


Introduction
Migratory swans and geese have historically played a special role in the spiritual lives of humans in Asia, as the harbingers of the seasons and as a source of food. In contemporary times, knowledge about the abundance and distribution of these migratory waterbirds has become increasingly important for a number of reasons. Firstly, international conventions and organisations, (such as Ramsar Convention, the Convention on Biological Diversity, and IUCN) as well as legislation require such data as indicators of biodiversity and wetland health (e.g. CBD Aichi Conservation Goal C and Target 12; see Butchart Reference Butchart2008). The Ramsar Convention on Wetlands defines that a site regularly supporting 1% or more of a waterbird population qualifies as a wetland of international importance, necessitating regular assessments of what constitutes the total population size in order to define the 1% level (Delany and Scott Reference Delany and Scott2006). This contributes to the identification of the network of protected sites such as those designated under the East Asian-Australasian Flyway Partnership (EAAFP 2015). Secondly, given that many geese are popular quarry species, international conventions (such as the CBD) as well as some domestic regulations require hunting of birds be undertaken in a manner compatible with the concepts of wise and sustainable use that requires monitoring to ensure such exploitation does not threaten the favourable conservation status of a population (Kanstrup Reference Kanstrup, Boere, Galbraith and Stroud2006). Thirdly, many (but by no means all) goose populations in Europe and North America are increasing rapidly (e.g. Fox et al. Reference Fox, Ebbinge, Mitchell, Heinicke, Aarvak, Colhoun, Clausen, Dereliev, Faragó, Koffijberg, Kruckenberg, Loonen, Madsen, Mooij, Musil, Nilsson, Pihl and van der Jeugd2010) and have become increasingly reliant on agriculture for food during the non-breeding season. This may potentially create conflict with farmers when geese exploit forage grasses, cereals and root crop monocultures and/or impact upon the breeding grounds in an adverse manner (e.g. Abraham et al. Reference Abraham, Jefferies and Alisauskas2005, Fox et al. Reference Fox, Madsen, Boyd, Kuijken, Norriss, Tombre and Stroud2005), requiring an understanding of their distribution and abundance to find potential solutions to such conflict. Finally, several populations of swans and geese are thought to be threatened or declining in East Asia (see Wetlands International 2015). Nature conservation programmes designed to restore their populations to more favourable conservation status require monitoring data in order to assess the changing conservation status of a population and ultimately whether their objectives have been met as a result of management actions.
There are three species of Cygnus swans and five species of Anser geese that commonly spend the non-breeding season in East Asia. Knowledge of the abundance and distribution of swans and geese in East Asia is rapidly improving, although information from South Korea and Japan (where organised counts have been undertaken for the last 20 and 40 years, respectively) is substantially better than that in China (where national assessments of the status and distribution have been forthcoming in the last 10 years). In this review, we estimate the population sizes of the Whooper Swan Cygnus cygnus, Tundra Swan C. columbianus, Swan Goose Anser cygnoides, Bean Goose A. fabalis, Greater A. albifrons and Lesser White-fronted Goose A. erythropus based on count information collated for the first time from throughout the non-breeding areas. We compare these with current population estimates and assess their respective conservation status, with particular emphasis on identifying key sites throughout the range. We also discuss the distribution between the core non-breeding areas in South Korea, Japan and China, consider how best to improve collaborative monitoring of these populations in the future through a joint international programme, and consider the conservation threats and challenges to the maintenance of these populations in the immediate future throughout these range states. Unfortunately, it is not currently possible to determine the status of swans and geese in North Korea, despite the fact that we know of the existence of important sites in that country for swans, White-fronted and Bean Geese. In spite of this gap in our knowledge, we still regard this analysis as an important contribution to international cooperation and sincerely hope North Korea will soon be involved in a flyway monitoring and conservation framework.
Methods
Study area and habitat use
In China, non-breeding swans and geese are mainly confined to natural wetlands especially among the many wetlands of the Yangtze River floodplain, where water recession during the non-breeding period creates extensive areas of suitable habitats, but also in smaller numbers along Yellow Sea coasts, across large areas of mudflats, bays and estuaries. Wetlands along the Yangtze River floodplain support most of the swans and geese in eastern China. The exceptions are the (i) Mute Swan Cygnus olor (a relatively rare species in Japan and China, not considered here), (ii) Whooper Swan, which mainly occurs along Yellow Sea coasts (Cao et al. Reference Cao, Zhang, Barter and Lei2010), (iii) Greylag Goose Anser anser which is poorly known and not well monitored in East Asia and (iv) small numbers of the nominate race of Bean Goose Anser fabalis fabalis, which are thought to spend the non-breeding period in Xinjiang Province. Obtaining better knowledge about all of these populations remains an urgent priority for the future but is beyond the scope of this analysis.
In Japan and South Korea, non-breeding swans and geese are mainly associated with agricultural areas. Rice fields comprise 54% and 36% of agricultural land in South Korea and Japan, respectively (Fujioka et al. Reference Fujioka, Don Lee, Kurechi and Yoshida2010). Large amounts of rice grains are left behind in the fields after harvest (Shimada Reference Shimada2003, Stafford et al. Reference Stafford, Kaminski and Reinecke2010), which are exploited by geese, ducks and cranes (although cranes also take macrophytes, plant root material, invertebrates and small mammals). Although rice fields may represent suboptimal habitats for some bird species (Richardson and Taylor Reference Richardson and Taylor2003) compared to natural marshes, many Anatidae tend to depend on rice fields heavily as a source of food (Fujioka et al. Reference Fujioka, Don Lee, Kurechi and Yoshida2010).
Non-breeding season waterbird censuses
South Korea
The Korean Ministry of Environment and its associated institutes, National Institute of Environmental Research (until 2007) and the National Institute of Biological Resources (since 2008), have conducted an annual nationwide census at most lakes, reservoirs, lengths of seashore and bays known to be important for waterbirds, including swans and geese, during the non-breeding period (192 sites as of 2011). The simultaneous two-day field counts are coordinated annually in mid- or late January by ornithologists, avian researchers, experienced birdwatchers and volunteers to estimate the distribution and abundance of waterbirds spending the non-breeding period in South Korea.
Japan
An annual mid-winter non-breeding (c.15 January) survey of waterfowl populations has been conducted since 1970 by the Ministry of the Environment with the assistance of prefectural governments. This nationwide survey covers all of the principal sites for swan, goose and duck species throughout the country during the non-breeding season (Ministry of the Environment of the Government of Japan 2014).
China
Surveys were carried out during mid-November to the end of February in 2002/03–2006/07, concentrating on the Yangtze River and Huai River floodplains, and the coasts of Shandong, Jiangsu, Zhejiang and Fujian Provinces. Incomplete surveys were carried out focusing on key wetlands (Poyang Lake, Dongting Lake and Anhui Lakes) along the Yangtze River floodplain in the non-breeding period of 2008/09 to 2012/13 (Cao Lei, East Dongting and Poyang Lake National Nature Reserve unpubl. data).
Data collection and organization
Total flyway population sizes for each species were estimated for the period 2007–2011 based on best available count data, together with their trends, the latter estimated subject to availability in the three countries (1988–2011 in China, 1996–2011 in Korea and Japan). On the basis of these, key sites in each country or region were selected based on non-breeding census data during 2007–2011 as being those that exceeded the derived 1% criterion for each of the waterbird subpopulations defined by Wetlands International (2015) and see the list in the online Supplementary Material. For Whooper Swan, Tundra Swan, Bean Goose and Lesser White-fronted Geese, 1% criteria were calculated for their entire populations in East Asia according to Wetlands International (2015), while for Greater White-fronted Goose, 1% criteria were calculated separately for the subpopulations in South Korea, Japan and China defined by Wetlands International (2015; Table 4). For Bean Goose, lack of identification to subspecies for many count data meant that separate estimates were not available for middendorffii and serrirostris. For this reason, we simply combined subpopulations of these two subspecies based on regions, i.e. the estimated subpopulations of middendorffii and serrirostris in South Korea, Japan and China were combined to create 1% criteria for these subspecies of Bean Goose in South Korea, Japan and China to the best of our current knowledge (Table 4).
Data used for analysis and mapping were listed as follows:
South Korea: January counts in total for each species during 1999–2011.
Japan: January counts at sites and in total for each species during 1996–2011.
China: Data during 1988–1993 and 1994–1999 were compiled from records held by Waterbird Research in China, the Waterbird Specialist Group of Chinese Ornithological Association, and an array of published and unpublished reports. Data for 2003–2007 were derived from the East China wetlands non-breeding surveys, and during 2008–2011 from the Yangtze River floodplain non-breeding surveys.
Prior assumptions
In deriving the population estimates for each country or region, we have assumed that the abundance and distribution of swans and geese did not vary during the time that they were surveyed in the three countries. In Japan and Korea, surveys were more or less synchronous in mid-January, while in China we cannot fully reject the possibility of missed birds and double counting as the surveys were not done simultaneously. We assume (but cannot fully dismiss) that during the middle of the non-breeding period, there was little exchange of any of these species between China and Japan or Korea. This is supported to some extent for the Greater White-fronted Goose by the existence of profound differences in haplotype frequencies in the mitochondrial DNA of geese sampled in each of the three countries (S. Moriguchi in litt.).
Accuracy of population estimates for South Korea, Japan and East China
Systematic national surveys have been conducted in South Korea and Japan since the 1990s, which gives a high level of confidence in the estimation of annual swan and goose abundance and associated trends there. In China, we have adopted different approaches to compensate for the incompleteness of surveys, which started in the 2000s, to estimate population status. According to studies of waterbird distribution and abundance carried out by Cao et al. (Reference Cao, Zhang, Barter and Lei2010), an assessment of the size of swan and goose populations in the whole of China could be derived almost entirely from counts of the Yangtze River floodplain wetlands during the non-breeding seasons of 2002–2011. This was because the vast majority of these birds were found there during earlier, more complete surveys (Table 1). Yangtze River floodplain surveys are listed according to month and year in Table 2. In years when no complete surveys were carried out, total numbers in the Yangtze River were estimated from counts at key sites in three provinces that were known to hold the majority of these species during the non-breeding season (Table 1). Furthermore, national population sizes were derived for all the species (except for Whooper Swan) based on the numbers in the Yangtze River as a proportion of the total non-breeding numbers in China (see Table 1).
Table 1. The percentage of the total non-breeding populations of the two swan and four goose species in the Yangtze River floodplain (YRF) that were counted in three provinces (Anhui, Jiangxi and Hunan) and those in eastern China that were counted in the YRF.

(Modified from Cao et al. Reference Cao, Barter, Lei and Yang2008b, Reference Cao, Zhang, Barter and Lei2010).
Table 2. Month of counts of the two swan and four goose species conducted in each province along the Yangtze River floodplain in China during 2003–2011.
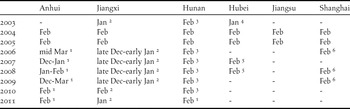
Notes:
1 unpubl. data from L. Cao.
2 unpubl. data from Poyang Lake National Nature Reserve.
3 unpubl. data from East Dongting Lake.
4 Barter and Lei (Reference Barter and Lei2003).
5 Hu et al. (Reference Hu, Liu, Tian, Luo, Li and Chen2008).
6 unpubl. data from Chongming Dongtan National Bird Nature Reserve.
Flyway population sizes estimates
The core distribution range of all swans and geese during the non-breeding season in East Asia lies within mainland China, South Korea and Japan (Wetlands International 2015). As some species may shift their distributions between different years, country counts or estimates for each year may not be independent (i.e. increased numbers in Korea might be attributed to a decrease in numbers in China). Thus, to obtain flyway population estimates for these six species, the counts or estimates from South Korea, Japan and China were summed for each of the years 2007–2011, from which we calculated the five-year total means as population estimates for each species.
Population trends in South Korea, Japan and China
To detect general trends, simple linear regressions were applied to time series data of counts or estimates for each species separately in South Korea, Japan and China.
Species distribution map
Survey data from each site during 2007–2011 were averaged to prepare distribution maps for the six most numerous species for Japan and South Korea. However, for China we do not have such annual data for all species, so data from 2008, 2010 and 2011 were used for the mapping. The maps, prepared from the species databases using ArcGIS 10.0, show the locations of all key wetlands for each species in these three countries. Key wetlands have been identified using the 1% criterion (see above). A table within each map provides information about internationally important concentrations (key sites) located in China, Japan and South Korea.
Results
Population estimates for South Korea, Japan and East China
The total counts from Japan and South Korea and population estimates from China for the six species since the 1990s are listed in Table 3.
Table 3. Population estimates for the two swan and four goose species in South Korea, Japan and eastern China during 1988–2011.

Notes: Months of counts conducted in China are listed in Table 2.
a Unpubl. data from Ministry of the Environment of South Korea, non-breeding birds census conducted at 120 sites of South Korea in January during 1999-2011 in January.
b Unpubl. data from Ministry of the Environment of Japan, January survey conducted nationwide from 1996-2011 in January.
c Estimates during 1988-1993 based on data reported to the Waterbird Specialist Group of the Chinese Ornithological Association (Cong et al. Reference Cong, Cao, Fox, Mark, Rees, Jiang, Ji, Zhu and Song2011, Wang et al. Reference Wang, Fox, Cong, Barter and Cao2012, Zhao et al. Reference Zhao, Cong, Barter, Fox and Cao2012).
d, e and g Estimates during 1994-1999, 2002-2006 and 2007-2011 were based on data reported to the Waterbird Specialist Group of Chinese Ornithological Association, China Bird Reports, China Birdwatch and other online resources (Cong et al. Reference Cong, Cao, Fox, Mark, Rees, Jiang, Ji, Zhu and Song2011, Wang et al. Reference Wang, Fox, Cong, Barter and Cao2012, Zhao et al. Reference Zhao, Cong, Barter, Fox and Cao2012).
f, h, j, l and m Estimates from Cao et al. (Reference Cao, Barter and Lei2008a), Cong et al. (Reference Cong, Cao, Fox, Mark, Rees, Jiang, Ji, Zhu and Song2011), Zhang et al. (Reference Zhang, Cao, Barter, Fox, Zhao, Meng, Shi, Jiang and Zhu2011), Zhao et al. (Reference Zhao, Cong, Barter, Fox and Cao2012) and Wang et al. (Reference Wang, Fox, Cong, Barter and Cao2012).
i Unpubl. data from Cao and Poyang Lake National Nature Reserve and from non-breeding counts at key sites along the Yangtze River floodplain in January and February 2011.
k Estimates from counts during 2004-2011, Cao in press.
Flyway population estimates
The calculated species totals for each of the countries and current estimates of the related flyway populations are given in Table 4. The calculated populations for each of the countries and regions can be directly compared with the current and previous estimates for the flyway populations of each species.
The calculated East Asia populations are lower than the existing flyway estimates for Whooper Swan, Greater White-fronted Goose (in Japan) and Lesser White-fronted Goose. On the other hand, the estimated numbers of Tundra Swan and Greater White-fronted Goose (in China) exceed the previous estimates. Other estimates correspond more or less with those that currently exist.
Table 4. Population estimates for the two swan and four goose species in East Asia based on non-breeding season counts in China, South Korea and Japan with current flyway estimates.
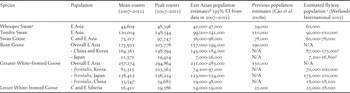
a Missing counts of Whooper Swans from 2008-2010 in China were replaced by mean counts from 2007 and 2011.
b New estimates from this study. Confidence intervals were calculated from 5-year-data collected between 2007-2011 and rounded to the nearest 1,000.
c Current flyway population estimates based on Wetlands International (2015), in which estimates of two Bean Goose subspecies (1 serrirostris and 2 middendorffii) were combined based on distribution of countries or regions.
Population trends for South Korea, Japan and East China
Population trends of each species tended to differ between the three countries during 1988–2011 (Figure 1). In South Korea and Japan, the Greater White-fronted Goose has been increasing significantly (P < 0.01), and Bean Goose has also increased but with a lower rate and level of statistical significance (P < 0.05). Whooper Swans in Korea and Tundra Swans in Japan have also both increased significantly (P < 0.01), while the Tundra Swan has declined significantly in South Korea (P < 0.01) and numbers of Whooper Swans have been stable in Japan. In contrast, non-breeding populations of swans and geese in China have generally shown stable (e.g. Whooper Swan) or fluctuating trends (e.g. Tundra Swan, Swan Goose and Bean Goose) with no significant trends, except for significant declines in abundance of Greater and Lesser White-fronted Geese (P < 0.01).

Figure 1. Population trends of swans and geese in South Korea, Japan and China during 1988–2011.
Over a longer time scale, the trends become clearer. Since the 1970s, swan and goose populations have been stable or increasing over the last three decades in South Korea and Japan (Amano Reference Amano2009, Fujioka et al. Reference Fujioka, Don Lee, Kurechi and Yoshida2010) in contrast to large declines in numbers and contractions in range in China since the 1950s (Cao et al. Reference Cao, Barter and Lei2008a).
Geographical distributions
Maps showing the key sites for the six most numerous species in the three countries are presented in Figure 2. Bean Goose and Greater White-fronted Goose are generally more widely distributed in China, South Korea and Japan. Lesser White-fronted Goose and Swan Goose are highly confined within eastern China, more specifically, to relatively few wetlands along the Yangtze River floodplain. As the Tundra Swan prefers to spend the non-breeding period in habitats experiencing milder climate conditions, relatively few spend the non-breeding period in South Korea (also see Table 2). In contrast, the larger bodied Whooper Swan can tolerate colder conditions and tends to spend the non-breeding period further north throughout East Asia.

Figure 2. Map showing the key sites for the six most numerous species of swans and geese spending the non-breeding period in South Korea, Japan and China.
Discussion
This study represents the first collaborative long-term assessment of overall and country population sizes, trends and current distributions of swans and geese in South Korea, Japan and China, which provide the most important areas for swan and goose species in the East Asian flyway during the non-breeding season. This analysis was essential to establish new 1% criteria to identify key sites throughout this flyway. These data are new and fundamental in their contribution to site safeguard networks and for the long-term effectiveness of conservation actions throughout the flyway. However, these are the first estimates of their kind and we have to draw attention to a number of caveats regarding their potential use now and in the future, which will also serve as pointers to future challenges for potential resolution and improvement in coming years.
Errors in the estimates
As shown in Table 3, waterbirds surveys in China were not synchronous and were incomplete in most non-breeding seasons. It is important to remember this source of potential error in the counts from China when considering the national and flyway estimates. Based on the counts from coastal surveys in February during 2007–2009 (Table 5), only a small proportion (less than 3% of estimates) of the swans and geese concerned in this study spent the non-breeding period along the coast. For this reason, we feel confident in inflating our estimates from the Yangtze River floodplain by such an amount to account for the relatively small numbers of birds missed in recent years on the coast. However, we urge more complete and synchronised counts of all wetlands potentially holding these species starting in the immediate future. Furthermore, it is clear that an additional source of error could be avoided in the future by organising coordinated counts in China to coincide with those undertaken regularly in South Korea and Japan (i.e. in mid-January). This would avoid both double counting within and between countries because of differential count coverage at sites along the flyway and would fall into line with the coordinated counts across the region that feed into the annual Asian Waterbird Census.
Table 5. Coastal survey counts of the two swan and four goose species in February during 2007–2009 and their proportional contributions to the overall estimates for the whole of China.
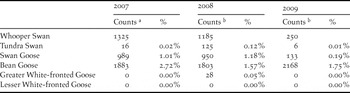
As shown in Table 1, Cao et al. (Reference Cao, Barter, Lei and Yang2008b, Reference Cao, Zhang, Barter and Lei2010) demonstrated that more than 95% of the most numerous swan and goose populations (except for Bean Goose) in China are distributed in Jiangxi, Hunan and Anhui Provinces, which were the best surveyed of wetlands in the Middle and Lower Yangtze floodplain in recent decades. Although this reduces the possibilities of missing large numbers of swan or goose populations in Jiangsu, Shanghai and Hubei (where numbers have been generally low and habitat degradation most rapid), in an ideal world, these areas should be covered on a similar basis to ensure the areas do not hold larger numbers than is currently thought to be the case. This is especially important for the Bean Goose, where it is thought that missed birds in these areas could potentially add more to the total numbers present.
The lack of fully synchronised surveys might introduce errors into the estimates because of population movements (within and between countries) during survey periods. However, satellite tracking showed that Swan Geese generally do not undertake long distance flights once settled during the northern winter (USGS 2009, Batbayar et al. Reference Batbayar, Takekawa, Newman, Prosser, Natsagdorj and Xiao2011). Movements of Greater White-fronted Geese were also relatively limited in Japan during the non-breeding season (Shimada Reference Shimada2009). Whilst these limited data suggest that geese move little within a season, there is a need to confirm this is the case through more extensive telemetry tracking of swans and geese. Such tracking would also fill an existing knowledge gap by enabling better definition of discrete subpopulations and hence effective management units within this flyway.
In the case of the Whooper Swan, due to its more northerly distribution along the coast compared to other swans and geese, the current surveys of the Yangtze River provide no accurate population information. For this species, as well as the nominate race of the Bean Goose (spending the non-breeding period in Xinjiang Province) and the Greylag Goose, there are urgent needs to extend the count coverage in China to generate better estimates, which also requires more cooperation and organisation of nationwide waterbird monitoring.
The coverage of the nationwide survey in South Korea is believed to be sufficient to estimate the total number of non-breeding swans and geese with a high degree of confidence, since it was designed to simultaneously estimate the number of waterbirds occurring in all known and potential sites in a relatively restricted geographical area. Many of the areas were repeatedly surveyed by the same, experienced observers or local researchers, so data quality is regarded as highly reliable, particularly for swans and geese. One two-day survey every January also minimises potential bias resulting from counts at different times of year or the possibility of double counts. Because of the decline of Tundra Swans in South Korea, there were some concerns about the reliability of swan data (e.g. possible misidentification of roosting Tundra and Whooper Swans) in the early part of the survey period. However, subsequent surveys have indicated real declines in the non-breeding numbers of Tundra Swans, which are not thought to be the result of human error; nevertheless, the cause of the declines remain unknown.
Assessments of swan and goose populations in East Asia flyway
Whooper Swan
The breeding range of the East Asian population of Whooper Swan extends from Central and Eastern Siberia to north-east China. In the non-breeding season individuals mostly stay in East Asia, including Korea, Japan and China with very small numbers recorded in North America (Mitchell Reference Mitchell1998). In China, the non-breeding range is confined to coastal wetlands along the Gulf of Bohai, of which Rongcheng and Yellow River Delta National Nature Reserve are the two most important sites. As survey data in China are not systematically gathered, the seemingly stable non-breeding population trend presented here needs to be viewed with some caution. It is more likely that this population will show adverse trends in numbers in this part of the range because of severe habitat loss and degradation within this region caused by land reclamation and industrial development in very recent years (Ma et al. Reference Ma, Melville, Liu, Chen, Yang, Ren, Zhang, Piersma and Li2014). In Japan, the main non-breeding range of Whooper Swan is along the Pacific coast in Hokkaido, and north of Ibaragi prefecture on the Pacific coast, and north of Niigata prefecture on the Sea of Japan coast. The non-breeding population of Whooper Swans in Japan increased in the 1980s and 1990s, partly due to an expansion in supplementary feeding (Albertsen and Kanazawa Reference Albertsen and Kanazawa2002). Satellite tracking of Whooper Swans showed the breeding area of the swans that spend the non-breeding period in Japan to be the middle Indigirka River and the lower parts of the Kolyma River in Russia (Kanai et al. Reference Kanai, Sato, Ueta, Minton, Higuchi, Soma, Mita and Matsui1997, Shimada et al. Reference Shimada, Yamaguchi, Hijikata, Hiraoka, Hupp, Flint, Tokita, Fujita, Uchida, Sato, Kurechi, Pearce, Ramey and Higuchi2014). Whooper Swans have expanded their breeding range in Yakutia, which includes the Indigirka River and the Kolyma River, since 1960s (Syroechkovski Reference Syroechkovski2002). Climate change in both the breeding and non-breeding areas was also suggested as a cause of population increase. Koyama et al. (Reference Koyama, Kasahara and Abe2013) showed that the mean highest daily temperatures in May on the breeding grounds had a positive effect on the subsequent proportions of non-breeding Whooper Swans that were juveniles, while the total snowfall of both previous and present years in the non-breeding range had a negative effect on this measure. As the most abundant swan in Korea (Park Reference Park2002), in the non-breeding season, its range is mainly confined to the west and south coasts, although they also commonly occur on inland lakes and reservoirs. Non-breeding birds in South Korea have been mainly linked with breeding populations in north-east Mongolia and the Daurian region of Russia, based on satellite telemetry studies (Newman et al. Reference Newman, Iverson, Takekawa, Gilbert, Prosser, Batbayar, Natsagdorj and Douglas2009). Resighted colour markings also support the main linkage with Mongolia, whereas some groups from Khabarovsk in Far Eastern Russia, such as Bolonskiy Nature Reserve, also spend the non-breeding period in the Nakdong River watershed (C. Y. Choi, unpubl. data), which supports two internationally important wetlands for this species in the south-eastern parts of the Korean Peninsula. Although numbers have been slowly increasing, in some areas, numbers have shown local declines due to habitat loss. Supplementary feeding has been implemented to compensate the reduction in natural food sources in Busan, for instance.
Tundra Swan
The Tundra Swan breeds in Central and East Siberia, mostly to the east of the Lena Delta (Rees et al. Reference Rees, Bowler and Beekman1997, Rees and Beekman Reference Rees and Beekman2010). Non-breeding sites are located throughout East Asia, including Korea, Japan and China. In China, pre-1996, the non-breeding range extended from the Yangtze River floodplain to inland Henan and Jiangsu, coastal Shanghai and Zhejiang (Cao et al. Reference Cao, Barter and Lei2008a). Based on a survey conducted in February 2003/04 and 2004/05, the majority (> 90% in China) were counted in Jiangxi and Anhui Province during the non-breeding season (Barter et al. Reference Barter, Chen, Cao and Lei2004, Reference Barter, Lei and Cao2006), illustrating a dramatic contraction in their distribution by that time (Cong et al. Reference Cong, Cao, Fox, Mark, Rees, Jiang, Ji, Zhu and Song2011). Two key sites, Poyang Lake and Baidang Lake, still retain abundant non-breeding numbers. Decreasing numbers in other lakes are thought to be related to the disappearance of submerged macrophytes, which might be caused by eutrophication, hydrological change and/or extensive aquaculture (Cong et al. Reference Cong, Cao, Fox, Mark, Rees, Jiang, Ji, Zhu and Song2011, Fox et al. Reference Fox, Cao, Zhang, Barter, Zhao, Meng and Wang2011). In Japan, the main non-breeding range of the Tundra Swan is in the south of Miyagi and Yamagata prefectures, north Chiba prefecture on the Pacific coast, north Ishikawa prefecture on the Sea of Japan coast, Lake Biwa in Shiga prefecture, Lake Nakaumi in Tottori prefecture, and Lake Shinjiko in Shimane prefecture. The numbers of Tundra Swan occurring in Japan in the non-breeding season increased in 1980s and the first half of 1990s. As is the case for the Whooper Swan, supplementary feeding has been suggested as a cause of the recent increases (Albertsen and Kanazawa Reference Albertsen and Kanazawa2002). The swans mainly forage in rice fields, feeding on leftover rice grains during daytime (Watanabe Reference Watanabe2003, Reference Watanabe2004). The increase in extent and availability of waste rice grain is suggested as the cause of the increase in numbers of Great White-fronted Geese (Shimada Reference Shimada1999), and this therefore could also contribute to similar increases amongst Tundra Swans. Climate change is also suggested as a contributory factor implicated in the increase. Lower snowfall in the non-breeding areas and higher temperatures on the breeding and stopover areas had positive effects on the swan population (Higuchi et al. Reference Higuchi, Koike and Shigeta2009). Historically, the Tundra Swan was once regarded as being as common locally as Whooper Swans in Korea (Austin Reference Austin1948). The Asian Waterbird Census (Li et al. Reference Li, Bloem, Delany, Martakis and Quintero2009) recorded 1,478 Tundra Swans with 738 Whooper Swans in 1994 at Nakdong River Estuary, suggesting many non-breeding groups still co-existed with Whooper Swans in Korea until the 1990s (Park Reference Park2002). However, in more recent years, their numbers have been rapidly declining and their non-breeding sites have been occupied by increasing numbers of Whooper Swans. Only small groups (fewer than 100–200 birds) are believed to now spend the non-breeding period annually in South Korea, but the reason for this decline is still unknown. Whooper Swans have been increasingly replacing Tundra Swans in Korea, which suggests that the decline in the Tundra Swan is not solely due to adverse conditions on the non-breeding grounds. It could be caused by declines in discrete breeding populations (e.g. associated with bad weather or adverse changes in habitat; Rees and Beekman Reference Rees and Beekman2010), or by simple displacement of non-breeding swans from Korea to Japan or China. If the latter is true, given the close proximity of the main non-breeding areas in south-east Korea and Japan, the Korean non-breeding populations may continue to gradually be attracted to Japan.
Swan Goose
The key breeding grounds of the Swan Goose lie in border areas between Russia, Mongolia and China. It also breeds in the lower reaches of the Amur river, north-western Sakhalin Island and Lake Khanka, Russia, western Mongolia and China (BirdLife International 2001, 2014). Non-breeding sites are almost exclusively confined now to China, although a very few spend the non-breeding period in Korea and fewer in Japan (Kear Reference Kear2005). Although around 1,000–2,000 were recorded in coastal Jiangsu and Fujian (Cao et al. Reference Cao, Zhang, Barter and Lei2010), it seems that almost the entire global population now spends the non-breeding period in the Yangtze River floodplain in China (Zhang et al. Reference Zhang, Cao, Barter, Fox, Zhao, Meng, Shi, Jiang and Zhu2011). Supporting about 99% of the national non-breeding population, Poyang Lake and Baidang Lake are currently the two most important sites for Swan Goose during the non-breeding season. As with Tundra Swans, Swan Geese mainly feed on submerged macrophytes in wetlands along the Yangtze River. Reduction in the extent and availability of submerged macrophytes, especially Vallisneria greatly constrains its non-breeding habitat, which has led to dramatic declines in numbers at lakes like Shengjin Lake (Fox et al. Reference Fox, Cao, Zhang, Barter, Zhao, Meng and Wang2011, Zhang et al. Reference Zhang, Cao, Barter, Fox, Zhao, Meng, Shi, Jiang and Zhu2011). Although non-breeding numbers seem to be fluctuating rather than decreasing, there is no doubt that this species is highly sensitive to habitat change due to its specialised diet and hence extreme habitat selection. Swan Geese in Korea are mainly passage migrants: the estuary of Yalu River (Amnok River) between China and North Korea has been identified as a major staging area for breeding populations from Mongolia and the Daurian region (Batbayar et al. Reference Batbayar, Takekawa, Newman, Prosser, Natsagdorj and Xiao2011). Another important staging area is known to be situated at the junction of Han River and Imjin River between South and North Korea, with a peak count of 1,858 geese in March 1998 (Park Reference Park2002). Given the movements of satellite-tracked Swan Geese from Mongolia heading for China (Batbayar et al. Reference Batbayar, Takekawa, Newman, Prosser, Natsagdorj and Xiao2011), and the resighting of Russian neck collars (C. Y. Choi, unpubl. data), it seems likely that the migrants staging on the Han and Imjin Rivers and the c.100 geese counted in South Korea probably belong to a distinct population that breeds in the Khabarovsk Region in Far Eastern Russia. Given the declining trend amongst numbers seen on passage on the Han River estuary since the mid-2000s, this population is possibly declining in number but its ultimate non-breeding grounds are still unknown (but likely somewhere in southern China).
Bean Goose
Three breeding populations occur in our study area: serrirostris breeding on the Kamchatka Peninsula, which spend the northern winter in western parts of Korea, small areas of Japan and eastern China; middendorffii breeding in Yakutia, occurring during the non-breeding season in East China and the south-east Korean Peninsula; and middendorffii breeding in Sayan and Altai, migrating to Dongting Lake during the non-breeding period. Bean Geese were previously more widely distributed in eastern China, with large numbers occurring in Henan, Hubei, Hunan, Jiangxi, Anhui and Jiangsu before 1996 (Cao et al. Reference Cao, Barter and Lei2008a). However, more than 90% of Bean Geese are now concentrated in the Yangtze River floodplain provinces of Anhui, Jiangxi, Hunan and Hubei, with numbers having decreased greatly in the inland regions of Jiangsu Province (Cao et al. Reference Cao, Zhang, Barter and Lei2010). There is little published information on distributions of these two subspecies, except that most of the relatively small numbers of Bean Geese at Dongting Lake are middendorffii (Fox et al. Reference Fox, Cao, Barter, Rees, Hearn, Cong, Wang, Zhang, Dou and Shao2008) and almost all Bean Goose at Shengjin Lake are serrirostris (Cheng et al. Reference Cheng, Cao, Barter and Xu2009). The total numbers occurring in China during the non-breeding season have fluctuated greatly between years, probably because of movements between sites and variable count coverage. To generate population estimates for the races of Bean Goose, we made the highly conservative, but reasonable worst-case assumption that in China middendorffii occur almost exclusively at Dongting Lake while serrirostris is distributed more widely elsewhere. On this basis, we tentatively assess the non-breeding populations of middendorffi and serrirostris in China at 6,000 and 95,000 respectively. In Japan, the main non-breeding range of the Bean Goose is in Miyagi, Akita, Niigata, Ishikawa, Shiga, and Shimane prefectures, where total non-breeding numbers of Bean Geese have been stable since the late 1990s (Figure1). Most Bean Geese occurring in Japan are middendorffii. The main area used by the less common serrirostris is limited to two marshes in Miyagi Prefecture during the northern winter. According to the count in January 2011, 2,122 serrirostris were counted in these marshes, with 9,316 middendorffii counted elsewhere. On the other hand, serrirostris is the dominant and widespread subspecies in South Korea, while middendorffii is restricted to a few wetlands in the Nakdong River watershed, such as Upo Wetland and Junam Reservoir (Park Reference Park2002). Although these two subspecies have not been counted separately in the field, typically 5,000–10,000 Bean Geese are regarded as middendorffii based on their limited geographic distribution in Gyeonggnam Province and Busan: for instance, 9,186 out of 74,761 geese in 2009, and 4,585 of 59,942 geese in 2011. Based on these new count data, but mindful of the caveats relating to our detailed knowledge of the distribution of the two subspecies, we therefore tentatively estimate the East Asian non-breeding populations of middendorffii and serrirostris to be 18,000 and 157,000 respectively. Key sites were mapped for both middendorffi and serrirostris with the 1% threshold based on the five subpopulations defined in Wetlands International (2015) (i.e. 35 for middendorffii, Sayan/eastern China; 125 for middendorffii, Yakutia/East Asia; 800 for middendorffii, Okhotsk/Kamchatka-Japan; 1,015 for serrirostris, Central and Eastern Siberia; 40 for serrirostris, Kamchatka/Japan; see Figure 3). The Bean Goose remains a species for which there would be substantial benefit from expanded telemetry and individual marking efforts, as well as improved mid-January count coverage and subspecies differentiation throughout their ranges.

Figure 3. Map showing the key sites for the two subspecies of Bean Goose spending the non-breeding period in South Korea, Japan and China. Black circles - middendorffii, white circles - serrirostris. Half-black/half-white is for both.
Greater White-fronted Goose
The breeding range of the East Asian subspecies of Greater White-fronted Goose A. a. frontalis is the Arctic tundra of eastern Russia, and the non-breeding range extends from Honshu Island in Japan, through western and southern coasts of South Korea, and areas along the Yangtze River in China (Kear Reference Kear2005). Numbers in China have decreased in the early 2000s and have been fluctuating. There has also been a contraction of range within China (Cao et al. Reference Cao, Zhang, Barter and Lei2010). Dongting Lake used to be an important resort during the non-breeding season, however, since 2008/2009 very few birds have been counted there, thought to be due to hydrologically induced habitat change (Zhao et al. Reference Zhao, Cong, Barter, Fox and Cao2012). As a result, the population is now mainly concentrated at Poyang and Shengjin Lakes where they are associated with extensive areas of Carex meadow, which is their preferred food (Zhao et al. Reference Zhao, Cong, Barter, Fox and Cao2012). In Japan, the main non-breeding range of Greater White-fronted Goose includes the Miyagi, Niigata, Ishikawa and Shimane prefectures, where numbers have been increasing dramatically and extending northwards (Takekawa et al. Reference Takekawa, Kurechi, Orthmeyer, Sabano, Uemura, Perry and Yee2000, Shimada et al. Reference Shimada, Hatakeyama, Miyabayashi and Kurechi2005). The total number of geese counted increased from 31,740 in 1996 to 156,231 in 2011. Rice fields are the main foraging areas for the geese in Japan (Amano Reference Amano2009), and the increase in this source of food may be a contributory factor to the increase in population. Modernisation of rice farming has resulted in more rice left behind in fields by modern combined harvesters than was previously the case using conventional reaper machines (Shimada Reference Shimada1999, Reference Shimada2009). While overall numbers of geese have been increasing, the number of sites used by the species during migration and in the northern winter remains limited, and to more effectively protect the species in Japan, expansion of the number of these areas is desirable (Moriguchi Reference Moriguchi, Amano and Ushiyama2013). The numbers of Greater White-fronted Geese in Korea have fluctuated between 20,000 and 110,000 birds, but this has recently stabilised at around 80,000–90,000. Higher numbers have been reported in central and western parts of South Korea (e.g. the Cheorwon Basin, Han River Estuary and Cheonsu Bay) during passage, i.e. the peak count in November (Park Reference Park2002) than during the middle of the northern winter, suggesting possible migration through the Korean Peninsula of geese continuing to known or potentially unknown non-breeding areas in China. Based upon colour marked geese rehabilitated from Cheorwon Wildlife Rescue Centre in South Korea and subsequently resighted in Miyagi Prefecture (five cases) and Hokkaido (four cases) in Japan during 2007–2011 (C. Y. Choi, unpubl. data; M. Kurechi, pers. comm.), there appear to be linkages between Korean and Japanese non-breeding areas (perhaps from common breeding areas in the Anadyr region, Far Eastern Russia). This suggests that exchange between these groups is a possible factor to explain annual fluctuations in these two countries.
Lesser White-fronted Goose
The population breeds in Central and Far Eastern Siberia and mainly spends the non-breeding period in Eastern China, with a few in Korea and Japan. Lesser White-fronted Geese might have been more widespread in the 1980s–1990s and with higher numbers in Anhui, Jiangxi and Jiangsu Provinces than those reported now. Evidence showed that a large decline in abundance and contraction of range has occurred in recent decades (Wang et al. Reference Wang, Fox, Cong, Barter and Cao2012). The Lesser White-fronted Goose is now mainly confined to East Dongting Lake and the contraction of range and reduction in numbers are likely to be explained by its specialist feeding ecology. They are highly dependent on recessional grasslands to store fat during the autumn for use during the middle part of the northern winter, but such habitats are becoming increasingly rare due to adverse hydrological management. Carex sedge meadows are used for feeding during January, but this food does not enable geese to balance their energy budgets, so they lose fat accumulated earlier in autumn whilst feeding on this food source (Wang et al. Reference Wang, Fox, Cong and Cao2013). Counts have been stable since 2002/03, at c.20,000 individuals (Wang et al. Reference Wang, Fox, Cong, Barter and Cao2012) but show signs of falling since 2011 due to specific losses of their specialist habitat.
Volunteer-based waterbirds surveys are needed in China
Whilst Japan and Korea have had well organised and extensive waterbird surveys in place over several decades to track changes in the abundance and distribution of swans and geese, the same has not been the case for China. A long-term and large-scale monitoring programme is required urgently to meet the needs of effective conservation and management of non-breeding swans and geese in China, which should be extended to cover all non-breeding waterbirds. However, this sort of large-scale monitoring programme involves the simultaneous survey of very extensive areas (vast lengths of marine shorelines, river basins, wetlands and lakes). It is evident that such a professional undertaking would be impossible on the grounds of limited resources, time and money. The networks of organised volunteers in Japan and Korea show that such extended monitoring efforts can be achieved by “citizen scientists”. These models demonstrate that monitoring information in the form of bird counts collected by trained competent volunteers can be immensely valuable when effectively organised by professionals to plan, structure and coordinate the nature, frequency and duration of monitoring. Such a system would potentially extend the scope of monitoring far beyond that which paid professionals could ever achieve themselves (Goffredo et al. Reference Goffredo, Piccinetti and Zaccanti2004, Kadoya and Washitani Reference Kadoya and Washitani2007). Unfortunately, there is no such tradition of volunteer-based monitoring work in China, with very few volunteers involved in biodiversity monitoring programmes since the 1980s. Nevertheless, since the early 1990s, more and more “citizen-scientists” have taken part in organised bird watching activities and volunteers have begun to participate in some projects, such as the China Coastal Waterbird Census and the National Biodiversity Demonstration Monitoring Programme. Although their potential has yet to be fully developed (especially when compared to other countries like Japan), these programmes give some confidence for the establishment of such volunteer based monitoring networks in the future.
In Japan, volunteers have been organised by 50 prefectural and city governments to monitor non-breeding waterfowl at a range of study sites since 1970, establishing a long and reliable database upon which to show changes in waterbird abundance and distribution (Kasahara and Koyama Reference Kasahara and Koyama2010). Here, counters are either directly coordinated by the prefectural governments, or they work with local birdwatching organisations, which are financially contracted to coordinate the volunteers and deliver the requisite quality data. This system could potentially serve as a good model for organising volunteer-based waterbird monitoring in China, especially at a time when more and more citizens are actively participating in bird watching.
International cooperation to improve population estimates and conservation
For the future, it is essential to better organise systematic, synchronous and complete surveys internationally to generate improved population estimates, better trends over time and more efficient assessment of the contributions of key sites to the effective protection of each of the populations throughout the flyway. This can potentially be improved by creating better cooperation under the existing Asian Waterbird Census, which is the regional framework for such waterbird monitoring work and is recognised by all Ramsar countries and East Asian Australasian Flyway Partnership Partners as the main system for such data collection. There is also a particular need to establish a recognised list of core sites of importance for Anatidae throughout the flyway as a key objective for effective monitoring and management. At present, there is also a major lack of coordinated demographic monitoring in the flyway, for instance, the scoring field ratios of first calendar year birds to interpret patterns of annual production and the individual capture, marking and resighting of collared or leg-ringed birds to generate annual survival estimates. This is particularly the case in China where a significant number of swans and geese spend the non-breeding period over huge geographical areas, where such data are urgently needed to contribute to our understanding of the drivers of population change. We therefore consider it a priority to coordinate sampling of age ratios from all the most numerous swan and goose populations considered here, with a form of annual reporting to disseminate such information to users, stakeholders and policy makers. In this connection, it is also essential in the longer term to coordinate capture and individual marking and resighting of swans and geese throughout the flyway to better understand linkages between different parts of the non-breeding range (supplemented with telemetry studies where feasible) and to generate estimates of annual survival through application of capture-mark-recapture techniques. Ideally, more data from telemetry studies will benefit our knowledge on population size and trends, by identifying unknown non-breeding grounds, migration connectivity, and relationships between different non-breeding populations.
In light of the continued serious habitat loss and declines in abundance amongst key goose and swan species in China, work to secure such coordinated monitoring should be accomplished as soon as possible before we lose the best waterbird sites in China. Opportunities have presented themselves at recent meetings in this region, for instance, at the 26th International Ornithological Congress (IOC) in August 2014 in Japan and the 16th Wetlands International / IUCN Species Survival Commission (SSC) Goose Specialist Group meeting in November 2014 in China. Commitments from range states to work more effectively with each other have been made as a result of such meetings.
Finally, it is important to urge greater international cooperation involving all the scientists active throughout the flyway, especially with researchers and managers in Russia and Mongolia, where these populations exploit important breeding, staging and moulting sites, to achieve better flyway monitoring of these populations and their and conservation.
Supplementary Material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0959270915000386
Acknowledgements
The study was supported by the National Basic Research Program of China (973 Program, Grant No. 2012CB956104), the National Natural Science Foundation of China (Grant No. 31370416), State Key Laboratory of Urban and Regional Ecology, Chinese Academy of Sciences (No. SKLURE2014-2-3), Science and Technology Service Network Initiative Project of the Chinese Academy of Sciences (KFJ-EW-ZY-004-06) and Special Fund for Forest Scientific Research in the Public Welfare (201304310).










