zapata, Calvo and Lande2005, McIntyre et al. Reference McIntyre, Steenhof, Kochert and Collopy2006, Jenkins et al. in press, Rollan et al. in press). However, detailed reports on habitat selection by these raptor species during the dispersal phase are scarce (Arroyo Reference Arroyo, Madroño, González and Atienza2004, Real Reference Real, Madroño, González and Atienza2004) and as a consequence, this life stage cannot be adequately considered in management strategies. Studies have shown that floaters of Bonelli’s and Golden Eagles may cover large areas during the dispersal phase (Cadahia et al. 2005, Soutullo et al. Reference Soutullo, Urios, Ferrer and Peñarrubia2006), becoming established in settlement areas with high densities of their main prey (Mañosa et al. Reference Mañosa, Real and Codina1998, Balbontín Reference Balbontín2005, Soutullo et al. Reference Soutullo, Urios, Ferrer and López-López2008b).
Here we provide data on some of the most important settlement areas of Bonelli’s Eagle and Golden Eagle in the southern Iberian Peninsula, characterising the habitat selection of dispersing eagles, and we examine the possible sympatry of the two eagles in these settlement areas. The results may help to identify potential settlement areas for floater eagles of both species elsewhere, thus having implications for management strategies for the two species.
Methods
Study area
The study area was located in southern Spain, in two different administrative regions, Andalucía and Murcia, where Bonelli’s and Golden Eagle populations are the most healthy and well known (Gil-Sánchez et al. Reference Gil-Sánchez, Moleón, Otero and Bautista2004, Ontiveros et al. Reference Ontiveros, Pleguezuelos and Caro2005, Carrete et al. Reference Carrete, Sánchez-Zapata, Tella, Gil-Sánchez and Moleón2006), with 294–333 pairs of Bonelli’s Eagle and 256–290 pairs of Golden Eagle (Arroyo Reference Arroyo, Madroño, González and Atienza2004, Real Reference Real, Madroño, González and Atienza2004). The area is largely mountainous, and the climate is typically Mediterranean, with annual temperatures averaging 15.6–19.5 °C and annual rainfall averaging 300–790 mm (CMA 1997, Carrete et al. Reference Carrete, Sánchez-Zapata, Martínez, Sánchez and Calvo2002). The field work covered 12 areas (see below and Figure 1) where natural vegetation consisted of shrubs, grasslands, and mixed forest of Quercus rotundifolia and Pinus spp., but human activity (notably farming) over the millennia has transformed the landscape into a mosaic. These areas suffer much anthropogenic disturbance and are not protected, which may lead to increasing mortality risk for floaters.

Figure 1. Study area showing settlement (solid circles) and non-settlement (empty circles) areas for Bonelli’s and Golden Eagles in Southern Spain. Numbers refer to the number of 9-km2 squares sampled in each area. For details, see the Methods section.
Raptor survey
Based on our field experience during the last 18 years and the literature (Real Reference Arroyo, Madroño, González and Atienza2004, Balbontín Reference Balbontín2005), we located six settlement areas in the study area. For comparative purposes, we selected six non-settlement areas in the region among those that matched the following characteristics: located outside breeding territories (at least 5 km from closest nest), far away from urban areas (at least 5 km from the central point), below 1,700 m asl (maximum altitude of hunting areas for Bonelli’s Eagle, and for most of the Golden Eagles in the study area), and with no record of floater eagles prior to this study (from the authors’ data). We surveyed the 12 different areas during 2001–2006. Firstly, sampling was performed in autumn and winter, seasons when maximum density can be attained for floater eagles in settlement areas (Mañosa et al. Reference Mañosa, Real and Codina1998, authors’ unpubl. data), to confirm presence/absence of floaters in settlement/non-settlement areas, respectively. Afterwards, areas with floater presence were sampled on a monthly basis while non-settlement areas were sampled more irregularly.
Eagle presence and density were determined by driving along line transects, at a slow speed (20 km hr−1), in as straight a line as possible, on days of good visibility, in the morning, with two people (one trained observer and a driver). This method has repeatedly proved adequate to gather information on the distribution and relative abundance of raptors (Mañosa et al. Reference Mañosa, Real and Codina1998, Sánchez-Zapata and Calvo Reference Sánchez-Zapata and Calvo1999; see discussion by Carrete et al. Reference Carrete, Tella, Blanco and Bertellotti2009). In total, 75 transects were sampled in the six settlement areas, and 20 in the six non-settlement areas, with a total of 1,543 km of line transects covered. The number of kilometres driven and number of individuals of each raptor species were recorded, and eagles were classified as juvenile (1 year), immature, (2–3 years for Bonelli’s Eagle and 2–4 years for Golden Eagle), subadult (4 years for Bonelli’s Eagle and 5 years for Golden Eagle), and adult (> 4 years for Bonelli’s Eagle and > 5 years for Golden Eagle), according to plumage criteria (Parellada Reference Parellada1984, Watson Reference Watson1997, Forsman Reference Forsman1999). Relative abundance of floater eagles in each survey was measured as the average number of birds detected per 100 km.
Habitat selection variables
Each area (both settlement and non-settlement) was divided into 3 × 3 km squares (UTM grid system), and according to the extent of the rather homogeneous landscape in the area, between two and four squares were selected. In total, we selected 29 squares, 16 with presence (for the six settlement areas) and 13 with absence of Bonelli’s and Golden Eagles (for the six non-settlement areas; Figure 1).
Landscape characteristics for each 3 × 3 km square in settlement and non-settlement areas were analysed by means of a Geographic Information System (GIS). Squares 3 × 3 km (900 ha) constitute an appropriate scale for habitat-selection studies in raptors (Mañosa et al. Reference Mañosa, Real and Codina1998, Sánchez-Zapata and Calvo Reference Sánchez-Zapata and Calvo1999, Balbontín Reference Balbontín2005, López-López et al. Reference López-López, García-Ripollés, Aguilar, García-López and Verdejo2006). We measured different variables (Table 1) and related them to the presence or absence of floater eagles in the dispersal phase. Topography, climate, and human population density variables were taken from Corine Land Cover (1:25,000; CEC 1991). Land use variables (habitat heterogeneity, percentage of orchards, pastureland, scrubland, and forest) were taken from soil-use maps of the regions of Andalucía and Murcia (CMA 1999, CAAMA 2000). The number of kilometres of paved and unpaved roads and the distance to the nearest nest of Bonelli’s or Golden Eagle were measured from 1:50,000 digital military maps using Arc View GIS 3.2. We controlled for spatial autocorrelation by including as covariates the geographic variables longitude (Long) and latitude (Lat) of the centre of the squares, as well as the terms Long2, Long3, Lat2, Lat3, Long2 × Lat, and Long × Lat2, according to Legendre (Reference Legendre1993). To compare the presence of the two eagle species, we also used abundance of Bonelli’s and Golden Eagles in breeding territories, computed as the frequency of UTM 10 × 10 km squares with eagle presence into UTM 50 × 50 km squares (Martí and Del Moral Reference Martí and Del Moral2003). Any significant change in land use was noted in the study area during the study period.
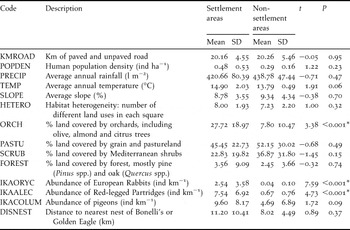
Table 1. Variables used (except geographical ones), their values, and comparison (Student’s t- test) between settlement and non-settlement areas of large diurnal raptors (Bonelli’s and Golden Eagles) in southern Spain. Sampling unit was 3 × 3 km square (UTM grid system). More information in the Methods section. Asterisks indicate significance after Bonferroni sequential correction.
Furthermore, we recorded the abundance of the main prey of the Bonelli’s and Golden Eagles in each 3 × 3 km square. In the study area, European Rabbits Oryctolagus cuniculus, Red-legged Partridges Alectoris rufa and pigeons (Columba palumbus and C. livia) account for 79.2% and 61.9%, respectively, of the dietary biomass of these eagles (Delibes et al. Reference Delibes, Calderón and Hiraldo1975, Gil-Sánchez et al. Reference Gil-Sánchez, Molino and Valenzuela1994, Ontiveros et al. Reference Ontiveros, Pleguezuelos and Caro2005). Line transects were used to provide an index of prey density in each square in the settlement and non-settlement areas. This method has proven effective in determining prey abundance for raptors and in comparing prey densities among different zones (Fitzner et al. Reference Fitzner, Rogers and Uresk1977), being less difficult to perform than absolute-density methods and equally useful (Caughley Reference Caughley1977). Prey density was measured as the mean number of individuals per kilometre of census. On average, 4 km of census per year were carried out in each square, stratified according to the surface area of the habitats (Caughley Reference Caughley1977), for two consecutive years (2005 and 2006). Censuses were made by one observer on foot, 06h00–09h30, on days of good visibility, at a speed of approx. 2 km h−1, during the same period (February–April) for all areas. We did not undertake censuses before February so as to avoid the hunting season, and after April because of demographic explosions of rabbits in the southern Iberian Peninsula (Soriguer Reference Soriguer1981). The line transect is the most accurate method for estimating rabbit abundance (Palomares Reference Palomares2001), and diurnal censuses proved to be useful for this species (Soriguer et al. Reference Soriguer, Pérez and Fandos1997, Serrano Reference Serrano1998, Palomares et al. Reference Palomares, Delibes, Revilla, Calzada and Fedriani2001), since rabbits, although primarily nocturnal, also show substantial diurnal activity (Soriguer and Rogers Reference Soriguer and Rogers1981, Moreno et al. Reference Moreno, Villafuerte and Delibes1996); thus, we deemed the diurnal rabbit census to be a more realistic estimate of prey density for strictly diurnal predators such as Bonelli’s and Golden Eagles (Ontiveros et al. Reference Ontiveros, Pleguezuelos and Caro2005).
Statistical procedures
Non-parametric variables were log-transformed (Sokal and Rohlf Reference Sokal and Rohlf1995), and we performed the statistical analysis in three steps:
i) We used the Student’s t-test to identify differences in the variables between settlement and non-settlement squares for floater eagles, and with Bonelli’s Eagle presence vs. Golden Eagle presence, and included squares with presence of both species once in each group. A circular statistic was used for analyses of orientation in settlement areas (Rayleigh’s test), since a simple arithmetic mean of recorded angles is inadequate (Fisher Reference Fisher1995).
ii) We used a General Discriminant Analysis (GDA) to identify the variables that explained differences between areas with presence or absence of floater eagles. These analyses allowed models to be generated by stepwise selection of predictors as well as by the selections of the best-subset of predictors. We chose the latter approach, which allows model uncertainty to be measured at the same time as parameter uncertainty, to assess the likely bias in parameters resulting from selections. We used Statistica 7.0 software, which may generate multivariate linear models, and ranks the set of all possible models by their Wilks’s lambda values (an estimate of the unexplained variance), separately within each model order (k = 1, 2, … 14), where a model’s order is defined as the number of predictor variables it includes. We retained the five best subset models from orders 1, 2, …13, and also the single model containing all 14 predictors, for a total of 66 candidate models. Smaller values indicate a better, more parsimonious model and denote strong group separation (Quinn and Keough Reference Quinn and Keough2003). From all possible models generated in the analysis, we chose those explaining the highest percentage of presence-absence of the eagles with fewer variables. Because in our analysis the sample size could not be increased to three times the number of variables measured, a jack-knife classification was carried out for the analysis (Willians and Titus Reference Willians and Titus1988).
iii) We used logistic regression (LR) (Siegel and Castellan Reference Siegel and Castellan1988, Jongman et al. Reference Jongman, Ter Braak and van Tongeren1995), to explore their coincidence with previous results from the GDA. Logistic regression is often applied to ornithological data for predicting presence or absence of species (Fielding and Bell Reference Fielding and Bell1997, Manel et al. Reference Manel, Dias, Buckton and Ormerod1999). To eliminate variables without significant effects on the presence of the eagles, we performed a forward stepwise model.
Results
In the settlement areas, we recorded 41 observations of Bonelli’s Eagles and 29 of Golden Eagles, with 2.27 ± 2.98 eagles 100 km−1 (mean ± standard deviation) and 2.31 ± 1.89 eagles 100 km−1 for Bonelli’s Eagle and Golden Eagle, respectively. The age ratio of the eagles was 73.17% and 58.62% juvenile, 17.07% and 31.04% immature, and 9.76% and 10.34% subadult individuals of Bonelli’s and Golden Eagles, respectively, as we did not observe any adult eagles during the censuses. No significant interspecific difference was found in the abundance index in settlement areas (Mann-Whitney U-test, Z = -0.322, P = 0.74, n = 6) and the age frequency (Kolmogorov-Smirnov test, Dmax = 0.25, P > 0.1, n = 4).
Overall, 16 3 × 3 km squares in six different settlement areas and 13 3 × 3 km squares in six non-settlement areas (Figure 1) were used to compare settlement preferences for floater Bonelli’s and/or Golden Eagles. In those settlement areas, we observed the presence of only Bonelli’s Eagle in four 3 × 3 km squares and only Golden Eagle presence in seven squares, while both eagles were observed in the remaining five squares. Squares with floater eagles differed significantly from those without floater eagles in terms of abundance of European Rabbits, Red-legged Partridges, and presence of orchards, which were all greater in settlement areas (Table 2). These differences remained significant after Bonferroni correction. No significant interannual difference was found in any prey density in any of the areas surveyed (Kruskal-Wallis test: H1,8 < 1.33, P > 0.25, in all cases). The mean orientation and angular deviation of slopes in squares with presence and absence of floater eagles were 237.96° ± 54.43° and 345.99° ± 54.99°, respectively. Squares with the presence of floater eagles revealed a trend towards a south-western orientation (Raleigh test = 0.35, P = 0.007).
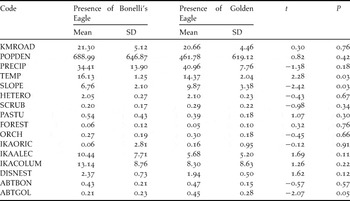
Table 2. Mean, standard deviation (SD), and comparison (Student’s t-test) for variables in settlement areas with presence of Bonelli’s and Golden Eagles in southern Spain.
In the GDA differentiating settlement and non-settlement areas, 66 best-subset models were generated. Wilks’s lambda values consistently stabilised after four variables. The differential score between the lowest Wilks’s lambda and the best model with four variables was only 0.062. The abundance of European Rabbits and Red-legged Partridges, the percentage of orchards, and the habitat heterogeneity, were considered the best predictors of presence or absence of floater eagles (Wilks’s lambda = 0.162, F(4,24) = 30.996, P < 0.001). The best models were (see Table 1 for the meaning of the variable acronyms):
Square with eagle presence=−10.15 − 0.97 IKAORYC + 0.27 IKAALEC + 0.01 ORCH + 2.28 HETERO
Square with eagle absence=−22.72 − 3.15 IKAORYC − 0.49 IKAALEC − 0.19 ORCH + 3.44 HETERO
With these equations, 100% of the squares with presence or absence of floater eagles were correctly classified. A jack-knife classification reduced the correct classification of presence to 99.3% and of absence to 95.9%.
The best LR model for the identification of settlement areas coincided with the GDA in three out of the four variables, also setting abundance of European Rabbit, Red-legged Partridge, and percentage of orchards, as the most parsimonious predictors (χ23=39.645, P<0.001). The best model was:
This model also classified correctly 100% of the squares, thus the inclusion of other variables did not improve the final model.
The best model of GDA and LR did not include any spatial variable, suggesting that there was no effect of spatial autocorrelation. Squares with Bonelli’s Eagle exhibited higher average annual temperature and slope than those with the Golden Eagle (Table 2), although the differences became non-significant after Bonferroni correction.
Discussion
All statistical analyses showed that settlement areas selected by floater eagles had a greater presence of their main prey and orchards than non-settlement areas; the best model selected by GDA also included habitat heterogeneity, defined as the number of different land use types in each square (see Atauri and de Lucio Reference Atauri and de Lucio2001). Furthermore, discriminant and logistic regression analysis coincided in identifying these variables to explain almost all the variance of the settlement areas, despite variability in landscapes throughout the wide geographic range of the study area. In the study area, orchards consisted mainly of olive trees, and several studies have shown the importance of this traditional landscape as a habitat for rabbits and partridges (Rogers et al. Reference Rogers, Arthur, Soriguer, Thomson and King1994, Vargas et al. Reference Vargas, Guerrero, Farfán, Barbosa and Real2006). Higher habitat heterogeneity also favours the presence of these game species, and landscapes with these characteristics can be considered good foraging habitats for raptors (Sánchez-Zapata and Calvo Reference Sánchez-Zapata and Calvo1999, Fortuna Reference Fortuna2002, Sergio et al. Reference Sergio, Marchesi and Pedrini2004). The statistical analysis of circular data also showed that settlement areas tended towards a south-western orientation. In the morning in the study area, the air warms up and ascends (thermal bubbles) on south-facing slopes, frequently used by raptors to save energy during gliding flight (Janes Reference Janes and Cody1985, Ontiveros Reference Ontiveros1999), especially Bonelli’s Eagle, which has poor flying lift in flat areas (Parellada et al. Reference Parellada, De Juan and Alamany1984). Similarly, Balbontín (Reference Balbontín2005) also found sunny orientations to be selected in the settlement areas of Bonelli’s Eagle in south-western Spain.
Two studies have analysed local settlement areas for floater eagles in the Iberian Peninsula, although only for Bonelli’s Eagle (Mañosa et al. Reference Mañosa, Real and Codina1998, Balbontín Reference Balbontín2005). Coinciding with Mañosa et al. (Reference Mañosa, Real and Codina1998), we found in the present study that settlement areas for both eagle species were selected mainly for food availability rather than for topography or landscape patterns. Contrary to the present study and Mañosa et al. (Reference Mañosa, Real and Codina1998), Balbontín (Reference Balbontín2005) found that the habitat selection of floater Bonelli’s Eagle was related to topography, land use, and human disturbance. However, although this author did not consider prey abundance, the land use variables in his study that were selected by eagles (scrub and pasture) were directly related to a higher presence and/or detectability of the main prey species, such as partridges and rabbits (Fortuna Reference Fortuna2002, Ontiveros et al. Reference Ontiveros, Pleguezuelos and Caro2005). Soutullo et al. (Reference Soutullo, Urios, Ferrer and López-López2008b) found that juvenile Golden Eagles used a wide range of habitats in eastern Spain, at least when yearlings (1–2 years old), though showed some preference for certain types of habitat, such as coniferous forest, sclerophyllous vegetation, and mosaic agricultural landscapes. As in our results, settlement areas of Spanish Imperial Eagle Aquila adalberti in southern Spain were characterised by high rabbit densities and the presence of pasture, farmlands and dehesas (Ferrer Reference Ferrer1993, Ferrer and Harte Reference Ferrer and Harte1997).
In relation to density, Mañosa et al. (Reference Mañosa, Real and Codina1998), reported an encounter rate of 2.7 eagles 100-km−1 in one settlement area of Bonelli’s Eagle in north-eastern Spain, similar to the value found in the present study as an average of six different settlement areas. Likewise, the percentages of juvenile, immature, and subadult individuals were similar to those found in our study.
Our results showed no significant interspecific differences in abundance and age frequency between the two eagle species, rather there was high habitat overlap between species in settlement areas. Only lower mean annual temperatures and the steepest slopes appear to account for the higher presence of Golden Eagle over Bonelli’s Eagle in settlement areas. The segregation mediated by environmental temperature has also been found in breeding populations, related to differences in their niche for that trait (López-López et al. Reference López-López, García-Ripollés, Aguilar, García-López and Verdejo2004, Moreno-Rueda et al. Reference Moreno-Rueda, Pizarro, Ontiveros and Pleguezuelos2009), because Bonelli’s Eagle is more thermophilic than Golden Eagle (Muñoz et al. Reference Muñoz, Real, Barbosa and Vargas2005, López-López et al. Reference López-López, García-Ripollés, Soutullo, Cadahía and Urios2007). It is remarkable that two large raptor species with similar ecological requirements coincided to some degree in the settlement areas considered here, also exhibiting similar values of abundance and age-frequency densities. This coincides with the recent perspective on breeding populations of both eagle species, which suggest that long-term coexistence is likely (López-López et al. Reference López-López, Soutullo, García-Ripollés, Urios, Cadahía and Ferrer2008), intraspecific competition being more important than interspecific for neighbouring pairs (Carrete et al. Reference Carrete, Sánchez-Zapata, Calvo and Lande2005; but see Gil-Sánchez et al. Reference Gil-Sánchez, Moleón, Otero and Bautista2004).
The southern Iberian Peninsula harbours large populations of Bonelli’s and Golden Eagles (Arroyo Reference Arroyo, Madroño, González and Atienza2004, Real Reference Real, Madroño, González and Atienza2004), and both species are mainly cliff-nesting raptors that occupy the most rugged areas. However, settlement areas lack adequate nesting cliffs and are closely surrounded by breeding territories. In settlement areas, floaters wait for the occurrence of breeding opportunities within the reproductive fraction of the population. Probably, young eagles select settlements areas under two cues, areas with sufficient food availability in which they can avoid competition with breeders. However, the major threats for both eagles in such areas include prey decrease, habitat change, and persecution, both direct (shooting) and indirect (poisoning) on private hunting estates (Arroyo Reference Arroyo, Madroño, González and Atienza2004, Real Reference Real, Madroño, González and Atienza2004). At least for Bonelli’s Eagle, floater mortality plays a key role in determining the overall population trend in Iberian Peninsula (Soutullo et al. Reference Soutullo, López-López and Urios2008a), and the same applies to other threatened raptors, such as Spanish Imperial Eagle (Penteriani et al. Reference Penteriani, Otalora, Sergio and Ferrer2005, Reference Penteriani, Otalora and Ferrer2008).
Management implications
Because both Bonelli’s and Golden Eagles spend a significant portion of their life (2–4 years) within settlement areas, the location and protection of such areas for these eagles would lengthen their lifespan by protecting the habitat during the floater stage (Balbontín Reference Balbontín2005). Because settlement areas are unknown or difficult to detect, fewer efforts have so far been devoted to the conservation of these sites than to protecting breeding territories (Penteriani et al. Reference Penteriani, Otalora, Sergio and Ferrer2005). However, management action for the conservation of these species should also be focused at minimising floater mortality in settlement areas (Ontiveros et al. Reference Ontiveros, Real, Balbontin, Carrete, Ferrero, Ferrer, Mañosa, Pleguezuelos and Sánchez-Zapata2004, Real Reference Real, Madroño, González and Atienza2004, Soutullo et al. Reference Soutullo, López-López and Urios2008a). Moreover, in Mediterranean Spain, the protection of such settlement areas would benefit both large raptors of conservation concern.
Due to the high importance of rabbit and partridge abundance in settlement areas, the management of these prey species and their habitats is crucial. These game species sustain a large number of natural predators in the Iberian Peninsula (Valkama et al. Reference Valkama, Korpimäki, Arroyo, Beja, Bretagnolle, Bro, Kenward, Mañosa, Redpath, Thirgood and Viñuela2005, Delibes-Mateos et al. Reference Delibes-Mateos, Redpath, Angulo, Ferreras and Villafuerte2007), but are also important for hunting by humans on private hunting estates (Lucio Reference Lucio, Fuentes, Sánchez and Pajuelo1991, Villafuerte et al. Reference Villafuerte, Viñuela and Blanco1998). All the settlement areas considered in this study are included in hunting estates. For these reasons, vigilance of hunting quotas on these estates is necessary for the maintenance of appropriate prey-species abundance. In this way, authorities could establish agreements with hunters, such as economic compensation for not hunting in some years, or through the purchase and management of these areas. Furthermore, agri-environmental measures that encourage low-intensity farming systems should be implemented in these areas. Finally, the conservation of mosaic habitats should be the basis for increasing land-cover diversity and thereby favouring the abundance of the main prey species of floater eagles in the settlement areas.
Acknowledgements
We thank Pedro Gutierrez for his contribution to the field work. Gregorio Moreno provided helpful suggestions on a previous draft, José Poquet helped with the statistical analyses, and David Nesbitt improved the English. Most of this study was performed without financial support.



