



Introduction
The genus Psophia (hereafter, trumpeters) comprises eight species endemic to the Amazon and Guiana regions of northern South America (Ribas et al. Reference Ribas, Aleixo, Nogueira, Miyaki and Cracraft2012, Pacheco et al. Reference Pacheco, Silveira, Aleixo, Agne, Bencke, Bravo, Brito, Cohn-Haft, Maurício, Naka, Olmos, Posso, Lees and Figueiredo2021). Trumpeters are large-bodied (> 1 kg), terrestrial, group-living and highly territorial (Sherman Reference Sherman1995b, Sherman and Eason Reference Sherman and Eason1998). They are predominantly frugivorous (Erard et al. Reference Erard, Thery and Sabatier1991, Reference Erard, Théry and Sabatier2007, Winkler et al. Reference Winkler, Billerman, Lovette, Billerman, Keeney, Rodewald and Schulenberg2020) and play an important role in forest dynamics as they swallow entire fruits and disseminate their intact seeds (Erard et al. Reference Erard, Théry and Sabatier2007).
Trumpeters are forest-interior specialists and thus highly sensitive to disturbance (Parry et al. Reference Parry, Barlow and Peres2007, Moura et al. Reference Moura, Lees, Aleixo, Barlow, Dantas, Ferreira, Lima and Gardner2014, Michalski and Peres Reference Michalski and Peres2017, Benchimol and Peres Reference Benchimol and Peres2021). They tend to be rare or absent from disturbed and secondary forests (Parry et al. Reference Parry, Barlow and Peres2007), and they often disappear from all but the largest fragments in fragmented forest landscapes (Moura et al. Reference Moura, Lees, Aleixo, Barlow, Dantas, Ferreira, Lima and Gardner2014, Michalski and Peres Reference Michalski and Peres2017, Benchimol and Peres Reference Benchimol and Peres2021). Indeed, they share most of the ecological attributes that make a bird species sensitive to forest disturbance, including large body size, slow breeding rate, and a dependency on forest habitat and its fruit and invertebrate food resources (Newbold et al. Reference Newbold, Scharlemann, Butchart, Şekercioǧlu, Alkemade, Booth and Purves2013). In addition, their large body size, conspicuousness and group-living habits makes them vulnerable to hunters (Thiollay Reference Thiollay2005). Consequently, three trumpeter species are currently categorized as threatened and two as ‘Near Threatened’ on the IUCN Red List of Threatened Species (IUCN 2021).
The Black-winged Trumpeter (Psophia obscura; Fig. S3 in the online supplementary material) is endemic to the Belém Centre of Endemism in extreme eastern Amazonia (Oppenheimer and Silveira Reference Oppenheimer and Silveira2009, Ribas et al. Reference Ribas, Aleixo, Nogueira, Miyaki and Cracraft2012). The Belém Centre of Endemism coincides with the oldest and most deforested human occupation frontier of the biome and its most threatened region (Moura et al. Reference Moura, Lees, Aleixo, Barlow, Dantas, Ferreira, Lima and Gardner2014, Vedovato et al. Reference Vedovato, Fonseca, Arai, Anderson and Aragão2016, Celentano et al. Reference Celentano, Rousseau, Muniz, Varga, Martinez, Carneiro, Miranda, Barros, Freitas, Narvaes, Adami, Gomes, Rodrigues and Martins2017, Silva Junior et al. Reference Silva Junior, Celentano, Rousseau, de Moura, van D. Varga, Martinez and Martins2020). Due to continued habitat loss and its vulnerability to hunting, the Black-winged Trumpeter is suspected to be declining with possibly <250 individuals left in the wild. It is therefore categorized as ‘Critically Endangered’ by both IUCN (IUCN criteria A and C; BirdLife International 2018) and the Brazilian Red List of threatened species (IUCN criterion A; Ministério do Meio Ambiente - MMA 2014, Silveira Reference Silveira2018). However, there are no population size estimates for the species, so its categorization, based on the suspected number of mature individuals (criterion C) is questionable.
Despite its threatened status, little is known about the species apart from occasional records in bird inventories throughout its range (Portes et al. Reference Portes, Carneiro, Schunck, de Sousa e Silva, Zimmer, Whittaker, Poletto, Silveira and Aleixo2011, Lees et al. Reference Lees, de Moura, Santana, Aleixo, Barlow, Berenguer, Ferreira and Gardner2012, Lima and Raices Reference Lima and Raices2012, Moura et al. Reference Moura, Lees, Aleixo, Barlow, Dantas, Ferreira, Lima and Gardner2014). Current distribution is limited to some of the largest and most undisturbed fragments of the Belém Centre of Endemism (Portes et al. Reference Portes, Carneiro, Schunck, de Sousa e Silva, Zimmer, Whittaker, Poletto, Silveira and Aleixo2011, Moura et al. Reference Moura, Lees, Aleixo, Barlow, Dantas, Ferreira, Lima and Gardner2014) and most of the remaining individuals belong to one of two disjunct populations: a western population at unprotected forest remnants in the municipality of Paragominas and an eastern population at Gurupi Biological Reserve and contiguous Indigenous Lands (Lees et al. Reference Lees, de Moura, Santana, Aleixo, Barlow, Berenguer, Ferreira and Gardner2012, Lima and Raices Reference Lima and Raices2012, BirdLife International 2018).
In this study, we investigate site occupancy dynamics of a Black-winged Trumpeter population at Gurupi Biological Reserve, a key stronghold for conservation of the species. More specifically, we use data from five years of camera trap monitoring and a dynamic occupancy modelling approach to investigate potential factors affecting occupancy rates and trends in the reserve as a function of apparent survival and colonization rates.
Methods
Study area
Gurupi Biological Reserve is a 270,000-ha protected area located in extreme eastern Amazonia (Fig. 1). Together with contiguous Indigenous Lands, the reserve comprises the last remaining block of continuous Amazonian forests in the Belém Centre of Endemism (Silva Junior et al. Reference Silva Junior, Celentano, Rousseau, de Moura, van D. Varga, Martinez and Martins2020), and is one of the two most important strongholds for the Black-winged Trumpeter (Lima et al. Reference Lima, Martínez and Raíces2014, BirdLife International 2018). The reserve has a tropical monsoonal climate with mean annual temperatures >26 Co and mean annual rainfall of 1,800 mm (Alvares et al. Reference Alvares, Stape, Sentelhas, Gonçalves and Sparovek2013). The terrain is flat to undulating with elevation ranging from 50 to 340 m above sea level. The reserve was entirely covered by evergreen tropical forest, but has lost about 30% of its forest cover to illegal deforestation in the last decades (Celentano et al. Reference Celentano, Rousseau, Muniz, Varga, Martinez, Carneiro, Miranda, Barros, Freitas, Narvaes, Adami, Gomes, Rodrigues and Martins2017). Much of its remaining forests is degraded by illegal selective logging and fires (Celentano et al. Reference Celentano, Rousseau, Muniz, Varga, Martinez, Carneiro, Miranda, Barros, Freitas, Narvaes, Adami, Gomes, Rodrigues and Martins2017, Paiva et al. Reference Paiva, Ruivo, da Silva Júnior, Maciel, Braga, de Andrade, dos Santos Junior, da Rocha, de Freitas, Leite, Gama, Santos, da Silva, Silva and Ferreira2020). Still, it safeguards a significant portion of the regional biodiversity, including the full complement of medium- to large-sized terrestrial vertebrates (Lopes and Ferrari Reference Lopes and Ferrari2000, Lima et al. Reference Lima, Martínez and Raíces2014, Carvalho et al. Reference Carvalho, Mendonça, Martins and Haugaasen2020).

Figure 1. Map of Gurupi Biological Reserve, showing the distribution of camera trap stations. The inset map shows the location of the study area in Northeastern Brazil.
Camera trapping
Camera trap surveys were conducted between 2016 and 2020 as part of the Brazilian in situ monitoring program of Federal Protected Areas (Programa Monitora). Sampling followed the Tropical Ecology Assessment and Monitoring (TEAM) protocol for vertebrates (Rovero and Ahumada Reference Rovero and Ahumada2017): during every dry season (August to November), we deployed camera-traps (model Bushnell Trophy Cam) at 61 permanent sampling sites distributed in two regular arrays with a density of one sampling site per 2 km2 (Fig. 1). Cameras were attached to trees at knee height, perpendicular to the ground and facing either north or south to avoid direct sunlight at sunrise and sunset, and the vegetation directly in front of cameras was cleared. Cameras were set to operate continuously for at least 30 days per year. Images were processed in the wild.ID software (Fegraus and MacCarthy Reference Fegraus, MacCarthy, Rovero and Zimmermann2016). We assumed a 60 min interval for independence between detection events at the same sampling site. Although the same sites were sampled in all years, the number of operational cameras varied between years due to occasional camera malfunctions (Table. S1).
Occupancy predictors
We quantified seven site-level variables to represent environmental and anthropogenic factors that may plausibly affect Black-winged Trumpeter occupancy and detection rates: (1) Site elevation (mean = 141.7 m, range: 77–270 m) was extracted from the ALOS global digital surface model provided by the Japan Aerospace Exploration Agency (JAXA) (Tadono et al. Reference Tadono, Ishida, Oda, Naito, Minakawa and Iwamoto2014). (2) Distance to water (mean = 1.2 km, range: 0.1–3.8) is the shortest distance between sampling sites and their nearest stream. (3) Distance to edge (mean = 2 km, range: 0.2–5) is the shortest distance between sampling locations and the nearest forest edge, estimated using the 30 m resolution land cover classification of the MapBiomas monitoring system for 2016 (Souza et al. Reference Souza, Shimbo, Rosa, Parente, Alencar, Rudorff, Hasenack, Matsumoto, Ferreira, Souza-Filho, de Oliveira, Rocha, Fonseca and Marques2020). (4) Site-level tree density (mean = 798 trees/ha, range: 523–1569) and (5) basal area (mean = 27.4 m2/ha, range: 8.2–45) were estimated using the point-centred quarter method (Cottam and Curtis Reference Cottam and Curtis1956). Starting from each camera location, we ran three 50-m transects in the direction of 0, 120 and 240 magnetic degrees. Along each transect, we established five sampling points at 10-m intervals. The area around each point was divided into four quarters and the diameter at breast height (dbh) of the nearest tree with dbh ≥ 10 cm at each quarter was recorded. Tree density was estimated using the equation:
 $ D=1/{\overline{r}}^2 $
, where
$ D=1/{\overline{r}}^2 $
, where
 $ \overline{r} $
is the mean point-to-tree distance across all quarters (Cottam and Curtis Reference Cottam and Curtis1956). Basal area was estimated using the equation
$ \overline{r} $
is the mean point-to-tree distance across all quarters (Cottam and Curtis Reference Cottam and Curtis1956). Basal area was estimated using the equation
 $ BA= meanBA\cdot D $
, where
$ BA= meanBA\cdot D $
, where
 $ meanBA $
is the mean basal area of sampled trees across all quarters (Cottam and Curtis Reference Cottam and Curtis1956), with basal area of individual trees given by the equation
$ meanBA $
is the mean basal area of sampled trees across all quarters (Cottam and Curtis Reference Cottam and Curtis1956), with basal area of individual trees given by the equation
 $ BA=\pi \cdot {\left( dbh/2\right)}^2 $
. To minimize the weight of a few exceptionally large trees, basal area of trees in the top 2.5% quantile (n = 9 trees) were replaced by the quantile threshold value. Finally, we quantified two variables representing site-level impacts of past illegal logging: (6) recovery time (mean = 13, range: 6–21) as the number of years elapsed since any portion of a buffer of 500 m around each sampling site was logged for the last time, and (7) logging bouts (mean = 2, range: 1–5) as the number of different years in which each 500 m buffer zone was logged. To recover the history of illegal logging, we used visual interpretation of 1984–2016 Landsat time series data (Carvalho et al. Reference Carvalho, Mendonça, Martins and Haugaasen2020). Evidence of logging, such as roads, log decks and large canopy gaps are detectable in Landsat imagery for one to several years after logging (Stone and Lefebvre Reference Stone and Lefebvre1998, Matricardi et al. Reference Matricardi, Skole, Cochrane, Pedlowski and Chomentowski2007, Asner et al. Reference Asner, Rudel, Aide, Defries and Emerson2009). We used the USGS Earth Explorer interface (http://earthexplorer.usgs.gov) to download Landsat images from every year of the series and selected, whenever possible, cloudless images from dry season months. We then recorded the presence or absence of logging signs within 500-m buffers centred on each camera trap site and for each year in the series. By this procedure we estimated both recovery time and the number of logging bouts. For unlogged sites, we set maximum recovery time as 33 years, corresponding to the start of the time series. More details are provided in Carvalho et al. (Reference Carvalho, Mendonça, Martins and Haugaasen2020).
$ BA=\pi \cdot {\left( dbh/2\right)}^2 $
. To minimize the weight of a few exceptionally large trees, basal area of trees in the top 2.5% quantile (n = 9 trees) were replaced by the quantile threshold value. Finally, we quantified two variables representing site-level impacts of past illegal logging: (6) recovery time (mean = 13, range: 6–21) as the number of years elapsed since any portion of a buffer of 500 m around each sampling site was logged for the last time, and (7) logging bouts (mean = 2, range: 1–5) as the number of different years in which each 500 m buffer zone was logged. To recover the history of illegal logging, we used visual interpretation of 1984–2016 Landsat time series data (Carvalho et al. Reference Carvalho, Mendonça, Martins and Haugaasen2020). Evidence of logging, such as roads, log decks and large canopy gaps are detectable in Landsat imagery for one to several years after logging (Stone and Lefebvre Reference Stone and Lefebvre1998, Matricardi et al. Reference Matricardi, Skole, Cochrane, Pedlowski and Chomentowski2007, Asner et al. Reference Asner, Rudel, Aide, Defries and Emerson2009). We used the USGS Earth Explorer interface (http://earthexplorer.usgs.gov) to download Landsat images from every year of the series and selected, whenever possible, cloudless images from dry season months. We then recorded the presence or absence of logging signs within 500-m buffers centred on each camera trap site and for each year in the series. By this procedure we estimated both recovery time and the number of logging bouts. For unlogged sites, we set maximum recovery time as 33 years, corresponding to the start of the time series. More details are provided in Carvalho et al. (Reference Carvalho, Mendonça, Martins and Haugaasen2020).
All variables were standardized before the analysis. Distance to edges was log-transformed before the analysis. We used Pearson coefficients (r) to test for collinearity among predictors, retaining for analysis only one variable from any pair with high (|r| > 0.60) correlation. Thus, distance to water and logging bouts were removed from analysis as they were correlated with elevation and recovery time respectively.
Data analyses
We used multi-season occupancy modelling (MacKenzie et al. Reference MacKenzie, Nichols, Hines, Knutson and Franklin2003) to investigate Black-winged Trumpeter occupancy trends. This approach uses detection/non-detection data to estimate occupancy rates (the proportion of sites occupied by the species) and to model temporal changes in occupancy as a function of local survival and colonization processes, while accounting for imperfect detection (Royle and Dorazio Reference Royle and Dorazio2008, Kéry and Schaub Reference Kéry and Schaub2012, MacKenzie et al. Reference MacKenzie, Nichols, Royle, Pollock, Bailey and Hines2017). The model requires sampling at two temporal scales, namely primary and secondary periods. Occupancy at any given sampling site may change between primary periods, but not between secondary periods that are nested within primary periods (MacKenzie et al. Reference MacKenzie, Nichols, Hines, Knutson and Franklin2003). In our analysis, primary periods corresponded to years and secondary periods to six-day sampling occasions. To meet the assumption of population closure within primary periods, we only use data from the first 30 days of sampling for any site and year. To increase detection probabilities and facilitate model convergence in data analysis, we collapsed data into six-day sampling occasions.
We modelled occurrence of the species at site i in year k (zi,k ) as a Bernoulli outcome governed by occupancy probability at site i in year k (ψi,k ):
 $$ {z}_{i,k}\sim Bern\left({\psi}_{i,k}\right) $$
$$ {z}_{i,k}\sim Bern\left({\psi}_{i,k}\right) $$
We modelled observations, consisting of detection/non-detection of the species at site i, sampling occasion j and year k (yi,j,k ) as Bernoulli outcomes governed by the product of zi,k and detection probability at site i, sampling occasion j and year k (pi,j,k ):
 $$ {y}_{i,j,k}\sim Bern({z}_{i,k}\cdot {p}_{i,j,k}) $$
$$ {y}_{i,j,k}\sim Bern({z}_{i,k}\cdot {p}_{i,j,k}) $$
We used a logit link function to model detection probability as a function of random site and year effects, while assuming constant detection within the same site and year:
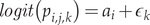 $$ logit({p}_{i,j,k})={a}_i+{\epsilon}_k $$
$$ logit({p}_{i,j,k})={a}_i+{\epsilon}_k $$
We used a logit link function to model initial occupancy (year k=1) as a function of random site effects, elevation, distance to edge, basal area, tree density, and recovery time:
 $$ logit\left({\psi}_{i,1}\right)={\displaystyle \begin{array}{l}{a}_i+{b}_1\cdot {elevation}_i+{b}_2\cdot {distEdge}_i+{b}_3\cdot {basalArea}_i+{b}_4\cdot {treeDensity}_i+{b}_5\\ {}\cdot \hskip0.35em {recovery}_i\end{array}} $$
$$ logit\left({\psi}_{i,1}\right)={\displaystyle \begin{array}{l}{a}_i+{b}_1\cdot {elevation}_i+{b}_2\cdot {distEdge}_i+{b}_3\cdot {basalArea}_i+{b}_4\cdot {treeDensity}_i+{b}_5\\ {}\cdot \hskip0.35em {recovery}_i\end{array}} $$
We modelled occupancy in subsequent years as a function of year-specific survival (φ) and colonization (γ) rates, estimated from the data:
 $$ {\psi}_{i,k+1}={\psi}_{i,k}\cdot {\phi}_k+\left(1-{\psi}_{i,k}\right)\cdot {\gamma}_k $$
$$ {\psi}_{i,k+1}={\psi}_{i,k}\cdot {\phi}_k+\left(1-{\psi}_{i,k}\right)\cdot {\gamma}_k $$
To assess whether year-to-year changes in occupancy were significant, we estimated the derived parameter growth rate (λ) as follows (Royle and Dorazio Reference Royle and Dorazio2008):
 $$ \lambda =\frac{\psi_{k+1}}{\psi_k} $$
$$ \lambda =\frac{\psi_{k+1}}{\psi_k} $$
We fitted the model in a Bayesian framework, adapting the specifications provided by (Kéry and Schaub Reference Kéry and Schaub2012). We implemented the model in JAGS (Plummer Reference Plummer2015) using the R2jags package (Su and Yajima Reference Su and Yajima2012). We used non-informative priors for all parameters and ran three chains with 250,000 Markov Chain Monte Carlo (MCMC) iterations, with a burn-in of 100,000 and a thinning rate of 150. We evaluated parameter convergence using the Gelman-Rubin diagnostic (Gelman and Shirley Reference Gelman, Shirley, Brooks, Gelman, Jones and Meng2011). We considered that there was support for a covariate effect when the 95% posterior credible interval (CI) for the parameter did not include zero. We considered that there was evidence for significant change in occupancy between a given year k and k+1 if the posterior credible interval of λ did not overlap 1 (Ahumada et al. Reference Ahumada, Hurtado and Lizcano2013). Data and R codes for analysis are available at https://github.com/ICMBio-CENAP/Psophia-obscura.
Results
A total effort of 8,674 camera-trap days across five years of sampling yielded 2,876 photos of Black-winged Trumpeter, corresponding to 181 independent detection events (figures refer to data not yet collapsed into six-day occasions; Table S1). In any given year, the species was recorded at 14–22 sites, corresponding to naïve (i.e., uncorrected for imperfect detection) occupancy rates of 0.24–0.39. Table 1 presents posterior summaries for selected model parameters.
Table 1. Posterior means, standard deviation and 95% credible intervals of selected model parameters. Occupancy (ψ) and detection (p) rates are estimated by site and year, such that the values shown in the table are averaged across sites. Survival (φ), colonization (γ) and growth (λ) rates are estimated on an annual basis for all sites combined.

Average initial (i.e. for 2016) occupancy probability was 0.57 and average initial detection was 0.14. Initial occupancy probability was negatively related to elevation and tree density, and positively related to recovery time (Table 2, Fig. 2, Fig. S1). Basal area was positively but not significantly related to occupancy, while distance to edges had no relationship with occupancy (Table 2, Fig. S1).
Table 2. Posterior estimates for the effects of covariates on Black-winged Trumpeter occupancy probabilities.


Figure 2. Predicted effect of model covariates on Black-winged Trumpeter initial occupancy probabilities at Gurupi Biological Reserve: (A) elevation (masl); (B) tree density (stems/ha); (C) post-logging recovery time (years). Solid black line represent posterior mean and light grey lines the uncertainty in estimates, based on a random posterior sample of 200 iterations.
The dynamic occupancy model reveals fluctuating but stable occupancy rates across years, with average annual ψ fluctuating between 0.57 and 0.67 (Table 1, Fig. 3) and always consistently higher than naïve occupancy rates. Detection rates were low and varied little across years, ranging between 0.09 and 0.15 (Table 1). Between-year growth rates (λ) ranged between 0.92 and 1.2 over years and were never significantly different from unity (Table 1, Fig. S2), which is consistent with the observed stability in occupancy rates. Occupancy dynamics was characterized by high between-year survival probabilities, with φ ranging between 0.72 and 0.81 across years, while between-year colonization probabilities were considerably lower, with γ ranging between 0.36 and 0.47 (Table 1).

Figure 3. Temporal dynamics in Black-winged Trumpeter occupancy probabilities at Gurupi Biological Reserve, 2016-2020. The solid black line represents the posterior mean, and light grey lines the uncertainty in the estimate, based on a random posterior sample of 200 iterations.
Discussion
This study provides the first assessment of factors affecting the distribution and trends of the Black-winged Trumpeter in one of its key conservation areas. Black-winged Trumpeter initial occupancy rate was affected by elevation, tree density and recovery time, a set of variables that represent natural or anthropogenic factors, or a combination of both, while occupancy rates remained stable throughout the study.
The effect of elevation was negative. This is a key determinant of biodiversity distribution in tropical forests as it is correlated to a range of biotic and abiotic variables including water availability, soil and vegetation characteristics, and microclimate (de Castilho et al. Reference de Castilho, Magnusson, de Araújo, Luizão, Luizão, Lima and Higuchi2006, de Toledo et al. Reference de Toledo, Magnusson, Castilho and Nascimento2011, Norris et al. Reference Norris, Fortin and Magnusson2014). The negative effect of elevation on trumpeter occupancy may relate to higher humidity and productivity in lower-lying areas; trumpeters prefer the moistest parts of the forest for foraging (Erard et al. Reference Erard, Théry and Sabatier2007) and tend to be more abundant at sites closer to water (Michalski et al. Reference Michalski, Norris, De Oliveira and Michalski2015, Paredes et al. Reference Paredes, Norris, De Oliveira and Michalski2017, Mere Roncal et al. Reference Mere Roncal, Middendorf, Forsyth, Cáceres, Blake, Almeyda Zambrano and Broadbent2019, del Hoyo et al. Reference del Hoyo, Sherman, Kirwan, Collar, Christie, Sharpe, Billerman, Keeney, Rodewald and Schulenberg2020). Such habitat specificity probably increases the species’ vulnerability to climate change. Habitat suitability models predict that it might lose more than 70% of suitable areas due to climate change by 2050 (de Moraes et al. Reference de Moraes, Santos, Gonçalves, de Oliveira, Gomes and Lima2020).
The results confirm that the species is an undisturbed primary forest specialist (Portes et al. Reference Portes, Carneiro, Schunck, de Sousa e Silva, Zimmer, Whittaker, Poletto, Silveira and Aleixo2011, Moura et al. Reference Moura, Lees, Andretti, Davis, Solar, Aleixo, Barlow, Ferreira and Gardner2013, Lima et al. Reference Lima, Martínez and Raíces2014, del Hoyo et al. Reference del Hoyo, Sherman, Kirwan, Collar, Christie, Sharpe, Billerman, Keeney, Rodewald and Schulenberg2020), even though it can tolerate low levels of disturbance (Lima and Raices Reference Lima and Raices2012). The two variables representing forest structure were strongly related to Black-winged Trumpeter occupancy probability, with tree density having a significantly negative effect and basal area having a positive, albeit non-significant, effect. Both variables are indicators of forest successional stage, with basal area increasing (Lu et al. Reference Lu, Mausel, Brondízio and Moran2003, Myster Reference Myster2016, Caron et al. Reference Caron, Chuma, Sandi and Norris2021) and tree density decreasing (Wright Reference Wright2005) as forests mature. Such a preference for undisturbed forests seems to be universal for the genus (Parry et al. Reference Parry, Barlow and Peres2007, Michalski et al. Reference Michalski, Norris, De Oliveira and Michalski2015, Michalski and Peres Reference Michalski and Peres2017).
The precise mechanism by which forest structure affects the species is unclear and we can only speculate. Perhaps early successional or disturbed forests lack essential resources. For example, disturbed areas have fewer large trees, which provide the bulk of fruit consumed by trumpeters (Erard et al. Reference Erard, Théry and Sabatier2007), and may lack trees with appropriate cavities for nesting (Sherman Reference Sherman1995a, Cornelius et al. Reference Cornelius, Cockle, Politi, Berkunsky, Sandoval, Ojeda, Rivera, Hunter and Martin2008). Dense understorey in disturbed areas may also hinder communication and vigilance. This is detrimental to trumpeters, as they require constant acoustic and visual contact between group members to coordinate their movements and watch for predators (Seddon et al. Reference Seddon, Alvarez and Tobias2002). Whatever the mechanism, the species prefers areas with a structure like mature forests.
Since most logging in the area took place over a decade ago, the positive effect of recovery time implies that logging effects are persistent. The removal of large trees and the increase in understorey density due to logging gaps have similar effects to those discussed above. Even though some parameters such as canopy cover and microclimate recover relatively quickly after logging (Senior et al. Reference Senior, Hill, Benedick and Edwards2018, Mollinari et al. Reference Mollinari, Peres and Edwards2019), recovery of forest biomass (Gatti et al. Reference Gatti, Castaldi, Lindsell, Coomes, Marchetti, Maesano, Paola, Paparella, Valentini, Cazzolla, Simona, Lindsell, Coomes and Cazzolla Gatti2014), floristic composition (Gaui et al. Reference Gaui, Costa, Coelho de Souza, Amaral, de Carvalho, Reis and Higuchi2019) and availability of large trees (Pinho et al. Reference Pinho, Peres, Leal and Tabarelli2020) may take much longer. This is particularly prominent in illegally logged areas, which undergo more severe damage in their structure and composition (Pacheco et al. Reference Pacheco, Cerutti, Edwards, Lescuyer, Mejía, Navarro, Obidzinski, Pokorny, Sist, Kleinschmit, Mansourian, Wildburger and Purret2016). Overall, trumpeter responses to logging seem to depend on the amount of damage to the residual forests. Previous studies found variable responses from trumpeters, from negative responses to conventional logging that persist for over a decade (Thiollay Reference Thiollay1997) to slightly positive responses to reduced-impact logging in the short-term (Bicknell and Peres Reference Bicknell and Peres2010). A previous study using a subset of the same data used here did not find a significant response to logging by the Black-winged Trumpeter, although the direction of the response was consistent with what is reported here (Carvalho et al. Reference Carvalho, Mendonça, Martins and Haugaasen2020).
The dynamic model revealed fluctuating but stable occupancy rates across years. Given that trumpeters are highly territorial with relatively inflexible home range boundaries (Sherman and Eason Reference Sherman and Eason1998) and that camera spacing was large enough to ensure there was no more than one camera per territory (assuming territories of approximately 70 ha; Sherman and Eason Reference Sherman and Eason1998), this suggests that the number of occupied territories, and possibly the population, has remained constant throughout the study. This finding underlines the importance of the Gurupi Biological Reserve as a core site for the conservation of the species, whose decline is inferred mostly from the continued loss of habitat (BirdLife International 2018, Silveira Reference Silveira2018), as there are no monitoring data available.
Stable occupancy rates can be interpreted as evidence of territorial saturation, where all suitable sites are occupied, and no vacant territories are available for expansion. Territorial saturation is probably the rule among trumpeters, and has been suggested as a candidate driver for the evolution of cooperative breeding in the genus (Sherman Reference Sherman1995b). This is because trumpeter territory sizes are largely defined by food availability during the lean period (Sherman Reference Sherman1995b, Sherman and Eason Reference Sherman and Eason1998), and this limits the number of territories that can fit within a given area. Consistent with this view is the fact that population dynamics was shaped primarily by high survival rates and not colonization, as revealed by the consistently higher estimates for φ compared to γ. In fact, this was expected as adult trumpeters have high survival rates (Sherman Reference Sherman1995b) and groups can persist even longer than individuals, leading to continuous occupation of territories by the same group on a multi-year basis.
This view does not rule out the possibility of changes in the occupancy rate in the future. Suitable areas may become vacant if their resident groups collapse and are not replaced. The amount of suitable habitat may decrease if the reserve undergoes additional logging, forest fires or deforestation. The opposite is also possible as the amount of suitable habitat may increase as previously degraded areas recover from disturbance. Continuous monitoring is needed to reveal future trends, while additional studies may indicate the maximum attainable population size in the reserve and what factors may be limiting population growth and expansion.
Observed trends should be interpreted with caution, as the study duration was relatively short. Five years is just a little longer than the average tenure of a dominant individual over a group (Sherman Reference Sherman1995b), and approximately equivalent to the estimated generation length of the Black-winged Trumpeter (Bird et al. Reference Bird, Martin, Akçakaya, Gilroy, Burfield, Garnett, Symes, Taylor, Sekercioglu and Butchart2020). For example, the guidelines for using the IUCN Red List categories and criteria require information on population reduction over 10 years or three generations (Rodrigues et al. Reference Rodrigues, Pilgrim, Lamoreux, Hoffmann and Brooks2006), which for the Black-winged Trumpeter corresponds to 15 years (Bird et al. Reference Bird, Martin, Akçakaya, Gilroy, Burfield, Garnett, Symes, Taylor, Sekercioglu and Butchart2020). Therefore, more years of monitoring are needed for robust conclusions on trends for the species.
The apparent stable trumpeter population at Gurupi provides hope for the species within this protected area. However, the conservation prospects for the species remain highly precarious as a large but unknown fraction of its remaining population resides in unprotected forest patches that continue to be lost at alarming rates (Silva Junior et al. Reference Silva Junior, Celentano, Rousseau, de Moura, van D. Varga, Martinez and Martins2020). Furthermore, even protected areas are prone to habitat degradation from logging, fires, and climate change (de Moraes et al. Reference de Moraes, Santos, Gonçalves, de Oliveira, Gomes and Lima2020). We reinforce the main recommendations already provided for the conservation of the species, such as expanding the network of protected areas and improving the management of existing ones (BirdLife International 2018, Silveira Reference Silveira2018). We also recommend continued monitoring at Gurupi to further clarify trends in this key area for the species, as well as additional studies to provide reliable estimates of population size in the reserve and in other areas.
Supplementary Material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0959270922000077
Acknowledgements
We thank the managers of Gurupi Biological Reserve for logistical support. W.M. Silva, M. C. de Lima, F. C. Braga and C. R. D. V. Melo assisted in fieldwork. Ronaldo G. Morato commented on an earlier draft of this manuscript. This work was funded by ARPA – Programa Áreas Protegidas da Amazônia.








 Open access
Open access