



Non-technical Summary
Conodont microfossils extracted from limestone at “Fossil Hill” in central Nevada about 60 years ago were the first of Middle Triassic age (ca. 243 My) discovered in North America. These canoe-shaped elements were named Neogondolella constricta, a species that subsequently has been reported worldwide. However, the scope of this species has remained uncertain because it was based on comparatively small early growth stages characterized by features lost during growth. Abundant specimens of the N. constricta group recovered from the original type locality form the basis for documenting changes in morphology during their accretionary growth and provide better definition of the species. This facilitates discrimination of similar species, 10 of which are distinguished in Nevada, including four new taxa; most of these are also identified in British Columbia. Successive associations through the Fossil Hill strata include elements with similar platform shapes but different relative lengths, the younger species being longer. This succession is calibrated with established ammonoid fossil zones and contributes to a parallel conodont biozonation. Eurasian occurrences of the Neogondolella constricta group are assessed and their correlation with the Nevadan scheme summarized.
Introduction
The first conodont species described from Middle Triassic strata in North America was Neogondolella constricta (Mosher and Clark, Reference Mosher and Clark1965) from the classic outcrop of the Prida Formation at Fossil Hill in the Humboldt Range of central Nevada (Fig. 1). This species has subsequently been widely reported from elsewhere, yet the taxonomic scope of the species, and many similar taxa assembled here as N. ex gr. constricta, remain poorly known and variably interpreted. This partly arises from the nature of the holotype, which is a relatively small, early growth stage for which an ontogenetic series has never been documented. The characteristic ‘constricted’ morphology of the posterior platform in N. constricta is common in early growth stages of many contemporaneous neogondolellins (Kozur et al., Reference Kozur, Krainer and Mostler1994b), which also often lack modern descriptions. Poor definition of many related taxa has spawned a diverse nomenclature and inhibited a robust biozonation for the upper Anisian and lower Ladinian strata where N. ex gr. constricta are often the dominant taxa.

Figure 1. Maps of (1) Nevada and (2) British Columbia showing locations of sections and sites from which upper Anisian–lower Ladinian conodont collections are reported here.
In addition to the biostratigraphic utility of the group, it is noteworthy that Neogondolella constricta serves as the type species for Pridaella Budurov and Sudar, Reference Budurov and Sudar1989, a genus proposed to accommodate this group of allied forms. The authors of that genus offered no rationale for this proposal other than to imply that the multielement apparatus of those species differed in non-specified ways from that of the type species of Neogondolella, N. mombergensis Tatge, Reference Tatge1956, from the Germanic Basin. The natural conodont assemblage from Monte San Giorgio, Switzerland, proposed as that of Neogondolella (Rieber, Reference Rieber1980; Orchard and Rieber, Reference Orchard and Rieber1999; Goudemand et al., Reference Goudemand, Orchard, Urdy, Bucher and Tafforeau2011, fig. 2B, C) corresponds, in fact, to N. ex gr. constricta (see Orchard, Reference Orchard2005, fig. 10) from the Humboldt Range of Nevada, which would serve as an apparatus template for Pridaella should that of N. mombergensis be demonstrably different. The recent conclusion of Chen et al. (Reference Chen, Scholze, Richoz and Zhang2018) that N. haslachensis (Tatge, Reference Tatge1956), an endemic Germanic associate of N. mombergensis, has a non-bifid S3 element unlike the constricta group, suggests a potential justification for this nomenclatural change. However, the apparatus of N. mombergensis remains undescribed.
In this work, we present growth stages of topotype Neogondolella constricta from the upper Anisian Rotelliformis ammonoid Zone at Fossil Hill and reassess allied taxa from the Middle Triassic of Nevada and British Columbia (B.C.). We also compare these taxa with similar conodont faunas first described from Europe fifty years ago that were central to the formulation of European conodont zonation; the extent to which these can be applied in North America is controversial. We aim to stabilize species concepts, establish stratigraphic ranges in North America, and facilitate comparison with similar taxa reported from Eurasia.
Previous work
The Prida Formation at Fossil Hill (and nearby Saurian Hill) in Nevada is an important paleontological site that, apart from its pioneering conodont research, has been the focus for ammonoid studies for over a century (Smith, Reference Smith1914; Silberling, Reference Silberling1962; Silberling and Nichols, Reference Silberling and Nichols1982; Monnet and Bucher, Reference Monnet and Bucher2005a, Reference Monnet and Bucherb). Notably, it provides the standard American ammonoid biochronology for the upper Anisian–lower Ladinian, which comprise the (Anisian) Rotelliformis, Meeki, Occidentalis, and (Ladinian) Subasperum ammonoid zones (Silberling and Tozer, Reference Silberling and Tozer1968), as well as constituent subzones (Fig. 2). Conodont studies at Fossil Hill began with Mosher and Clark (Reference Mosher and Clark1965), who introduced Neogondolella constricta. This was followed by major and very different taxonomic revisions by Nicora and Kovács (Reference Nicora and Kovács1984) and Ritter (Reference Ritter1989). The application of nomenclature developed in Europe was challenged by Kozur et al. (Reference Kozur, Krainer and Mostler1994b), who introduced new Neogondolella taxa for some of the Nevadan fauna based on published illustrations. More recent summaries of the conodont succession across the Anisian–Ladinian boundary (ALB) at Fossil and Saurian hills were presented by Bucher and Orchard (Reference Bucher and Orchard1995) and Orchard (Reference Orchard2010), but differentiation of the N. constricta group was not attempted.

Figure 2. Composite stratigraphic section for Fossil and Saurian hills, Nevada, showing ammonoid horizons and zonal divisions (after Silberling and Nichols, Reference Silberling and Nichols1982) and the relative positions of conodont samples collected by M.J. Orchard (OF) and H. Bucher (HB). Sample numbers with dots are ammonoid-bearing samples. Subzones are abbreviated within the Rotelliformis Zone: bu. = burckhardti, cl. = clarkei, vo. = vogdesi, cr. = cricki, bl. = blakei; subzones are abbreviated within the Meeki Zone: ne. = nevadanus, me. = meeki, du. = dunni; and subzones are abbreviated within the Occidentalis Zone (positions uncertain): hy. = hyatti, hu. = humboldtensis, fu. = furlongi, ga. = gabbi. Subzones below the blakei Subzone have been revised as a single vogdesi Subzone (Monnet and Bucher, Reference Monnet and Bucher2005a). The original position of the type Neogondolella constricta is indicated by the large arrow.
Contemporaneous Canadian sections where both ammonoids and conodonts are known were the subject of studies by Mosher (Reference Mosher1973), Orchard and Tozer (Reference Orchard, Tozer, Moslow and Wittenberg1997), and Golding (Reference Golding2014). In B.C., Neogondolella ex gr. constricta are represented in many disjunct sections of the Toad and Liard formations, and the Vega Member of the Sulphur Mountain Formation. These strata include ammonoids of the Deleeni, Meeki, and Chischa zones in the upper Anisian, and of the Matutinum, Poseidon, and Meginae zones in the lower Ladinian (Tozer, Reference Tozer1994). The most recent ammonoid-based correlation of these B.C. ammonoid zones and those from Nevada was presented by Ji and Bucher (Reference Ji and Bucher2018).
Neogondolella ex gr. constricta also occurs elsewhere in western and northern Canada (e.g., Orchard, Reference Orchard, Orchard and McCracken1991, Reference Orchard, Colpron and Nelson2006; Orchard et al., Reference Orchard, Cordey, Rui, Bamber, Mamet, Struik, Sano and Taylor2001; Henderson et al., Reference Henderson, Golding and Orchard2018). Neither these nor recent descriptions of new Anisian conodonts from B.C. (Golding and Orchard, Reference Golding and Orchard2016, Reference Golding and Orchard2018) have featured analysis of the Neogondolella constricta group.
In Europe, much of the pioneering Middle Triassic conodont work was undertaken in Bulgaria (Budurov and Stefanov, Reference Budurov and Stefanov1972, Reference Budurov and Stefanov1973), from where several key species were first described. As was the norm at the time, published images of these early types are of poor resolution, which inhibits comparison. Contemporaneous Muschelkalk faunas were described from the relatively restricted Germanic Basin (Tatge, Reference Tatge1956; Trammer, Reference Trammer1975; Zawidzka, Reference Zawidzka1975; Rafek, Reference Rafek1976), while those representing the larger Tethyan oceanic region were described from Austria (Kozur and Mostler, Reference Kozur and Mostler1982), Italy (Nicora and Brack, Reference Nicora and Brack1995), Greece (Krystyn, Reference Krystyn and Zapfe1983), and Hungary (Kovács et al., Reference Kovács, Nicora, Szabo and Balini1990; Kovács, Reference Kovács1994). On the southern margin of Tethys, the Sephardic Province represented an additional faunal realm (Hirsch, Reference Hirsch, Guex and Baud1994) with unique attributes.
Knowledge of the stratigraphic and geographic distribution of Balkan, Germanic, and Tethyan neogondolellin conodont taxa in North America should improve zonation and illuminate paths of migration within and between Europe and eastern Panthalassa. In recent decades, many studies from China, particularly from Guizhou Province, have reported Neogondolella constricta and allied forms (Wu et al., Reference Wu, Yao, Ji and Wang2008; Lehrmann et al., Reference Lehrmann, Stepchinski, Altiner, Orchard and Montgomery2015), which broadens their correlation potential further.
The evolving taxonomy
Identity of Neogondolella constricta group members has been hampered by taxonomic and nomenclatural uncertainty. Juvenile growth stages of several species may be indistinguishable because species-diagnostic features only become apparent with later growth. In their original report of the Fossil Hill conodonts, Mosher and Clark (Reference Mosher and Clark1965) described a poorly diversified conodont succession dominated by segminiplanate (neogondolellin) platform conodonts that they assigned to three species of the genus Gondolella: G. constricta n. sp., G. mombergensis, and G. navicula Huckriede, Reference Huckriede1958. In the same year, Bender and Stoppel (Reference Bender and Stoppel1965) introduced the genus Neogondolella with G. mombergensis as the type species for Triassic taxa resembling Gondolella, a genus now confined to the Pennsylvanian. In this paper, the original binomial names of species are used first, and thereafter current generic nomenclature is applied.
The original Fossil Hill species reported by Mosher and Clark (Reference Mosher and Clark1965), Kovács and Nicora (Reference Kovács1984), and Ritter (Reference Ritter1989) included the Germanic Neogondolella mombergensis. Kovács and Nicora (Reference Kovács1984) recognized two subspecies that they assigned to N. m. mombergensis and N. m. longa Budurov and Stefanov, Reference Budurov and Stefanov1973, the latter based on a Balkan taxon. Ritter's (Reference Ritter1989) univariate and multivariate morphometric analyses of 18 successive Neogondolella Pa element populations from Fossil Hill concluded that the speciation criteria used by Kovács and Nicora (Reference Kovács1984) were arbitrary and unsuitable for discrimination of zones. Rather, Ritter (Reference Ritter1989) assigned all the conodonts to a single, morphologically diverse species, for which he regarded N. mombergensis as the priority name. However, as noted by Orchard (in Bucher and Orchard, Reference Bucher and Orchard1995) and illustrated by Orchard and Rieber (Reference Orchard and Rieber1999, fig. 1), N. mombergensis is a distinctive species that differs from all those so far identified in North America.
A second species originally reported from Fossil Hill by Mosher and Clark (Reference Mosher and Clark1965) was Gondolella navicula, but this is now recognized as a Late Triassic Norigondolella species (Kozur, Reference Kozur1990b) with a differing apparatus (Orchard, Reference Orchard2005). Hence, N. constricta retains nomenclatural seniority for the common elements of the upper Anisian fauna at Fossil Hill.
In Europe, early work in Bulgaria by Budurov and Stefanov (Reference Budurov and Stefanov1972, Reference Budurov and Stefanov1973, Reference Budurov and Stefanov1975a) introduced new upper Anisian–lower Ladinian neogondolellin conodont species that provided a standard for comparison. A decade later, Nicora and Kovács (Reference Nicora and Kovács1984) concluded that two of those Balkan species, N. cornuta Budurov and Stefanov, Reference Budurov and Stefanov1972, and N. balkanica Budurov and Stefanov, Reference Budurov and Stefanov1975a, were in fact later growth stages, and therefore junior synonyms of N. constricta. Further revisions of the constricta group taxa were undertaken by Budurov and Stefanov (Reference Budurov and Stefanov1984), Kovács et al. (Reference Kovács and Kozur1980, Reference Kovács, Nicora, Szabo and Balini1990), Kozur and Mostler (Reference Kozur and Mostler1982), Kovács (Reference Kovács1994), and Kozur et al. (Reference Kozur, Krainer and Mostler1994b). Kovács et al. (Reference Kovács, Nicora, Szabo and Balini1990) formalized the ‘growth stages’ of N. constricta first as morphotypes α and β, and later (Kovács, Reference Kovács1994) as the subspecies N. constricta cornuta (= morphotype α) and N. c. balkanica (= β); a third morphotype (from Hungary) identified as morphotype γ was later named N. c. postcornuta (Kovács, Reference Kovács1994). In contrast, Kozur et al. (Reference Kozur, Krainer and Mostler1994b) argued that (1) the Balkan species N. cornuta and N. balkanica were not conspecific with N. constricta; (2) the newly named N. c. postcornuta was an example of the previously described Gondolella (= N.) mesotriassica Kozur and Mostler, Reference Kozur and Mostler1982; and (3) Nevadan elements formerly assigned to N. mombergensis represented a new species, N. aldae (with two subspecies). Kozur et al. (Reference Kozur, Krainer and Mostler1994b) argued that none of the European taxa previously recorded at Fossil Hill occurred there.
Stratigraphic utility
Middle Triassic neogondolellin species, many of which are allied with N. constricta, have featured in biostratigraphic zonations under various guises. Neither Mosher and Clark (Reference Mosher and Clark1965) nor Ritter (Reference Ritter1989) recognized a difference in the stratigraphic range of the dominant species at Fossil Hill, although Sweet et al. (Reference Sweet, Mosher, Clark, Collinson, Hasenmueller, Sweet and Bergstrom1971) differentiated an upper Anisian constricta Zone and a Ladinian mombergensis Zone in their standard Triassic zonation. Kovács and Nicora (Reference Kovács1984) maintained these two ill-defined zones and noted that the contact between them was coincident with the base of the Occidentalis ammonoid Zone; they found no change in conodonts between the Occidentalis and Subasperum zones in Nevada. Studies by the first author (Bucher and Orchard, Reference Bucher and Orchard1995; Orchard, Reference Orchard2010) have documented more diverse faunas that include species of Paragondolella, Budurovignathus, and probable new genera that collectively provide an improved biozonation for the interval, but the constricta group is far more common.
Middle Triassic Muschelkalk conodont faunas from the Germanic Basin were divided into seven zones by Kozur (Reference Kozur1968b) but many of the constituent taxa appear confined to that basin. In Bulgaria, a Balkan Anisian conodont zonation featured successive range zones of Paragondolella bulgarica Budurov and Stefanov, Reference Budurov and Stefanov1975a, Ozarkodina (= Nicorella) kockeli Tatge, Reference Tatge1956, P. excelsa Mosher, Reference Mosher1968, Neogondolella cornuta, N. excentrica Budurov and Stefanov, Reference Budurov and Stefanov1972, and N. bakalovi Budurov and Stefanov, Reference Budurov and Stefanov1972 (Budurov and Stefanov, Reference Budurov and Stefanov1975b, Reference Budurov and Stefanov1983; Budurov, Reference Budurov1980; Budurov and Trifonova, Reference Budurov and Trifonova1994, Reference Budurov and Trifonova1995; Budurov and Vaptsarova, Reference Budurov and Vaptsarova1994; Budurov et al., Reference Budurov, Trifonova and Zagorčev1995). This Balkan succession is not obviously applicable in Nevada, and neither is that of Kozur (Reference Kozur1980) who summarized his earlier work and that of Mosher (Reference Mosher1968) to propose a “standard zonation” for the interval that featured successive conodont zones of N. constricta, N. pseudolonga Kovács, Kozur, and Mietto, Reference Kovács, Kozur and Mietto1980, and N. transita Kozur and Mostler, Reference Kozur and Mostler1971. Later, Kozur (Reference Kozur1990a) presented a revised zonal scheme for Tethys that included three successive assemblage zones of N. constricta, G. (= N.) praetrammeri Kozur and Mostler, Reference Kozur and Mostler1982, plus N. mesotriassica, and N. transita.
To some extent these various zonations can be aligned (see Nicora, Reference Nicora and Gaetani1993), but differing taxonomic concepts, resultant nomenclature, and geographic provenance hinder correlation. Similarly, the absence in North America of Tethyan Gladigondolella spp. and ‘Neogondolella’ trammeri Kozur in Kozur and Mock, Reference Kozur and Mock1972, group; the Germanic endemics N. mombergensis, N. haslachensis, and Gondolatus spp.; and the Sephardic Pseudofurnishius restricts their global use. In contrast, some N. ex gr. constricta are not geographically restricted.
The most recent accounts of conodonts from the ALB sections in Hungary and Italy arose during deliberations concerning the choice of the global stratigraphic section and point (GSSP) for the stage boundary (Kovács, Reference Kovács1994; Nicora and Brack, Reference Nicora and Brack1995; Brack and Nicora, Reference Brack and Nicora1998). Accounts of the Bagolino GSSP section, Italy, document occurrences of 11 members of the Neogondolella constricta group, including taxa regarded here as synonyms and others unknown in North America. Later, although no taxonomic discussions were presented, the summaries provided by Muttoni et al. (Reference Muttoni, Nicora, Brack and Kent2004) differentiated only six members, whereas Brack et al. (Reference Brack, Rieber, Nicora and Mundil2005) included just five. Notably, these works do not record all the same taxa. A synthetic summary of the published constricta group conodont occurrences in Europe is given in Figure 3.

Figure 3. European ammonoid zones and the ranges of Neogondolella constricta group members within the late Anisian–early Ladinian based on the literature. Those of Southern Alps of Italy after Kozur et al. (Reference Kozur, Krainer and Mostler1994b), Nicora and Brack (Reference Nicora and Brack1995), Brack et al. (Reference Brack, Rieber, Nicora and Mundil2005), and Muttoni et al. (Reference Muttoni, Nicora, Brack and Kent2004); the Balaton Highlands, Hungary after Kovács (Reference Kovács1994) and Vörös et al. (Reference Vörös, Szabó, Kovács, Dosztály and Budai1996); and from Bulgaria after Budurov and Stefanov (Reference Budurov and Stefanov1972). Elements allied with N. transita are probable examples of N. excentrica of this work. Synonymies of N. balkanica and N. pseudolonga are discussed in the text.
Materials
The foundations for this paper are numerous and often abundant collections of conodonts recovered from North American strata that have provided an ammonoid biochronology for the late Anisian and early Ladinian (Tozer, Reference Tozer1967, Reference Tozer1994; Silberling and Tozer, Reference Silberling and Tozer1968; Monnet and Bucher, Reference Monnet and Bucher2005a, Reference Monnet and Bucherb) (Figs. 4, 5). Nearly all of the 100+ conodont collections recovered are dominated by the Neogondolella constricta group.

Figure 4. Summary of observed ranges of Neogondolella constricta group members with respect to the late Anisian–early Ladinian Nevadan ammonoid zones and subzones at Fossil and Saurian hills. Ammonoid zonation after Silberling and Tozer (Reference Silberling and Tozer1968), Silberling and Nichols (Reference Silberling and Nichols1982), and Monnet and Bucher (Reference Monnet and Bucher2005a, Reference Monnet and Bucherb). Three intervals of N. constricta group assemblages are shown on the right. New taxa identified in this work are Neogondolella quasiconstricta n. sp., Neogondolella excentrica primitiva n. subsp., Neogondolella quasicornuta n. sp., and Neogondolella excentrica sigmoidalis n. subsp.

Figure 5. Summary of observed ranges of Neogondolella constricta group members with respect to the late Anisian–early Ladinian ammonoid zones in British Columbia (see Table 2). Ammonoid zonation after Tozer (Reference Tozer1994), and Monnet and Bucher (Reference Monnet and Bucher2005b). Three intervals of N. constricta group assemblages are shown on the right. New taxa identified in this work are Neogondolella excentrica primitiva n. subsp., Neogondolella quasiconstricta n. sp., Neogondolella excentrica sigmoidalis n. subsp., and Neogondolella quasicornuta n. sp.; aff. = Neogondolella excentrica aff. primitiva n. subsp.
In 1992, the Fossil Hill section and the nearby Saurian Hill section were sampled for conodonts in conjunction with H. Bucher, who guided the first author through the sections of the Prida Formation in which ammonoid faunas of the Rotelliformis, Meeki, Occidentalis, and Subasperum zones were identified. Additional matrix samples were taken from ammonoid collections, including bed-by-bed samples from an excavated trench exposure at Saurian Hill. Successions of 35 conodont collections from Fossil Hill and 37 from Saurian Hill were recovered (Table 1), in addition to others from nearby Wheeler Mine in the northern Humboldt Range. Some contemporaneous ammonoid-bearing samples were also collected from the Tobin Range (Rotelliformis and Meeki zones) (Fig. 1; Table 2). In total, about 80 samples from Nevada were processed and nearly all produced large conodont fauna.
Table 1. Occurrences of Neogondolella constricta group members at Fossil Hill (above) and Saurian Hill (below), Nevada. Ammonoid zones are indicated above the sample numbers, which are in bold when the collections contain at least 100 constricta group elements. See text for relative abundances of species.

Table 2. Occurrences of the Neogondolella constricta group members from spot samples in other mostly late Anisian–early Ladinian localities in British Columbian and Nevada. GSC curation numbers are given for these collections.

Small Middle Triassic conodont samples from Canadian ammonoid matrices were originally provided by E.T. Tozer to Mosher (Reference Mosher1973). Later, the first author collected bulk samples during 1983 field work with E.T. Tozer in the Toad–Liard River area. Follow-up fieldwork along the Alaska Highway by the present authors in 2011 yielded supplementary collections from locations that underpin parts of the standard Canadian ammonoid biochronology (Tozer, Reference Tozer1967, Reference Tozer1994). Upper Anisian and lower Ladinian strata yielded conodonts from nine sections that collectively spanned Deleeni through Meginae ammonoid zones, with representation of all but the Meeki Zone (Table 2). In addition, matrix samples were obtained from archival ammonoid collections representing the Deleeni Zone on Chischa River, the Matutinum Zone at Wapiti Lake, and the type localities of the Deleeni (Alaska Highway, milepost 375) and Poseidon (Tuchodi Bluffs) zones. Additional suites of contemporaneous samples were collected from sections of the Whistler and Llama formations near Wapiti Lake (1997–1998), and from the Toad Formation exposed at Ursula Creek on Williston Lake (1992, 1999, 2001) (Fig. 1).
Repository and institutional abbreviation
Illustrated material is deposited in the National Type Collections of the Geological Survey of Canada and bear 6-digit type numbers prefixed with “GSC”. Supporting collections bear the following GSC curation numbers: C-300201–300236 (Fossil Hill); C-201563–201566, C-201581–201594, C-201572, C-209954, C-209955, C-300240–300252, C-301228 (Saurian Hill).
Results and discussion
Many of these recovered collections contain hundreds of conodont elements, or part elements, including good representation of the multielement ramiform components (Orchard, Reference Orchard2005, fig. 10). The P1 platform elements representing the Neogondolella constricta group dominate most collections. Among them, the ‘conservative’ N. constricta and N. cornuta along with many less-specific early growth stages predominate, with the latter species largely replacing the former early in the Meeki Zone. Concurrently, N. posterolonga became increasingly common into the Ladinian. Far less common are eight other taxa: N. aldae, N. excentrica primitiva new subspecies, N. e. excentrica, N. e. sigmoidalis new subspecies, N. ex gr. mesotriassica, N. postcornuta, N. quasicornuta new species, and N. quasiconstricta new species. To date, 10 taxa have been recognized in the less-diverse faunas recovered from Canadian locations (Table 2).
The reconstructed growth series of topotype Neogondolella constricta from beds containing Rotelliformis Zone ammonoids at Fossil Hill, Nevada, reveals growth progression from early growth stage elements with a constricted posterior platform (as in the holotype) through progressively broader elements that have a narrowly and then broadly rounded posterior platform. An inclined terminal denticle lies posterior of the cusp and becomes increasingly prominent as growth proceeds: it may lie at the posterior end of the platform, or a narrow posterior platform brim may be developed.
The ontogenetic and other variation in platform configuration shown within topotype N. constricta populations are here considered to embrace diagnostic features of the Balkan species N. cornuta and N. balkanica (as was also determined by Nicora and Kovács, Reference Nicora and Kovács1984), and of the later differentiated N. tardocornuta Budurov and Stefanov, Reference Budurov and Stefanov1984. The growth and variation seen in N. constricta are essentially the same as those observed in the more elongate species that succeeds it, for which N. cornuta is regarded as the appropriate name. The two Nevadan species display similar posterior morphogenesis but can be distinguished by their platform shape and length:breadth ratio. Based on the relative platform dimensions of the holotypes, we suggest that both N. balkanica and N. tardocornuta are junior synonyms of N. constricta, and that the Alpine N. pseudolonga is an early growth stage of N. cornuta or perhaps N. posterolonga. Restudy of these European species from their type localities, as has been done here for N. constricta, will be necessary to confirm these suggested synonymies.
The two subspecies of Neogondolella aldae introduced by Kozur et al. (Reference Kozur, Krainer and Mostler1994b), N. a. aldae and N. a. posterolonga, are raised to species level. These species, which retain the juvenile feature of a narrow posterior platform, are reported from the upper Anisian and lower Ladinian in both the USA and Canada, and from European localities (see below). These species were formerly confused with the Germanic Neogondolella mombergensis and Balkan N. longa, neither of which are certainly known from North America.
Later growth stages of both Neogondolella constricta and N. cornuta have essentially rounded, symmetrical posterior platforms of uniform width, but they commonly co-occur with elements of similar relative length that have either asymmetric or expanded and truncated posterior platforms. Hence, in Nevada, N. constricta is associated with the asymmetric N. excentrica primitiva n. subsp. and the quadrate N. quasiconstricta n. sp. in the Rotelliformis Zone, and later N. cornuta co-occurs with N. quasicornuta n. sp. and the asymmetric N. e. excentrica in the Occidentalis Zone. Most of the intervening Meeki Zone contains the asymmetric N. e. aff. cornuta, which may have an accessory posterior denticle but lacks a secondary posterior process. In the Ladinian Subasperum Zone, rare N. e. sigmoidalis n. subsp. occurs (Fig. 4), often with N. posterolonga.
These associations exhibit symmetry transition in their posterior platform configuration that is like that shown by the Pelsonian triad Paragondolella bulgarica, P. hanbulogi Sudar and Budurov, Reference Sudar and Budurov1979, and P. bifurcata Budurov and Stefanov, Reference Budurov and Stefanov1972. As a backdrop to this pattern, two major trends are recognized within the constricta group. The first is an elongation and relative narrowing of the platform, which is quite abrupt early in the Meeki Zone; the second is the elaboration of the posterior platform to produce a longer, more differentiated secondary process and extended keel in N. excentrica, starting with N. aff. N. cornuta.
In B.C., these taxa are less common, but N. constricta and N. e. primitiva n. subsp. occur in the Deleeni Zone; N. cornuta occurs with N. e. excentrica in the uppermost Anisian Chischa Zone; and N. e. sigmoidalis n. subsp. occurs in association with N. posterolonga in the Ladinian Matutinum and Poseidon zones, and maybe into the Meginae Zone based on its occurrence with the ammonoid Silenticeras bamberi Fauna, which is known to be bracketed between the Poseidon and Meginae III zones (Tozer, Reference Tozer1994, p. 33). No conodonts are known from the Meeki Zone in B.C., but an appearance of N. cornuta earlier than N. e. excentrica (N. transita of previous authors) would align with Nevadan and European sections (Figs. 3, 4). The interval with N. e. sigmoidalis n. subsp. may be equivalent in age (and perhaps in content) to the Tethyan transita Zone (Kozur, Reference Kozur1990a) and the Balkan bakalovi Zone.
One outcome of this study is that Neogondolella constricta sensu stricto is resurrected as an upper Anisian species that demonstrably appears prior to N. cornuta in Nevada and B.C. and thus identifies an interval that predates a cornuta Zone. A constricta Zone has not been widely used in Europe recently, but a cornuta Zone has been regarded as immediately succeeding a Paragondolella excelsa Zone in the much older Trinodosus Zone. Occurrences of N. cornuta at these levels may be based on similar elements with a prominent posterior denticle, which has been regarded as diagnostic for that species.
The “standard zonation” for the ALB interval proposed by Kozur (Reference Kozur1980) features the successive conodont zones of N. constricta, N. pseudolonga, and N. transita. This succession compares moderately well with the Nevadan succession because N. pseudolonga may represent an early growth stage of the contemporaneous N. cornuta or N. posterolonga, and N. transita may include elements here assigned to N. excentrica sigmoidalis n. subsp.
The Tethyan zonal scheme of Kozur (Reference Kozur1990a), with successive assemblage zones of N. constricta, N. praetrammeri – N. mesotriassica, and N. transita has less applicability in North America because the trammeri group does not occur, and the scope of N. mesotriassica is unclear.
The main conodont events recognized in the Balaton Highland, Hungary by Kovács (Reference Kovács1993a, Reference Kovács and Gaetanib; Fig. 3) also have limited expression in Nevada where the first event, the appearance of N. postcornuta, is not readily discernible due to its rarity, and the second event, the appearance of ‘N.’ trammeri, is not known. A third event in Hungary (Kovács, Reference Kovács1993a, Reference Kovács and Gaetanib) features the appearance of N. transita near the base of the Curionii Zone, which may also be comparable with the appearance of N. e. sigmoidalis n. subsp.
The conodont succession at the ALB GSSP at Bagolino (Fig. 3) in the southern Alps of Italy is unclear because different species have been recorded by Nicora and Brack (Reference Nicora and Brack1995), Muttoni et al. (Reference Muttoni, Nicora, Brack and Kent2004), and Brack et al. (Reference Brack, Rieber, Nicora and Mundil2005). The present account represents a North American taxonomic yardstick for comparison through which the identity and morphological scope of these European species may be better understood.
Conclusions
A study of upper Anisian and early Ladinian conodonts from classic Middle Triassic sites in Nevada and B.C. focused on related taxa united as the Neogondolella constricta group. These are by far the most common components of the North American faunas as well as having a worldwide distribution. Their potential to contribute to a temporal framework for the interval has not been fully realized due to conflicting taxonomic interpretations. This is rooted in the choice of an early growth stage as the holotype of N. constricta, an original description that lacks an ontogenetic component, and poorly defined group members. This work provides an improved definition for the central species based on abundant growth stages of topotype material, and critically reviews previous records of the species group.
The distinction of Neogondolella constricta from N. cornuta, and the suggested synonymy of several other taxa (N. balkanica, N. tardocornuta, N. pseudolonga), is here newly based on relative platform dimensions rather than differences in posterior platform–cusp configuration (nature of terminal denticle, presence of platform brim), which varies during growth and is regarded as intraspecific. The nature and distribution of the allied species N. aldae and N. posterolonga, based on Nevadan material, is documented from both North America and Europe, whereas the European-based taxa N. bakalovi, N. longa, N. mesotriassica, and N. postcornuta, are comparatively rare or absent in the present faunas.
Less common elements of the upper Anisian constricta group conodont faunas are those that show elaboration of the posterior platform through a change in symmetry and the development of accessory denticles. Among posteriorly curved platforms that have been previously assigned to Neogondolella excentrica and N. transita, three subspecies are differentiated, two of which are new: N. e. primitiva n. subsp. and N. e. sigmoidalis n. subsp. Elements characterized by a broader, ‘truncated’, and denticulate posterior platform are assigned to N. quasiconstricta n. sp. and N. quasicornuta n. sp., taxa that are morphological analogues of the older Paragondolella bifurcata and the Germanic Gondolella (= N.) prava Kozur, Reference Kozur1968a.
In North America, the species N. constricta and N. cornuta are associated with comparable elements showing the extended posterior symmetries: (1) N. constricta + N. excentrica primitiva n. subsp. + N. quasiconstricta n. sp.; and (2) N. cornuta + N. e. excentrica + N. quasicornuta n. sp. (Fig. 4). Calibrated with the Nevadan ammonoid zonation, the first conodont assemblage (1) occurs alone in the Rotelliformis Zone and is joined and overwhelmed by the second association (2) by the Occidentalis Zone. In the intervening Meeki Zone, cornuta-like elements have an accessory posterior denticle but no extended process. A different association (3) is characterized by N. posterolonga and N. e. sigmoidalis n. subsp. and occurs in the Ladinian Subsaperum Zone.
These intervals can also be recognized in B.C.: the constricta association in the Deleeni Zone; the cornuta association in the Chischa Zone; and the posterolonga fauna in the Matutinum, Poseidon, and ?Meginae (bamberi fauna) zones (Fig. 5). The ranges of less common species of the constricta group are variable but generally long ranging through the interval under study. Nevadan faunas are also more diverse than those from B.C., as may be expected from their lower paleolatitude. Many of the North American constricta group representatives also occur in Europe and farther afield.
Correlation with Neogondolella ex gr. constricta elements in Europe and their succession is unclear due to differing taxonomic criteria employed for species differentiation. Nevertheless, a similar pattern can be discerned within the various zonal schemes proposed in Europe, although some elements do not occur in North America, namely the Germanic N. mombergensis – Ng. haslachensis lineage, and the Tethyan “N.” ex gr. trammeri and Gladigondolella spp. Re-assessment of European faunas, especially those described here as N. excentrica subspp., may support the proposed North American succession.
Systematic paleontology
Taxonomic scope
The Neogondolella constricta group includes segminiplanate platform-bearing species known or suspected to be characterized by an early growth stage with a narrow, constricted posterior platform, and a relatively low blade–carina. Included among these are species that are only known as later growth stages that resemble other constricta group members. The group is centered on the holotype of N. constricta from the Prida Formation of Nevada, a juvenile element from the upper Anisian Rotelliformis Zone. Very similar juvenile elements occur throughout the upper Anisian and lower Ladinian strata worldwide, but they mature into a variety of different species.
Differentiation of members of the constricta group must generally rely on morphological features of adult elements. The following previously described taxa (with original genus name) are discussed here: Neogondolella aldae Kozur, Krainer and Mostler, Reference Kozur, Krainer and Mostler1994b; Neogondolella bakalovi Budurov and Stefanov, Reference Budurov and Stefanov1972; Neogondolella balkanica Budurov and Stefanov, Reference Budurov and Stefanov1975a; Gondolella constricta Mosher and Clark, Reference Mosher and Clark1965; Neogondolella cornuta Budurov and Stefanov, Reference Budurov and Stefanov1972; Neogondolella excentrica Budurov and Stefanov, Reference Budurov and Stefanov1972; Neogondolella longa Budurov and Stefanov, Reference Budurov and Stefanov1973; Gondolella mesotriassica Kozur and Mostler, Reference Kozur and Mostler1982; Gondolella postcornuta Kovács, Reference Kovács1994; Neogondolella posterolonga Kozur, Krainer and Mostler, Reference Kozur, Krainer and Mostler1994b; Gondolella pseudolonga Kovács, Kozur, and Mietto, Reference Kovács, Kozur and Mietto1980; Neogondolella tardocornuta Budurov and Stefanov, Reference Budurov and Stefanov1984; and Gondolella transita Kozur and Mostler, Reference Kozur and Mostler1971.
Class Conodonta Pander, Reference Pander1856
Order Ozarkodinidae Dzik, Reference Dzik1976
Family Gondolellidae Lindström, Reference Lindström1970
Subfamily Neogondolellinae Hirsch, Reference Hirsch, Guex and Baud1994
Genus Neogondolella Bender and Stoppel, Reference Bender and Stoppel1965
Type species
Gondolella mombergensis Tatge, Reference Tatge1956 (Ta 1956/5) from the upper Muschelkalk, Schmidtdiel Quarry, Momberg, near Marburg, Germany.
Neogondolella constricta (Mosher and Clark, Reference Mosher and Clark1965)
Figures 6, 7
- p* Reference Mosher and Clark1965
Gondolella constricta Mosher and Clark, p. 560, pl. 65, figs. 11, 18, 21, 24, 25 (only).
- p Reference Mosher and Clark1965
Gondolella mombergensis Tatge; Mosher and Clark, p. 560, pl. 65, figs. ?20, 28 (only).
- ? Reference Budurov and Stefanov1975a
Neogondolella balkanica Budurov and Stefanov, p. 792–794, pl. 1, figs. 24–31.
- p Reference Rafek1976
Neogondolella constricta; Rafek, pl. IV, fig. 24 (only).
- p Reference Rafek1976
Neogondolella cf. longa Budurov and Stefanov; Rafek, pl. I, figs. 34, 35 (only).
- Reference Kovács, Kozur and Mietto1980
Gondolella constricta; Kovács and Kozur, pl. 3, fig. 4a–c.
- Reference Kozur, Krainer and Mostler1981
Neogondolella cornuta; Mietto and Petroni, p. 552–553, pl. 57, figs. 5a, b.
- Reference Kozur, Krainer and Mostler1981
Neogondolella longa; Mietto and Petroni, p. 553–554, pl. 57, figs. 8a–c.
- non Reference Papšová and Pevný1982
Neogondolella constricta; Papšová and Pevný, Pl. XX, figs. 7–12.
- ? Reference Budurov and Stefanov1984
Neogondolella tardocornuta Budurov and Stefanov, p. 605–607, pl. 1, figs. 15–20.
- Reference Farabegoli, Levanti, Perri and Veneri1984
Neogondolella cornuta; Farabegoli et al., figs. a1–3, b1–3.
- p Reference Kovács1986
Gondolella constricta; Kovács, pl. IV, fig. 4a–d; pl. V, figs. 5a–d, 6a–d; pl. VI, figs. 3a, b, 4a, b; pl. X, figs. 4a–c, 5a–c, 6a–c (only).
- non Reference Dȕrkoop, Richter and Stritzke1986
Gondolella constricta; Dürkoop et al., pl. 21, figs. 1a–c, 2a, b.
- non Reference Ding and Huang1990
Neogondolella constricta; Ding and Huang, pl. 1, figs. 2, 3.
- Reference Koca, Gedik and Balcioğlu1992
Neogondolella constricta; Koca et al., pl. 1, figs. 11a–c, 13.
- Reference Koca, Gedik and Balcioğlu1992
Neogondolella cf. cornuta; Koca et al., pl. 1, figs. 15a–c.
- Reference Chhabra and Kumar1992
Neogondolella constricta; Chhabra and Kumar, pl. 3, figs. 2, 3a, b, 4a–c, 6a, b.
- Reference Kozur, Krainer and Mostler1994b
Neogondolella constricta; Kozur et al., p. 172–174, pl. 4, figs. 12–15, 17, 20.
- non Reference Polak, Havrila, Filo and Pevný1996
Neogondolella constricta; Polak et al., pl. 12, figs. 1–5.
- ? Reference Buryi, Baud, Popova, Dickins, Lucas and Zacharov1997
Neogondolella constricta; Buryi, pl. 1, fig. 15.
- non Reference Pevný and Salaj1997
Gondolella constricta; Pevný and Salaj, p. 101–102, pl. IX, figs. 14, 15; pl. X, figs. 1, 2; ?pl. XI, figs. 2, 3.
- Reference Pevný and Salaj1997
Gondolella balkanica; Pevný and Salaj, p. 103, pl. XI, figs. 6–8; pl. XV, figs. 5–8.
- Reference Orchard and Rieber1999
Neogondolella ex gr. constricta; Orchard and Rieber, pl. 4., figs. 1–12 (multielement).
- p Reference Orchard, Colpron and Nelson2006
Neogondolella ex gr. constricta; Orchard, pl. 5., figs. 9, 10 (only).
- non Reference Sun, Sun, Hao and Jiang2006
Neogondolella constricta; Sun et al., pl. 1, figs. 21–24.
- non Reference Dong and Wang2006
Neogondolella constricta; Dong and Wang, pl. 41, figs. 10, 18.
- non Reference Wu, Yao, Ji and Wang2008
Neogondolella constricta; Wu et al., pl. II, figs. 12–15.
- p Reference Wu, Yao, Ji and Wang2008
Neogondolella mombergensis (Clark and Mosher) (sic); Wu et al., pl. II, figs. 4, 5, 18, 22.
- p Reference Wu, Yao, Ji and Wang2008
Neogondolella navicula (Clark and Mosher) (sic); Wu et al., pl. I, figs. 2, 3, 7, 10.
- non Reference Zhang, Zhou, Lu, Xie, Lou and Liu2009
Neogondolella constricta; Zhang et al., figs. 3. 13, 16.
- Reference Orchard2010
Neogondolella constricta; Orchard, fig. 9. 14, 15.
- Reference Golding2014
Neogondolella ex gr. constricta morphotype gamma; Golding, fig. 2.24, parts 1–6.
- Reference Golding2014
Neogondolella ex gr. constricta morphotype delta; Golding, fig. 2.25, parts 1–6.
- non Reference Sun, Jiang, Sun and Hao2014
Neogondolella constricta; Sun et al., Reference Sun, Jiang, Sun and Hao2014, fig. 3h, k–m.
- Reference Lehrmann, Stepchinski, Altiner, Orchard and Montgomery2015
Neogondolella ex gr. constricta; Lehrmann et al., fig. 6.9, 6.10.
- non Reference Bo, Yao, Xiao, Bai and Peng2017
Neogondolella constricta; Bo et al., fig. 4.1–4.4.
- Reference Zhang, Orchard, Algeo, Chen, Lyu, Zhao, Kaiho, Ma and Liu2019
Neogondolella constricta; Zhang et al., fig. 7.13a–c.
- non Reference Xie, Liu, Lou, Hu, Zhou, Huang and Wen2019
Neogondolella constricta; Xie et al., fig. 3.2, 3.3, 3.7.
- non Reference Golding2021
Neogondolella ex gr. constricta (Mosher and Clark); Golding, p. 584, pl. 3., figs. 29–35.
- non Reference Qin, Golding, Jiang, Chen, Zhang, Kang, Wang and Yuan2021
Neogondolella constricta; Qin et al., pl. 8., figs. 8–13.

Figure 6. Neogondolella constricta (Mosher and Clark) from the Rotelliformis Zone, Fossil Hill, Nevada. (1–3) USNM 145189, refigured holotype, clarkei Subzone; (4–6) GSC 141848, FH15, clarkei Subzone; (7) GSC 141849, FH6, burckhardti Subzone; (8–10) GSC 141850, FH24, cricki Subzone; (11, 12) GSC 141851, FH8, burckhardti Subzone.; (13–15) GSC 141852, FH17, vogdesi Subzone; (16, 17) GSC 141853, FH22, cricki Subzone. Scale bar = 200 μm.

Figure 7. Neogondolella constricta (Mosher and Clark) from B.C. (1–3) GSC 141854, sample 83/205B, Toad Formation, Toad River Canyon, Deleeni Zone; (4, 5) GSC 141855, and (9–11) GSC 141856, both sample 92/AH2, Yellow Bluffs, Alaska Highway, Deleeni Zone; (6–8) GSC 141857, sample 97/WapA6, Llama Formation, Cirque B, Ganoid Ridge, bracketed between the Poseidon and Meginae zones. Scale bar = 200 μm.
Holotype
USNM 145189 (Mosher and Clark, Reference Mosher and Clark1965, pl. 65, figs. 21, 24, 25). Re-illustrated in Figure 6.1–6.3. From sample FH3, clarkei Subzone of the Rotelliformis Zone, upper Anisian, Prida Formation, Fossil Hill, Nevada; deposited in the US National Museum of Natural History.
Diagnosis
The relatively short and arched segminiplanate P1 element has at first a slender, biconvex platform that broadens with growth and maintains a typical length breadth ratio of ~4:1. A posterior platform constriction present in juveniles disappears during growth as the posterior platform expands and becomes as broad as the median platform. The rounded posterior margin of later growth stages may include a narrow brim around a large, upright to slightly reclined terminal denticle that generally becomes larger than the cusp during growth, which is typically the penultimate carina denticle. During growth, the anterior blade denticles become increasingly elevated relative to those of the median carina, which become increasingly fused. On the underside, a symmetrical basal loop of uniform width surrounds the subterminal pit.
Occurrence
Neogondolella constricta occurs in Nevada throughout the Fossil Hill Member of the Prida Formation at Fossil Hill, where it is abundant (many hundreds of specimens) in the Rotelliformis Zone, but much less so through the Meeki and Occidentalis zones, and into the Subasperum Zone. Elsewhere in Nevada, the species is known from the Wheeler Mine area, and in the Tobin Range. In B.C., it is particularly common in the Deleeni Zone, including in the type locality of that zone and additional locations nearby. Elsewhere in B.C. it ranges through the Poseidon Zone (Table 2).
Neogondolella constricta has a global reach: upper Muschelkalk, northern Germany (Rafek, Reference Rafek1976); Campogrosso section, NE Italy (Mietto and Petroni, Reference Mietto and Petroni1981); middle Bivera Formation, southern Alps, northern Italy (Farabegoli et al., Reference Farabegoli, Levanti, Perri and Veneri1984); Alsóhegy, N. Hungary (Kovács and Kozur, Reference Kozur1980); NE Rudabánya Mountains, Hungary (Kovács, Reference Kovács1986); eastern Turkey (Koca et al., Reference Koca, Gedik and Balcioğlu1992); Kalapani Limestone, northern India (Chhabra and Kumar, Reference Chhabra and Kumar1992); western Carpathians, Slovakia (Pevný and Salaj, Reference Pevný and Salaj1997); chert of Tsentralnaya Mts, Sikhote–Alin (Buryi, Reference Buryi, Baud, Popova, Dickins, Lucas and Zacharov1997); subsurface Polish lowlands (Narkiewicz, Reference Narkiewicz1999); Jones Lake Formation, distal allochthon, south-central Yukon Territory (Orchard, Reference Orchard, Colpron and Nelson2006); Xinyuan Formation, Guandao, Nanpanjiang Basin, South China (Lehrmann et al., Reference Lehrmann, Stepchinski, Altiner, Orchard and Montgomery2015); Kamura Formation, Kamura, central Kyushu Island, Japan (Zhang et al., Reference Zhang, Orchard, Algeo, Chen, Lyu, Zhao, Kaiho, Ma and Liu2019). In China, several reports of N. constricta are discounted (see synonymy) or are insufficiently illustrated (Wang and Wang, Reference Wang and Wang1976; Wang et al., Reference Wang, Wang, Li and Wei2005), but the species does occur in the Qingyan Formation in Guizhou Province (Wu et al., Reference Wu, Yao, Ji and Wang2008). Those reported from Bithynian strata at Deşli Caira, Romania (Golding, Reference Golding2021) are now excluded from this species.
Description
The holotype is a comparatively small, elongate element with a platform that extends throughout the length of the element but narrows at a posterior constriction near the cusp and expands around the final denticle. The axial blade–carina is composed of ~15 relatively low, closely spaced denticles, the anterior ones of which are slightly larger, higher, and more discrete than those of the median carina. The cusp is the penultimate denticle and is twice the size of the adjacent posteriormost denticle. Later growth stages show the posterior platform constriction progressively overgrown to produce at first a tapered posterior outline and then a broad, rounded posterior outline equal in breadth to that of the median part. Concurrently, the anterior denticles of the fixed blade become relatively higher, those of the median carina become increasingly fused into a low ridge, and the posteriormost denticle enlarges and dominates the posterior margin; a narrow posterior platform brim often develops. Very large elements may have some subdued anterior platform crenulation.
Comparisons
Adult specimens of Neogondolella constricta differ from those of N. cornuta in their comparatively shorter and broader platform (4–4.5:1 compared with 5–5.5:1), an outline that is commonly biconvex rather than subrectangular, and in their less conspicuous posterior carina and cusp. Both N. aldae and N. posterolonga differ from N. constricta by possessing platforms that are wider in the anterior and much narrower in the posterior.
Remarks
The distinctive posterior denticle (not the cusp) of Neogondolella cornuta is often larger than that in N. constricta, but later growth stages of both species can display a prominent and reclined terminal denticle, which has been regarded as diagnostic for N. cornuta. Variation in this feature led to separation of both N. balkanica (upright denticle with a platform brim) and N. tardocornuta (upright denticle without a brim) in the Balkans. Nicora and Kovács (Reference Nicora and Kovács1984) previously suggested that N. balkanica was synonymous with N. constricta, while N. tardocornuta was originally combined in N. balkanica by its authors (Budurov and Stefanov, 1975). The holotypes of both Balkan species have platform proportions (~4:1) comparable to that of N. constricta, and variability of the posterior margin in the Nevadan material is such that those species are provisionally included in synonymy here. As pointed out by Kozur et al. (Reference Kozur, Krainer and Mostler1994b), the types of all these species originated in different parts of the late Anisian–early Ladinian, but this does not exclude a long range for N. constricta.
Neogondolella aldae Kozur, Krainer and Mostler, Reference Kozur, Krainer and Mostler1994b
Figure 8
- p Reference Mosher and Clark1965
Gondolella navicula Huckriede; Mosher and Clark, p. 560–561, pl. 66, figs. 10, 17, 18, 21 (only).
- p Reference Nicora and Kovács1984
Gondolella mombergensis longa (Budurov and Stefanov, Reference Budurov and Stefanov1973); Nicora and Kovács, p. 150, pl. 10, figs. 2, 4, 9 (only).
- * Reference Kozur, Krainer and Mostler1994b
Neogondolella aldae Kozur, Krainer, and Mostler, p. 179–181.
- * Reference Kozur, Krainer and Mostler1994b
Neogondolella aldae aldae Kozur, Krainer, and Mostler, p. 181–182 (see synonymy).
- p Reference Wu, Yao, Ji and Wang2008
Neogondolella mombergensis (Mosher and Clark) (sic); Wu et al., pl. II, fig. 9.
- p Reference Golding2014
Neogondolella ex gr. transita, morphotype beta; Golding, fig. 2.29, parts 1–3.
- Reference Golding2014
Neogondolella constricta morphotype beta; Golding, fig. 2.23, parts 1–9.

Figure 8. Neogondolella aldae Kozur, Krainer, and Mostler from (1–10) the Toad Formation in B.C. and (11–19) the Prida Formation in Nevada. (1–3) GSC 141858, sample 92/AH21, Yellow Bluffs, Alaska Highway, Chischa Zone; (4, 5) GSC 141859, sample 92/AH2, Toad Formation, Yellow Bluffs, Alaska Highway, Deleeni Zone; (6–8) GSC 141860, sample 83/205B, Toad River Canyon, Deleeni Zone; (9, 10) GSC 141861, sample 83/ MJO-Bone. Toad River Canyon, ?Deleeni Zone; (11, 12) GSC 141862, sample FH47, Meeki–Occidentalis zonal boundary; (13–15) GSC 141863, sample SH516, Subasperum Zone; (16, 17) GSC 141864, sample SH531, Occidentalis Zone; (18, 19) GSC 141865, sample SH517, Subasperum Zone. Scale bar = 200 μm.
Holotype
The specimen, figured by Nicora and Kovács (Reference Nicora and Kovács1984, pl. 10, fig. 9) as Gondolella mombergensis longa Budurov and Stefanov. From sample N49 in the middle Prida Formation, Nevadites humboldtensis Smith, Reference Smith1914, beds of the lower Occidentalis Zone at Fossil Hill, Humboldt Range, Nevada.
Occurrence
Mosher and Clark (Reference Mosher and Clark1965) illustrated elements from the Meeki Zone in Nevada, and Kozur et al. (Reference Kozur, Krainer and Mostler1994b) gave a range of lower Meeki and Occidentalis zones. In the present study, the species was found to occur first in the middle of the Rotelliformis Zone and as high as the Subasperum Zone. Neogondolella aldae is one of the more common species of the constricta group in the Humboldt Range, being found in small numbers in about half the studied samples. Elsewhere in Nevada, the species is known from the Tobin Range. In Canada, the species occurs in the Deleeni and Chischa zones at Yellow Bluffs, in the Deleeni Zone of Toad River Canyon, in the Matutinum Zone at Wapiti Lake, and as high as the Meginae Zone along the Alaska Highway at milepost 386 (Table 2).
In Europe, Neogondolella aldae occurs in the Reitzi Zone and Nevadites Zone of the Alps and Bulgaria (Kozur et al., Reference Kozur, Krainer and Mostler1994b) but it has not featured in many studies. A specimen illustrated by Wu et al. (Reference Wu, Yao, Ji and Wang2008) from the Qingyan Formation in Guizhou, China, appears to represent this species.
Description
The platform of this arched segminiplanate element is widest around midlength, with biconvex margins in the anterior two-thirds to three-quarters, and subparallel margins in the remainder of the distinctly narrower posterior platform. Large specimens show increasing lateral growth in the posterior part, which remains narrower than the anterior platform. Overall, the typical length:breadth ratio is about 4:1. The narrowly rounded to subquadrate posterior margin of adult elements may include a very narrow platform brim around a large, moderately reclined terminal denticle, which is separated from the smaller cusp immediately in front of it; both these denticles commonly lie in the posterior constricted part. To the anterior, denticles of the carina of mature elements are largely fused into a low ridge that rises to the posterior and extends to the anterior relatively elevated fixed blade, which is commonly composed of 4 discrete denticles. On the underside of the largest specimens, the pit is anteriorly shifted within the basal scar that extends behind it.
Comparisons
Unlike Neogondolella constricta and N. cornuta, the posterior platform constriction present in juveniles of N. aldae does not disappear during growth as the posterior platform remains narrow. Neogondolella posterolonga differs in its longer and more slender platform. The older N. shoshonensis Nicora, Reference Nicora1976, has a similar platform outline but is narrower with a higher posterior carina.
Remarks
The original description of Neogondolella aldae (Kozur et al., Reference Kozur, Krainer and Mostler1994b) included reference to its distinct terminal “cusp” that was not fused with the posterior platform margin, unlike in N. cornuta. As discussed, previously, such fusion is variable and a more reliable character to distinguish the two species is their platform shape. Note that the true cusp in the adults of the constricta group is the penultimate denticle of the carina and not the larger terminal denticle.
Some Nevadan specimens previously illustrated by Nicora and Kovács (Reference Nicora and Kovács1984) and included in Neogondolella aldae by Kozur et al. (Reference Kozur, Krainer and Mostler1994b) do not exhibit the constricted posterior platform present in the holotype; these are excluded from the present species.
Neogondolella cornuta Budurov and Stefanov, Reference Budurov and Stefanov1972
Figure 9.1–9.6, 9.9–9.15
- p Reference Mosher and Clark1965
Gondolella constricta Mosher and Clark, p. 560, pl. 65, fig. 22 (only).
- p Reference Mosher and Clark1965
Gondolella mombergensis Tatge; Mosher and Clark, p. 560, pl. 65, fig. 29 (only).
- p Reference Mosher and Clark1965
Gondolella navicula Huckriede; Mosher and Clark, p. 560–561, pl. 66, figs. 19, 20 (only).
- * Reference Budurov and Stefanov1972
Neogondolella cornuta Budurov and Stefanov, p. 839–840, pl. 3, figs. 9–15, 20–22.
- Reference Trammer1975
Gondolella cornuta; Trammer, pl. 22, figs. 8a, b, 9a, b.
- ? Reference Kovács, Kozur and Mietto1980
Gondolella pseudolonga Kovács, Kozur and Mietto, p. 218–219, pl. 1, figs. 1–4.
- Reference Vrielynck1984
Gondolella cornuta; Vrielynck, p. 187–189, pl. 3, figs. 4a–c.
- p Reference Vrielynck1984
Gondolella pseudolonga; Vrielynck, p. 194–195, pl. 3, figs. 2a–c (only)
- Reference Orchard and Austin1986
Neogondolella constricta (Mosher and Clark); Orchard, pl. 5.4, fig. 8 (only).
- non Reference Dȕrkoop, Richter and Stritzke1986
Gondolella cornuta; Dürkoop et al., pl. 17, fig. 5a, b; pl. 20, figs. 13a–c, 15a, b.
- Reference Uroševič and Sudar1991
Neogondolella cornuta; Uroševič and Sudar, pl. 1, fig. 13.
- ? Reference Ramovš and Goričan1995
Neogondolella cornuta; Ramovš and Goričan, pl. 8, fig. 4.
- Reference Pevný and Salaj1997
Gondolella cornuta; Pevný and Salaj, pl. XII, figs. 3, 11.
- ? Reference Narkiewicz1999
Neogondolella cornuta > N. mesotriassica; Narkiewicz, pl. 3, figs. 3, 4.
- Reference Orchard2005
Neogondolella ex gr. constricta; Orchard, fig. 10A–I (multielement).
- non Reference Wang, Wang, Li and Wei2005
Neogondolella cornuta; Wang et al., pl. II, fig. 20.
- non Reference Dong and Wang2006
Neogondolella constricta cornuta; Dong and Wang, pl. 41, fig. 3.
- Reference Orchard, Colpron and Nelson2006
Neogondolella ex gr. constricta; Orchard, pl. 4, fig. 15.
- Reference Nakrem, Orchard, Weitschat, Hounslow, Beatty and Mørk2008
Neogondolella ex gr. constricta; Nakrem et al., fig. 5.18–5.20.
- Reference Orchard2010
Neogondolella cornuta; Orchard, fig. 9.12, 9.13.
- non Reference Sun, Jiang, Sun and Hao2014
Neogondolella constricta cornuta; Sun et al., fig. 3.g?, i, j.
- ? Reference Sun, Jiang, Ji and Hao2016
Neogondolella constricta cornuta; Sun et al., fig. 3.5.
- non Reference Xie, Liu, Lou, Hu, Zhou, Huang and Wen2019
Neogondolella constricta cornuta; Xie et al., fig. 3.12.
- non Reference Qin, Golding, Jiang, Chen, Zhang, Kang, Wang and Yuan2021
Neogondolella cornuta; Qin et al., pl. 3, figs. 3, 4; pl. 7, fig. 4; pl. 9, figs. 9–11.
- Reference Karádi, Budai, Haas, Vörös, Piros, Dunkl and Tóth2022
Neogondolella cornuta; Karádi et al., fig. 9D.
- Reference Karádi, Budai, Haas, Vörös, Piros, Dunkl and Tóth2022
Neogondolella pseudolonga; Karádi et al., fig. 9C.

Figure 9. (1–6, 9–15) Neogondolella cornuta Budurov and Stefanov from the Prida Formation, Nevada, Occidentalis Zone; (11–15) the Llama Formation, B.C., Chischa Zone. (1, 2) GSC 141866, sample FH56; (3, 4) GSC 141867, sample SH524; (5, 6) GSC 132583, sample SH524; (9, 10) GSC 141869, sample SH527; (11–13) GSC 141870; (14, 15) GSC 141871, both from sample 97/WapB18, Cirque B, Ganoid Ridge. (7, 8) Neogondolella aff. N. cornuta Budurov and Stefanov; GSC 141872, sample SH524. Scale bar = 200 μm (1–13), 250 μm (14, 15).
Holotype
Budurov and Stefanov, Reference Budurov and Stefanov1972, pl. 3, figs. 20–22, Bu 1045/1, from middle–upper Illyrian strata, III γ = cornuta conodont zone; Golo–Bârdo mountains south of Pernik, Bulgaria. Budurov's collections (Bu) are in the Geologisches Institut, Sofia, Bulgaria.
Occurrence
In Nevada, Neogondolella cornuta appears in the Meeki Zone, is abundant (many hundred specimens) through the Occidentalis Zone and is less common in the Subasperum Zone. It also occurs at Wheeler Mine and in the Tobin Range. In B.C., it is known from the Chischa and Poseidon zones at Wapiti Lake, and with the bamberi ammonoid fauna at milepost 386, Alaska Highway (Table 2).
In addition to the Bulgarian types that characterize the Illyrian (Budurov and Trifonova, Reference Budurov and Trifonova1995), Neogondolella cornuta is recorded in southwestern Holy Cross Mountains, Poland (Trammer, Reference Trammer1975); Carnic Alps–Dolomites, NE Italy (Vrielynck, Reference Vrielynck1984); Stuhini Group, Stikine terrane, northern B.C. (Orchard, Reference Orchard and Austin1986, Reference Orchard, Orchard and McCracken1991); eastern Serbia (Uroševič and Sudar, Reference Uroševič and Sudar1991); central Slovenia (Ramovš and Goričan, Reference Ramovš and Goričan1995); western Carpathians, Slovakia (Pevný and Salaj, Reference Pevný and Salaj1997); ?subsurface Polish lowlands (Narkiewicz, Reference Narkiewicz1999); Table Mountain Formation, Sylvester Allochthon, northern B.C. (Orchard, Reference Orchard, Colpron and Nelson2006); Botneheia Formation, Milne Edwardsfjellet, Svalbard (Nakrem et al., Reference Nakrem, Orchard, Weitschat, Hounslow, Beatty and Mørk2008), and the Transdanubian Range, Hungary (Karádi et al., Reference Karádi, Budai, Haas, Vörös, Piros, Dunkl and Tóth2022). In China, a probable juvenile growth stage of this species was illustrated by Sun et al. (Reference Sun, Jiang, Ji and Hao2016) from the Upper Guanling Formation in Guizhou Province, but other reports are suspect (see synonymy).
Description
A relatively elongate, arched segminiplanate P1 element with a comparatively long and slender subrectangular platform with subparallel platform margins and a typical length:breadth ratio of ~5–5.5:1. Submature elements show the characteristic biconvex anterior platform and constricted posterior that disappears during growth as the posterior platform broadens. A prominent, slightly to strongly reclined and terminal denticle (“horn”) is characteristic of mature elements but this may be overtaken by the cusp in the largest specimens, wherein a narrow platform brim may be developed on the broad, rounded posterior margin. During growth, the anterior blade denticles become slightly elevated relative to those of the median carina, which may be wholly fused. The lowest part of the carina is at element midlength, beyond which the posterior fused carina is often relatively elevated in front of the large posterior denticles. On the underside, the pit is anteriorly shifted within the basal loop, which extends a short distance to the posterior with a rounded or subquadrate outline.
Comparisons
Neogondolella cornuta was regarded as a later growth stage and therefore a synonym of N. constricta by Nicora and Kovács (Reference Nicora and Kovács1984), but in the present work the taxon is interpreted as its direct descendant. Small specimens of the two species may be indistinguishable, but the relatively long platform and longer posterior keel characteristic of N. cornuta become more common features in the constricta group populations through the upper Anisian. Posterior platform configuration may appear similar in the two species, but greater weight is given here to the relative platform length and outline.
Neogondolella aldae differs from N. cornuta in outline, with the relatively broad anterior and reduced posterior platform of the former contrasting with the sub-parallel margins of the latter. The platform of N. posterolonga is like that of early growth stages of N. cornuta and may have arisen through neotony. According to Kovács (Reference Kovács1994), the most diagnostic feature for distinguishing N. cornuta from its successor species N. postcornuta is the penultimate denticle of the carina in specimens of moderate size: this is a conspicuous cusp in the former, ancestral species but is not evident in N. postcornuta, which also has the terminal denticle fused to the platform.
Remarks
Gondolella pseudolonga, which is based on a submature growth stage from Italy (Kovács et al., Reference Kovács, Kozur and Mietto1980), has denticulation characteristic of relatively early growth stages of Neogondolella ex gr. constricta. The character of adult N. pseudolonga is conjectural, but the platform length to breadth ratio of its holotype is 5:1, which corresponds to that of both N. cornuta and N. posterolonga. Neogondolella pseudolonga may be a junior synonym of N. cornuta, although Kozur et al. (Reference Kozur, Krainer and Mostler1994b, p. 174) regarded it as an earlier growth stage and synonym of N. longa from the excentrica Zone of western Bulgaria. However, the holotype of N. longa has a length: breadth ratio of ~6:1 and is narrower than the present species, although its variability is unknown.
Neogondolella cornuta ladinica Kozur et al., Reference Kozur, Krainer and Mostler1994b, is based on elements with an extended basal scar on an upturned basal posterior margin, a feature that becomes more common through the range of the species in Austria, finally replacing the nominate subspecies. Differentiation of such forms has not proven possible in the North American material.
Neogondolella aff. N. cornuta Budurov and Stefanov, Reference Budurov and Stefanov1972
Figure 9.7, 9.8.
Remarks
These elements resemble Neogondolella cornuta in all respects other than they have a posteriormost denticle that is offset from the main carinal axis. They are distinctly longer than Neogondolella excentrica primitiva n. subsp., but show a similar, generally rudimentary posterior elaboration. In N. e. excentrica, a posterior secondary process and keel are developed, which are absent in N. aff. N. cornuta. About 85 elements with this morphology occur, always with N. cornuta; this is a little more common than N. e. excentrica.
Neogondolella excentrica Budurov and Stefanov, Reference Budurov and Stefanov1972
- *Reference Budurov and Stefanov1972
Neogondolella excentrica Budurov and Stefanov, p. 840–841, pl. 4, figs. 9–28.
Holotype
Bu 1707/1, from lα = excentrica conodont-Zone 1, Golo–Bârdo mountains south of Pernik, Bulgaria. Budurov's collections (Bu) are in the Geologisches Institut, Sofia, Bulgaria.
Description
The P1 elements assembled here are elongate segminiplanate conodonts in which a posterior platform constriction occurs in front of a variably inturned posterior platform that may bear up to 3 additional nodes posterior of the inconspicuous cusp. On the underside, one side of an asymmetrical basal scar extends as a posterior keel. In adult specimens, the anterior blade and posterior carina denticles are higher and more discrete than those of the more fused median carina.
Remarks
Many populations of the Neogondolella constricta group from Nevada and B.C. include posteriorly asymmetric elements with a variably inturned posterior platform process above a posterior keel. In a few specimens, there is no deflection of the posterior platform, although an extended keel is well developed, implying transition from other members of the constricta group. Most of these forms previously have been assigned to either N. excentrica or to N. transita. Kozur et al. (Reference Kozur, Krainer and Mostler1994b) regarded these two species as synonyms, with the subsymmetrical platform of N. transita being connected with the strongly curved platforms of N. excentrica through a morphological continuum. Budurov and Stefanov (Reference Budurov and Stefanov1972) included a variety of platform symmetries in their species, but none resembles the holotype of N. transita (platform ratio 4:1; straight axis; large, triangular cusp; upturned, pointed posterior platform tip; aff. N. suhodolica Budurov and Stefanov, Reference Budurov and Stefanov1973). As noted by Kovács (Reference Kovács1984), a comparative study of populations of each taxon in their respective type areas is needed to resolve the scope of each. In this work, we subdivide N. excentrica into three subspecies, two of which are new. These differ in their relative platform dimensions and posterior platform configuration. Records of N. transita in Hungary and Italy are shown as N. ex gr. excentrica in Figure 5.
Neogondolella excentrica excentrica Budurov and Stepanov, Reference Budurov and Stefanov1972
Figure 10.1?, 10.2–10.7
- Reference Mosher and Clark1965
Polygnathus tethydis Huckriede; Mosher and Clark, p. 563, pl. 66, fig. 13.
- * Reference Budurov and Stefanov1972
Neogondolella excentrica Budurov and Stefanov, p. 840–841, pl. 4, figs. 9–28.
- p Reference Trammer1975
Gondolella excentrica (Budurov and Stefanov); Trammer, pl. 25, figs. 4a–c (only).
- p Reference Zawidzka1975
Gondolella excentrica; Zawidzka, pl. 42, figs. 5a, b (only).
- Reference Zawidzka1975
Gondolella navicula Huckriede; Zawidzka, pl. 40, figs. 4a, b.
- Reference Rafek1976
Neogondolella transita Kozur and Mostler; Rafek, pl. IV, figs. 31–33.
- Reference Nicora1976
Neogondolella excentrica; Nicora, p. 639–640, pl. 84, figs. 3–5.
- Reference Mietto and Petroni1979
Neogondolella excentrica; Mietto and Petroni, p. 9, pl. I, figs. 5a–c.
- Reference Mietto and Petroni1981
Gondolella transita; Mietto and Petroni, p. 555–556, pl. 57, figs. 9a–c.
- ? Reference Mietto and Petroni1981
Gondolella basisymmetrica huckriedei (Budurov and Stefanov); Mietto and Petroni, p. 555–556, pl. 57, figs. 2a, b.
- Reference Kolar-Jurkovšek1983
Neogondolella excentrica; Kolar-Jurkovšek, p. 341–342, pl. 12, figs. 1a–d.
- Reference Vrielynck1984
Gondolella lindstroemi (Budurov and Stefanov); Vrielynck, p. 192–193, pl. 3, figs. 5a–c.
- ? Reference Uroševič and Sudar1991
Gondolella transita (Kozur and Mostler, Reference Kozur and Mostler1971); Uroševič and Sudar, pl. 1, fig. 16.
- Reference Márquez-Aliaga, Valenzuela-Rios, Calvet and Budurov2000
Neogondolella excentrica; Márquez-Aliaga et al., fig. 6.11.
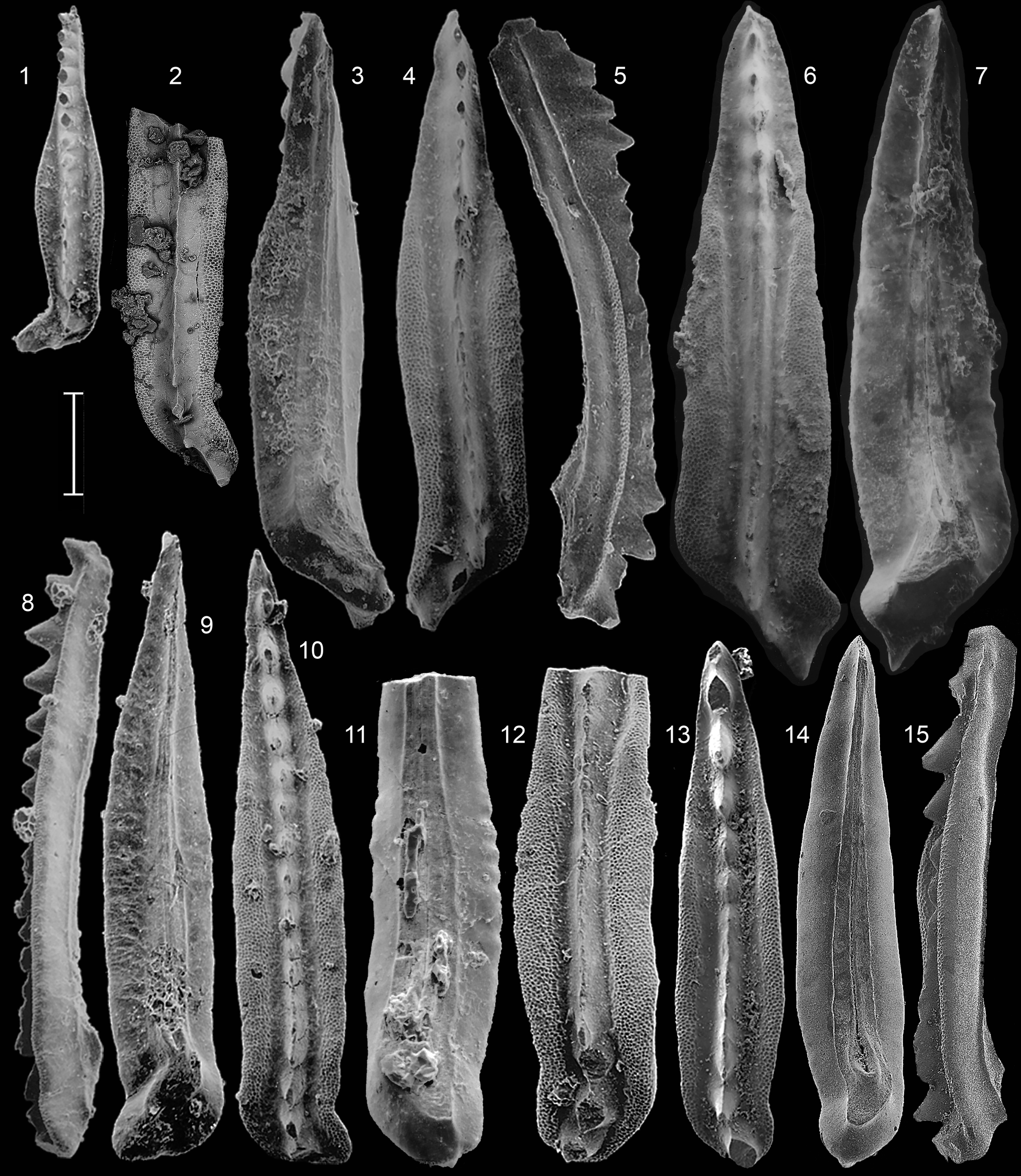
Figure 10. (1) Neogondolella excentrica aff. excentrica Budurov and Stefanov from Prida Formation, Fossil Hill, Nevada. GSC 141873, sample FH13, Rotelliformis Zone/clarkei Subzone. (2–7) Neogondolella excentrica excentrica Budurov and Stefanov from (2) Llama Formation, B.C. and (3–7) Prida Formation, Saurian Hill, Nevada. (2) GSC 141874, sample WAP-B17, Cirque B, Ganoid Ridge, Chischa Zone; (3–5) GSC 141875, sample SH529; (6, 7) GSC 141876, sample SH529, both Occidentalis Zone. (8–15) Neogondolella excentrica sigmoidalis n. subsp. from (8–12) sample SH512, Prida Formation, Nevada, Subasperum Zone, and (13–15) Llama Formation, B.C. (8–10) GSC 141877, holotype; (11, 12) GSC 141878; (13–15) GSC 141879, sample 97/WapA6, Cirque B, Ganoid Ridge, Wapiti Lake area, bracketed between Poseidon and Meginae zones. Scale bar = 200 μm.
Diagnosis
As for species plus this slender morphotype has a length:breadth ratio in adult specimens of ≥5:1; commonly subparallel platform margins; an asymmetric posterior margin bearing a tapered, often pointed and offset posterior platform; and a slightly to strongly extended postero-lateral basal keel. The posterior platform is typically inturned and may bear several denticles forming a secondary carina.
Occurrence
About 60 specimens of Neogondolella excentrica excentrica occur in collections from the Occidentalis and lower Subasperum zones in Nevada; the posterior fragment illustrated by Mosher and Clark (Reference Mosher and Clark1965) came from the Protrachyceras Beds. Nicora (Reference Nicora1976) also illustrated the species from Fossil Hill. Rare specimens of N. e. aff. excentrica occur in the Rotelliformis Zone. The subspecies occurs also in Chischa Zone on Ganoid Ridge, and as high as the bamberi Fauna along the Alaska Highway in B.C.
In Europe, Kozur et al. (Reference Kozur, Krainer and Mostler1994b) noted specimens like these occurred in the Nevadites Zone and lower Curionii Zone. Examples of this subspecies are reported from southwestern Holy Cross Mountains, Poland (Trammer, Reference Trammer1975); Upper Muschelkalk, northern Germany (Rafek, Reference Rafek1976); ?San Ulderico section, NE Italy (Mietto and Petroni, Reference Mietto and Petroni1979); Campogrosso section, NE Italy (Mietto and Petroni, Reference Mietto and Petroni1981); Idrske Krnice, Slovenia (Kolar-Jurkovšek, Reference Kolar-Jurkovšek1983); Carnic Alps-Dolomites, NE Italy (Vrielynck, Reference Vrielynck1984); Eastern Serbia (Uroševič and Sudar, Reference Uroševič and Sudar1991); NE Spain (Márquez-Aliaga et al., Reference Márquez-Aliaga, Valenzuela-Rios, Calvet and Budurov2000).
Remarks
This subspecies corresponds to the holotype of the species and other elements originally illustrated by Budurov and Stefanov (Reference Budurov and Stefanov1972), most of which have an inturned posterior platform. The posterior platform is variably developed in Nevadan material, but the presence of a distinct secondary lobe or process is regarded as diagnostic. Although typically offset posteriorly, relatively straight elements occur rarely in the Nevadan material. Mature specimens of N. e. excentrica have long rectangular platforms comparable to that of associated N. cornuta. Two much smaller elements with a strongly deflected posterior platform (Fig. 10.1) occur earlier than typical specimens and are given an aff. designation.
Neogondolella excentrica primitiva new subspecies
Figure 11.1–11.11, 11.12?–11.14?
- ? Reference Rafek1976
Neogondolella prava (Kozur); Rafek, pl. IV, figs. 28, 29.
- Reference Nicora1976
Neogondolella excentrica; Nicora, p. 639–640, pl. 84, figs. 1a, b, 2a, b.
- ? Reference Kozur and Mirăuța1980
Gondolella transita Kozur and Mirăuța, taf. 1, figs. 1a–d.
- ? Reference Bagnoli1982
Gondolella cf. transita Kozur and Mostler; Bagnoli, p. 7, pl. 1, figs. 4a, b.
- Reference Uroševič and Sudar1991
Neogondolella excentrica Budurov and Stepanov; Uroševič and Sudar, pl. 1, fig. 15.
- ? Reference Kozur, Krainer and Mostler1994b
Neogondolella sp. aff. N. transita (Kozur and Mostler); Kozur et al., pl. 3, figs. 21, 24.
- p Reference Wu, Yao, Ji and Wang2008
Neogondolella constricta (Mosher and Clark); Wu et al., pl. II, figs. 12, 15.
- p? Reference Wu, Yao, Ji and Wang2008
Neogondolella mombergensis (Mosher and Clark) (sic); Wu et al., pl. I, fig. 13.
- p? Reference Golding2014
Neogondolella ex gr. constricta, morphotype epsilon; Golding, fig. 2.26, parts 1–3.
- Reference Sun, Jiang, Sun and Hao2014
Neogondolella excentrica; Sun et al., fig. 4e.

Figure 11. (1–11) Neogondolella excentrica primitiva n. subsp. (1–7, 9–11), from Prida Formation, Nevada. (1, 2) GSC 141880, sample FH8, Rotelliformis Zone/ burckhardti Subzone; (3, 4) GSC 141881, sample FH44, Meeki Zone/dunni Subzone; (5–7) GSC 141882, holotype, FH6, Rotelliformis Zone/burckhardti Subzone; (8) GSC 141883, sample 92AH-2, Deleeni Zone; (9–11) GSC 141884, sample FH48, Occidentalis Zone; (12–14) Neogondolella excentrica aff. primitiva n. subsp. GSC 141885, sample AH26, Minor Zone. Scale bar = 200 μm.
Holotype
GSC 141882 (Fig. 11.5–11.7), from the Fossil Hill Member of the Prida Formation, burckhardti Subzone of the Rotelliformis Zone (sample FH-6), Fossil Hill, Nevada.
Diagnosis
This is a relatively short subspecies of N. excentrica with a length: breadth 4.5: 1; a short, obliquely directed, posteriorly rounded postero-lateral platform lobe; a relatively low blade/carina in front of the 1 or 2 larger offset terminal denticles; and a weakly extended, asymmetrical basal scar posterior of the pit on the lower side.
Occurrence
In Nevada, about 35 elements are assigned to Neogondolella excentrica primitiva n. subsp., mostly from the Rotelliformis Zone, with rare elements occurring in the Meeki Zone at Fossil Hill and in the Tobin Range. Nicora (Reference Nicora1976) also illustrated the subspecies from the Star Canyon section in Nevada. In B.C., the subspecies occurs in the Deleeni Zone at Yellow Bluffs. A specimen assigned to N. e. aff. primitiva n. subsp. occurs in the middle Anisian Minor Zone along the Alaska Highway (Fig. 11.12–11.14).
Kozur et al. (Reference Kozur, Krainer and Mostler1994b) noted forms similar to this subspecies occurring in the Reitziites reitzi Zone in Austria, and additional records may occur in the Upper Muschelkalk of northern Germany (Rafek, Reference Rafek1976); in the northern Apuseni Mountains in Romania; in Punta Bianca, NW Italy (Bagnoli, Reference Bagnoli1982); eastern Serbia (Uroševič and Sudar, Reference Uroševič and Sudar1991); in the Qingyan Formation in Guizhou Province (Wu et al., Reference Wu, Yao, Ji and Wang2008); and the Upper Guanling Formation in Guizhou Province, China (Sun et al., Reference Sun, Jiang, Sun and Hao2014).
Etymology
From Latin prīmitīvus, signifying the first representative of the species.
Comparisons
These elements share relative platform dimensions with Neogondolella constricta but differ from that species in their asymmetric posterior platforms. They differ from the nominate subspecies in its comparatively shorter length, lower posterior carina height, shorter basal scar, and the rounded posterior platform lobe. The latter margin is like that of N. e. sigmoidalis n. subsp., but both the entire element and the posterior basal keel of Neogondolella excentrica primitiva n. subsp. is much shorter. Kozur et al. (Reference Kozur, Krainer and Mostler1994b) assigned similar forms to N. sp. aff. transita, although only a posterior fragment was illustrated. An older specimen assigned to N. e. aff. primitiva n. subsp. differs from these elements in possessing a slightly higher blade–carina.
Remarks
The asymmetric posterior platform of Neogondolella excentrica primitiva n. subsp. is generally manifest as a deflected rounded lobe that may appear as a secondary process, as in the holotype, or simply as a posterior-lateral expansion with an incurved posterior carina. Commonly there is an additional offset posteriormost denticle like that seen in younger and longer elements assigned to Neogondolella aff. N. cornuta.
Neogondolella excentrica sigmoidalis new subspecies
Figure 10.8–10.15
- Reference Rafek1976
Neogondolella excentrica Budurov and Stefanov; Rafek, pl. II, figs. 1–3.
- Reference Chhabra and Kumar1992
Neogondolella excentrica; Chhabra and Kumar, pl. 3, figs. 8a–c, 9a–c.
- p Reference Kozur, Krainer and Lutz1994a
Neogondolella cf. aequidentata Kozur et al., pl. 3, fig. 2a–c.
- p Reference Kovács1994
Gondolella transita; Kovács, p. 487, taf. 3, figs. 1, 3 (only).
- p Reference Muttoni, Nicora, Brack and Kent2004
Neogondolella bakalovi Budurov and Stefanov; Muttoni et al., pl. 1, figs. 5a–c.
- p Reference Golding2014
Neogondolella ex gr. transita morphotype alpha; Golding, fig. 2.28, parts 1–9.
Holotype
GSC 141877 (Fig. 10.8–10.10), from the upper member of the Prida Formation, upper Subasperum Zone (sample 512), Saurian Hill, Nevada.
Diagnosis
A subspecies of Neogondolella excentrica with an elongate, weakly arched platform of largely uniform width and a length:breadth ratio of 5.5:1. In upper view, the posteriormost one-fifth of the platform has a sigmoidal outline, is inturned, and terminates in a rounded posterior margin. A low blade/carina has relatively discrete anterior denticles, partly fused medial carina, and several large and discrete denticles posterior of the cusp. The basal scar extends beneath the posterior inturned platform far posterior of the pit.
Occurrence
This relatively uncommon subspecies is represented by about 12 elements from the Subasperum Zone at Saurian Hill in Nevada, and the Matutinum and Poseidon zones at Wapiti Lake, Oyster Springs, and Tuchodi Bluffs in B.C. Specimens from the Chischa Zone have less-developed keels, whereas typical specimens occur with ammonoids of the bamberi Fauna.
Other records are those of the ?Upper Muschelkalk of northern Germany (Rafek, Reference Rafek1976); the Curionii Zone at Felsőörs in Hungary (Kovács, Reference Kovács1994) and in the Belvedere section in the Italian Dolomites (Muttoni et al., Reference Muttoni, Nicora, Brack and Kent2004); the Carnic Alps in Austria (Kozur et al., Reference Kozur, Krainer and Lutz1994a); and the Kalapani Limestone, northern India (Chhabra and Kumar, Reference Chhabra and Kumar1992).
Etymology
From the Greek word sigmoeidḗs, shaped like the letter sigma.
Remarks
The long, slender platform and extended keel differentiates Neogondolella e. sigmoidalis n. subsp. from Neogondolella e. primitiva n. subsp., while the rounded posterior platform distinguishes it from N. e. excentrica. The specimen illustrated by Kozur et al. (Reference Kozur, Krainer and Lutz1994a) differs from typical N. aequidentata Kozur, Krainer, and Lutz, Reference Kozur, Krainer and Lutz1994a, in its asymmetry and lower, less arched carina.
In lower Ladinian samples from B.C., elements of Neogondolella posterolonga occur in which posterior platform asymmetry occurs without an extended keel on the underside. These appear to be transitional to Neogondolella e. sigmoidalis n. subsp., which shares the same relative platform dimensions. This mirrors the intermediate nature of N. aff. N. cornuta between N. cornuta and N. e. excentrica, and the variants included in N. e. primitiva n. subsp.
Neogondolella ex gr. mesotriassica (Kozur and Mostler, Reference Kozur and Mostler1982)
Figure 12
- p* Reference Kozur and Mostler1982
Gondolella mesotriassica Kozur and Mostler, p. 293–294, pl. 1, figs. ?2, 3, 4.
- p Reference Kozur, Krainer and Mostler1994b
Neogondolella mesotriassica; Kozur et al., pl. 4, figs. 7–9 (only).
- Reference Budurov and Stefanov2014
Neogonolella constricta longa Budurov and Stafanov; Sun et al., fig. 4d.

Figure 12. Neogondolella ex gr. mesotriassica (Kozur and Mostler) from (1–3) Toad Formation, B.C. and (4–9) Prida Formation, Nevada. (1–3) GSC 141898, sample 83/215F, Toad River Canyon, Minor–Deleeni zonal boundary; (4–6) GSC 141899, sample FH13, Rotelliformis Zone/clarkei Subzone; (7–9) GSC 141900, sample FH48, Occidentalis Zone/hyatti Subzone. Scale bar = 200 μm.
Holotype
Kozur and Mostler, pl. 1, fig. 4, sample BM 44/75, from within the Reiflingen limestones at Buchberg, near Göstling, lower Austria.
Occurrence
Slightly different specimens assigned to this group occur sporadically through the upper Anisian in Nevada, largely in the Rotelliformis Zone, but also in younger strata. In B.C., the occurrence of this species is bracketed by the Minor and Deleeni zones on Toad River. The characteristic form starts at the base of the Reitzi Zone in Austria (Kozur et al., Reference Kozur, Krainer and Mostler1994b). A similar element was illustrated from the Upper Guanling Formation in Guizhou Province, China (Sun et al., Reference Sun, Jiang, Sun and Hao2014).
Description
The segminiplanate P1 elements are arched, with a subrectangular platform with a squared-off or truncated posterior margin that may have a v-shaped notch visible in upper view. The platform is slightly broader at midlength and tapers in both directions before often broadening at the posterior end. The blade denticles are more discrete than the largely fused median carina, and together they impart an arcuate upper profile to the element. The carina ends in a small terminal denticle that is separated from those to the anterior, including the cusp. Additional denticles commonly are developed on the posterior platform margin. The basal pit is subterminal in the keel, with an asymmetrical basal loop that reflects the lateral expansion of the posterior end.
Remarks
The holotype of Neogondolella mesotriassica has a v-shaped notch on the posterior margin, but it has neither a truncated posterior platform, nor accessory posterior denticles, as cited in the original diagnosis. The terminal denticle of the holotype is discrete and quite unlike those of the two paratypes, which have the final denticle fused with the posterior platform margin. The latter feature led Kovács et al. (Reference Kovács, Nicora, Szabo and Balini1990, p. 171) to suggest that one paratype was an example of the gamma morphotype of N. constricta, later named N. postcornuta (Kovács, Reference Kovács1994). On the contrary, Kozur et al. (Reference Kozur, Krainer and Mostler1994b) regarded most examples of the latter species as examples of N. mesotriassica, but did not provide a formal taxonomic revision. Both species are retained here based on the divergent morphology of the holotypes.
These elements resemble Paragondolella bifurcata, which is also characterized by a squared-off posterior margin and accessory denticles, but that species differs in its high carina and blade. Neogondolella quasiconstricta n. sp. has similar features but is much broader posteriorly.
Uncommon examples of Neogondolella mesotriassica from both Nevada and B.C. differ in detail but are united by the narrow, truncated, and denticulate character of the posterior margin. The morphology of the holotype and the diversity in the current material suggest further revision will be necessary within this group. The three examples illustrated are from progressively younger horizons and appear to lengthen and show anterior pit migration, which is the same trend recognized in other members of the constricta group.
Neogondolella postcornuta (Kovács, Reference Kovács1994)
Figure 13
- Reference Trammer1975
Gondolella longa; Trammer, pl. 23, figs. 2a, b, 3a, b.
- p Reference Kovács, Nicora, Szabo and Balini1990
Gondolella constricta morphotype γ Kovács et al., p. 188, pl. 1, fig. 4a–c; pl. 3, fig. 5.
- * Reference Kovács1994
Gondolella constricta postcornuta Kovács, p. 484, pl. 1, figs. 4, 5; pl. 2, figs. 2–5; pl. 6, fig. 4 (see synonymy).

Figure 13. Neogondolella postcornuta (Kovács) from Prida Formation, Fossil Hill, Nevada. (1, 2) GSC 141901, sample FH35, Meeki Zone/nevadanus Subzone; (3, 4) GSC 141902, sample FH47, Meeki–Occidentalis zonal boundary; (5–7) GSC 141903, sample FH8, Rotelliformis Zone/burckhardti Subzone; (8–10) GSC 141904, sample FH50, Occidentalis Zone. Scale bar = 200 μm.
Holotype
Kovács, p. 484, pl. 1, fig. 4a–d, catalogue number T-6451, sample No. 4, Öskü road-side section, Balaton Highland, Hungary, reposited in the Museum of the Hungarian Geological Survey (Kovács et al., Reference Kovács, Nicora, Szabo and Balini1990, fig. 10). Lower Curionii Zone, Vászoly Limestone Member of the Buchenstein Formation.
Occurrence
Similar specimens occur throughout upper Anisian and into Ladinian strata at Fossil Hill, Nevada, but their occurrence is sporadic because they are not always distinguishable from some growth stages of N. cornuta. The species has not been recognized in B.C.
In the Balaton Highland, Hungary, Neogondolella postcornuta occurs from the uppermost part of the meriani B horizon to the lower Curionii Zone. The first primitive representatives of the species occur in the uppermost bed of the meriani B horizon, whereas common typical ones occur in the liepoldti horizon (Kovács, Reference Kovács1994). A very similar element to the largest illustrated here was figured from southwestern Holy Cross Mountains, Poland (Trammer, Reference Trammer1975). Similar transitional forms occur in the upper Curionii Zone in the Vászoly and Felsőörs sections (Kovács, Reference Kovács1994).
Description
The segminiplanate P1 element is relatively narrow, elongate, and slightly arched with a posteriorly inclined main denticle completely fused with the posterior platform end. The platform is narrowly biconvex for the most part but narrows in its posterior part. The cusp is inconspicuous in the low carina that passes anteriorly into higher and less fused denticles. The small basal pit is subterminal in position.
Comparisons
According to Kovács (Reference Kovács1994), Neogondolella postcornuta differs from intermediate-sized elements of its forebear Neogondolella cornuta by the lack of a conspicuous cusp anterior of the last denticle of the carina, and by the complete fusion of the terminal denticle to the platform end. However, Kovács (Reference Kovács1994) stressed that these distinguishing characteristics applied chiefly to elements of intermediate size, and both early juvenile and hyperadult growth stages were less distinctive. The Nevadan specimens of this species do not exhibit the subparallel lateral platform margins and truncated posterior margin seen in N. cornuta.
As discussed by Kovács (Reference Kovács1994), N. postcornuta gives rise to N. bakalovi, a Balkan species characterized by a strongly extended posterior platform. Although rare specimens from Nevada show a similar feature, typical elements of the latter species are not recorded in North America. Specimens of N. posterolonga have a similar platform shape but are narrower, longer, and lack the fused posterior denticle.
Remarks
Elements exhibiting the characteristic posterior configuration of Neogondolella postcornuta occur sporadically through the range of N. cornuta, and it is difficult to separate many smaller specimens in which a terminal denticle is located on the platform margin. Chen et al. (Reference Chen, Krystyn, Orchard, Lai and Richoz2015) provided a broad review of Middle Triassic conodonts and suggested N. postcornuta was in fact a junior synonym of N. pseudolonga. As discussed previously, N. pseudolonga is here regarded as a probable synonym of N. cornuta or N. posterolonga. This emphasizes the difficulty of interpreting different growth stages of these allied species.
Neogondolella posterolonga Kozur, Krainer, and Mostler, Reference Kozur, Krainer and Mostler1994b
Figure 14
- p Reference Rafek1976
Neogondolella navicula (Huckriede); Rafek, Pl. III, fig. 7 (only).
- ? Reference Bagnoli1982
Gondolella pseudolonga Kovács, Nicora, and Mietto; Bagnoli, p. 6, pl. 1, figs. 5a, b.
- p Reference Nicora and Kovács1984
Gondolella constricta Mosher and Clark; Nicora and Kovács, p. 144–148, pl. 8, fig. 3.
- p Reference Nicora and Kovács1984
Gondolella mombergensis mombergensis Huckriede; Nicora and Kovács, p. 149–150, pl. 9, figs. ?7, ?10.
- p Reference Nicora and Kovács1984
Gondolella mombergensis longa (Budurov and Stefanov); Nicora and Kovács, p. 150, pl. 10, figs. 6, 7.
- * Reference Kozur, Krainer and Mostler1994b
Neogondolella aldae posterolonga Kozur et al., p. 182–183.
- Reference Ramovš and Goričan1995
Neogondolella constricta; Ramovš and Goričan, pl. 9, fig. 3a, b.
- Reference Pevný and Salaj1997
Gondolella constricta; Pevný and Salaj, pl. XI, figs. 2, 3.
- ? Reference Orchard, Cordey, Rui, Bamber, Mamet, Struik, Sano and Taylor2001
Neogondolella ex gr. constricta; Orchard et al., pl. 1, fig. 18.
- ? Reference Sano and Orchard2004
Neogondolella ex gr. constricta; Sano and Orchard, fig. 6.13, 6.14.
- p Reference Golding2014
Neogondolella transita morphotype beta; Golding, fig. 2.29, parts 4–6.
- p Reference Golding and Orchard2021
Neogondolella ex gr. constricta; Golding and Orchard, p. 29, pl. 15, figs. 1–4.
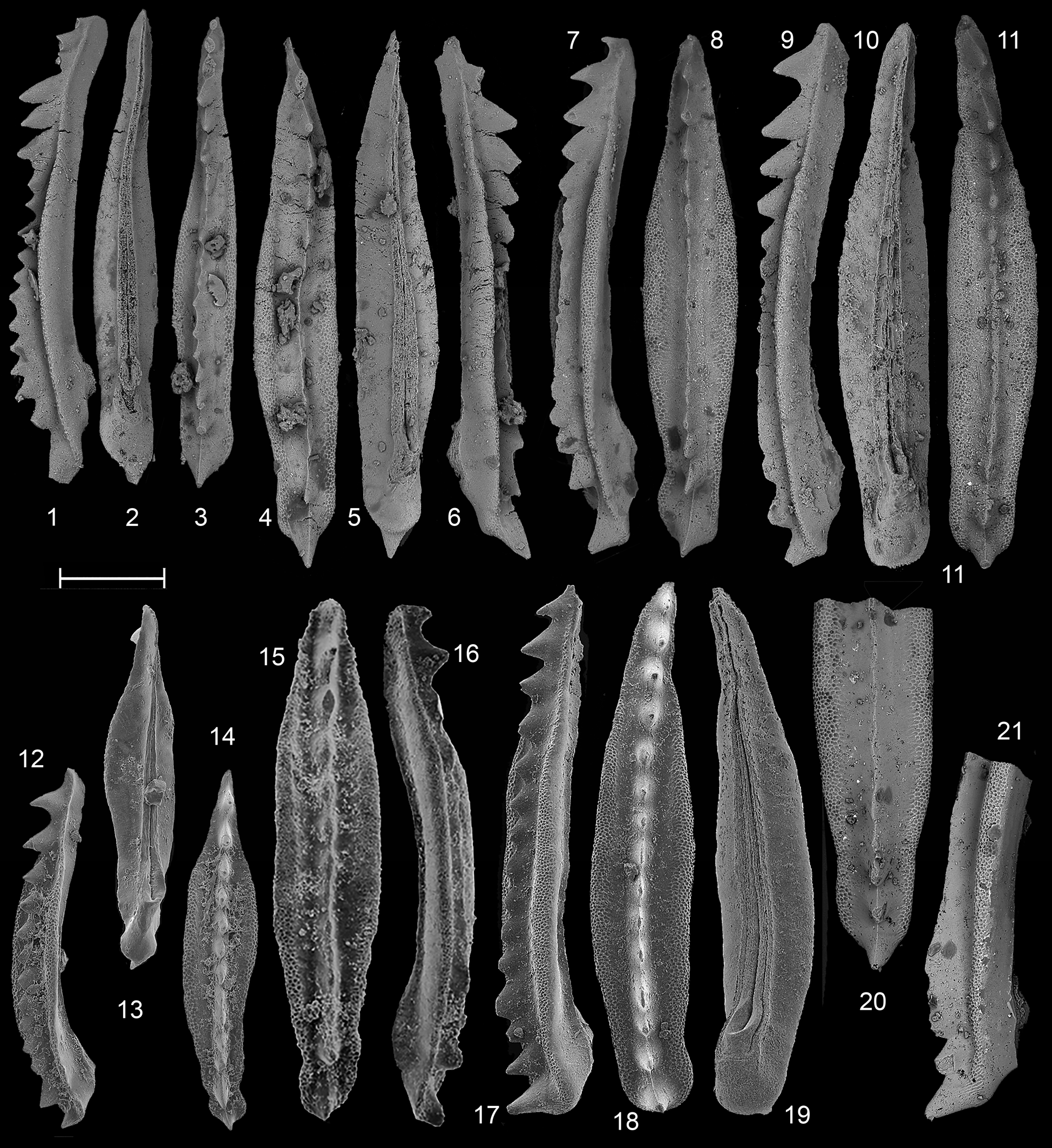
Figure 14. Neogondolella posterolonga Kozur, Krainer and Mostler from (1–11) Prida Formation, Nevada, Subasperum Zone, and (12–21) from B.C. (1–3) GSC 141890; (4–6) GSC 141891, both from sample SH520; (7, 8) GSC 141892; (9–11) GSC 141893, both from Subasperum Zone SH534; (12–14) GSC 141894, sample 97/WapA6, Llama Formation, Poseidon? Zone; (15, 16) GSC 141895, sample 82/AH1, Liard Formation, near milepost 386, Alaska Highway, 2 m below Meginae Zone; (17–19) GSC 141896, from sample GSC cur. No. O-83862, Llama Formation, Wapiti Lake, Matutinum Zone; (20, 21) GSC 141897, sample 01/URC9, Toad Formation, Ursula Creek, Williston Lake, 95 m above the Permian–Triassic Boundary. Scale bar = 200 μm.
Holotype
The specimen figured by Nicora and Kovács (Reference Nicora and Kovács1984, pl. 10, fig. 7) as N. mombergensis longa, from sample N50, in the middle Prida Formation, lower Occidentalis Zone age at Fossil Hill, Humboldt Range, Nevada.
Occurrence
Typical representatives of Neogondolella posterolonga appear in the Meeki Zone and form a larger component of the faunas in the Occidentalis and Subasperum zones in Nevada. Kozur et al. (Reference Kozur, Krainer and Mostler1994b) recorded first occurrences in the upper Meeki Zone, but some early growth stages of constricta group elements from the Rotelliformis Zone are similar. Elsewhere in Nevada, the species is known from Wheeler Mine, and in the Tobin Range. In B.C., examples occur in the Chischa Zone at Yellow Bluff; the Matutinum Zone at Wapiti Lake; the Poseidon Zone at Ganoid Ridge and Tuchodi Bluff; and the Meginae Zone along the Alaska Highway at milepost 386 (Table 2).
The specimen from Hungary comes from the Curionii Zone (Kovács, Reference Kovács1994). The species is also recorded in the southwestern Holy Cross Mountains, Poland (Trammer, Reference Trammer1975); Upper Muschelkalk, northern Germany (Rafek, Reference Rafek1976); ?Punta Bianca, NW Italy (Bagnoli, Reference Bagnoli1982); central Slovenia (Ramovš and Goričan, Reference Ramovš and Goričan1995); western Carpathians, Slovakia (Pevný and Salaj, Reference Pevný and Salaj1997); and ?Necoslie Breccia, Cache Creek terrane, Nechako (Orchard et al., Reference Orchard, Cordey, Rui, Bamber, Mamet, Struik, Sano and Taylor2001; Sano and Orchard, Reference Sano and Orchard2004; Golding and Orchard, Reference Golding and Orchard2021).
Description
The platform of this arched segminiplanate element is slender, widest anterior of unit mid-length, and relatively long and narrow for the posterior one-fourth to one-third of its length. Typical length:breadth ratios are 5.0–5.5:1. The blade carina is like other constricta group taxa, with more discrete anterior denticles, a lower, largely fused medial carina in later growth stages, and higher, more discrete posterior denticles, including an often-enlarged cusp and a prominent weakly to strongly inclined terminal denticle around which a very narrow platform brim may be developed. On the underside, there is a short, often irregular posterior extension of the basal loop.
Remarks
Kozur et al. (Reference Kozur, Krainer and Mostler1994b) originally included Neogondolella posterolonga as a subspecies of N. aldae, but it is here elevated to a species. In contrast to the latter, N. posterolonga has a much longer and narrower, less biconvex platform. In N. constricta and N. cornuta, the constriction disappears during later growth whereas in the present species the posterior platform is noticeably narrower. In this sense, the species is considered to retain a juvenile morphology in adult specimens.
Neogondolella posterolonga has been confused with N. longa, but that species is longer, narrower (length:breadth ratio ~6:1), and of more uniform width and with more discrete denticles. Neogondolella pseudolonga was regarded as an early growth stage of N. longa by Kozur et al. (Reference Kozur, Krainer and Mostler1994b) but it may be related to the present species or to N. cornuta. Both N. postcornuta and N. bakalovi may have a similar platform shape, but they have a terminal denticle fused with the platform margin.
Neogondolella quasiconstricta new species
Figure 15.1–15.7
- p Reference Szabo, Kovács, Lelkes and Oravecz-Scheffer1980
Gondolella prava Kozur; Szabo et al., pl. 59, fig. 12 (only)
- ? Reference Pisa, Perri and Veneri1980
Neogondolella bifurcata Budurov and Stefanov; Pisa et al., p. 817, pl. 80, figs. 5a, b, 6a–c, 7a, b.
- Reference Sudar1982
‘Gondolella’ bifurcata (Budurov and Stefanov); Sudar, pl. II, fig. 6; pl. III, fig. 1.
- ? Reference Farabegoli, Levanti, Perri and Veneri1984
‘Gondolella’ bifurcata; Farabegoli et al., fig. 5.d1–d3, e1, e2.
- Reference Kovács, Nicora, Szabo and Balini1990
Neogondolella bifurcata bifurcata; Kovács et al., pl. 1, fig. 2.
- ?p Reference Kovács and Vörös2003
‘Gondolella’ bifurcata; Kovács, pl. C-IV, figs. 3–5; C-VII, figs. 1, 2 (only).
- ?p Reference Kovács and Rálisch-Felgenhauer2005
Paragondolella bifurcata; Kovács and Rálisch-Felgenhauer, pl. XIV, fig. 2a–e; pl. XVI, fig. 1a–c.
- ? Reference Wu, Yao, Ji and Wang2008
Neogondolella bifurcate (sic); Wu et al., pl. I, figs. 4–6, 11, 12, 18, 19.
- ? Reference Golding2014
Neogondolella constricta morphotype zeta; Golding, fig. 2.23, parts 1–6.
Holotype
GSC 141906 (Fig. 15.4–15.6), from the Fossil Hill Member of the Prida Formation, vogdesi Subzone of the Rotelliformis Zone (sample FH17), Fossil Hill, Nevada.
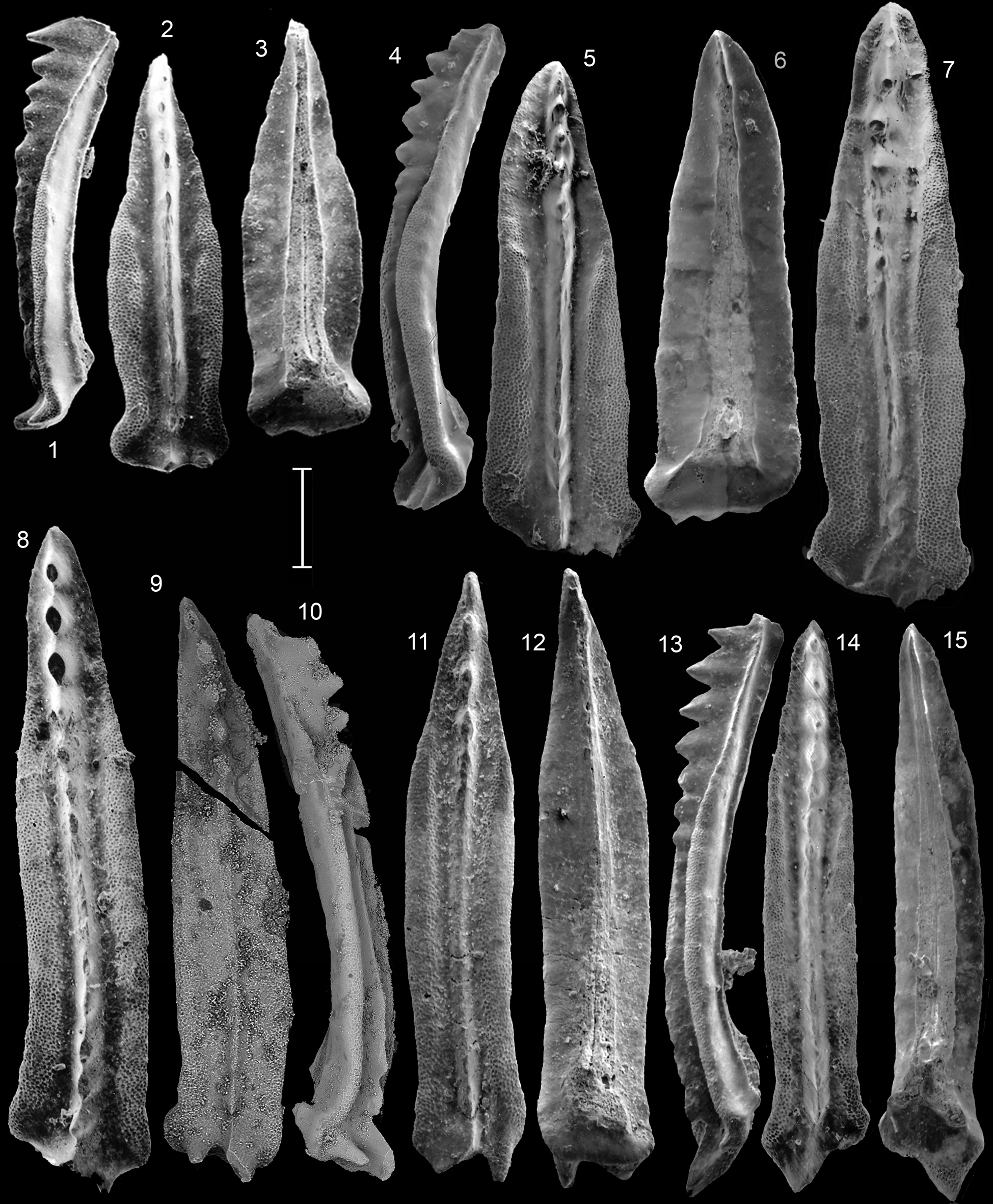
Figure 15. (1–7) Neogondolella quasiconstricta n. sp. from Prida Formation, Nevada. (1–3) GSC 141905, sample FH6, Rotelliformis Zone/burckhardti Subzone; (4–6) GSC 141906, holotype, sample FH17, Rotelliformis Zone/vogdesi Subzone; (7) GSC 141907, sample FH24, Rotelliformis Zone/cricki Subzone. (8–15) Neogondolella quasicornuta n. sp. from Prida Formation, Nevada and (9, 10) Liard Formation, B.C. (8) GSC 141908, sample SH524, Occidentalis Zone; (9, 10) GSC 141909, sample 92-AH-3, Alaska Highway, bamberi Fauna; (11, 12) GSC 141910, sample SH504, Subasperum Zone; (13–15) GSC 141911, holotype, sample SH519, Occidentalis Zone. Scale bar = 200 μm.
Diagnosis
The platform of the comparatively large P1 element has a length:breadth ratio of 3.5–4:1, an expanded postero-lateral margin, and commonly accessory denticles on one or both sides of the prominent terminal denticle. The blade carina is low throughout, with more discrete blade denticles passing into a largely fused median carina. The pit is subterminal within the triangular keel.
Occurrence
Neogondolella quasiconstricta n. sp. occurs principally in the Rotelliformis Zone in Nevada, always in association with, but as a minor component of abundant Neogondolella constricta collections. Younger occurrences are far less common. Elsewhere in Nevada, the species is known from the Tobin Range, and in Canada it occurs in the late Anisian of North Tetsa Phosphate section on the Alaska Highway. About 25 elements were assigned to this species.
The species is also recognized from the base of the Reitzi Zone in Felsőörs, Hungary (Szabo et al., Reference Szabo, Kovács, Lelkes and Oravecz-Scheffer1980; Kovács, Reference Kovács and Vörös2003); Han–Bulog Limestone, SE Bosnia (Sudar, Reference Sudar1982); ?middle Bivera Formation, southern Alps, northern Italy (Farabegoli et al., Reference Farabegoli, Levanti, Perri and Veneri1984), and southern Alps (Pisa et al., Reference Pisa, Perri and Veneri1980). Some elements from the the Qingyan Formation in Guizhou Province, China (Wu et al., Reference Wu, Yao, Ji and Wang2008) could be representatives of this species, although crucial lateral views were not provided.
Description
The large segminiplanate P1 element has a platform with a length:breadth ratio of 3.5–4:1 and an expanded posterior end with a subtriangular outline; the median platform is generally narrower but may be biconvex. The blade–carina is low throughout, with 4–5 discrete anterior blade denticles rising above the downturned anterior platform, and a largely fused median carina. The terminal posterior denticle is prominent and is commonly accompanied by discrete accessory denticles on one or both sides of it. The pit is subterminal within the basal scar that expands progressively to the posterior where the triangular outline mirrors that of the platform.
Etymology
Combination of the Latin quasi- as if, and constricta, the often-co-occurring species that shares the same platform length:breadth ratio.
Comparisons
Neogondolella quasiconstricta n. sp. is similar to, and has often been combined with Paragondolella bifurcata, which differs in having a distinctly high blade and carina, as is characteristic of the genus and the associated Pelsonian species, P. bulgarica and P. hanbulogi. Gondolella fueloepi pseudobifurcata Kovács, Reference Kovács1994, from the costatus horizon of the Reitzi Zone resembles Neogondolella quasiconstricta n. sp. but it has a distinctive posterior platform brim.
Remarks
Typical specimens of P. bifurcata illustrating the diagnostic blade–carina feature were figured by Budurov (Reference Budurov1980) and Qin et al. (Reference Qin, Golding, Jiang, Chen, Zhang, Kang, Wang and Yuan2021). On the contrary, many specimens assigned to P. bifurcata have a low carina (see synonymy), and although this may result from allometric growth (Chen et al., Reference Chen, Neubauer, Krystyn and Richoz2016), there are no high-bladed elements with this platform configuration in the Nevadan material.
Muschelkalk neogondolellins with a single secondary carina developed as nodes posterior of the cusp were assigned to Gondolella mombergensis prava by Kozur (Reference Kozur1968a, Reference Kozurb), and that designation was adopted by Szabo et al. (Reference Szabo, Kovács, Lelkes and Oravecz-Scheffer1980). Subsequently these forms have generally been assigned to Paragondolella bifurcata (e.g., Kovács, Reference Kovács and Vörös2003), however the holotype of Neogondolella prava (Kozur, Reference Kozur1968a, pl. 1, fig. 2a, b) is clearly allied with its Germanic associate N. mombergensis, as shown by similar arched platforms, blade–carina configuration, and geniculate anterior platform (see refigured holotype in Orchard and Rieber, 1998, fig. 1). Rafek (Reference Rafek1976, fig. 4) illustrated a variety of N. prava morphotypes from the upper Muschelkalk, which correspond to neither P. bifurcata nor N. quasiconstricta n. sp.
Neogondolella quasiconstricta n. sp. often occurs with both Neogondolella constricta and N. excentrica primitiva n. subsp., with which it shares relative platform dimensions and a relatively low blade–carina. The three species differ in their posterior configurations, as do the Pelsonian triad of species mentioned above.
Neogondolella quasicornuta new species
Figure 15.8–15.15
- ? Reference Mietto and Petroni1979
Neogondolella mombergensis prava (Kozur); Mietto and Petroni, p. 10, pl. II, fig. 6a, b.
- p Reference Szabo, Kovács, Lelkes and Oravecz-Scheffer1980
Gondolella prava Kozur; Szabo et al., pl. 59, fig. 13a, b (only).
- Reference Kolar-Jurkovšek1983
Neogondolella bifurcata (Budurov and Stefanov); Kolar-Jurkovšek, p. 336–337, pl. 7, fig. 1a–c (only).
- ?p Reference Kovács and Vörös2003
‘Gondolella’ bifurcata; Kovács, pl. C-IV, fig. 6a, b (only).
Holotype
GSC 141911 (Fig. 15.13–15.15), from the Fossil Hill Member of the Prida Formation, Occidentalis Zone (sample SH519), Saurian Hill, Nevada.
Diagnosis
The P1 platform element is comparatively long and narrow with a length:breadth ratio of 5–6:1, and subparallel lateral margins that broaden near the posterior end. The blade–carina is low throughout, higher in the anterior, fused in the central part, and slightly elevated in the posterior one-third. A prominent terminal denticle is commonly flanked by accessory denticles. The pit is slightly shifted anterior in the keel, which extends beneath the postero-lateral platform expansion posterior of the pit.
Occurrence
Neogondolella quasicornuta n. sp. is uncommon from high in the Meeki Zone through the low Occidentalis Zone and into the Subasperum Zone at Fossil Hill, and in the latter zone at Wheeler Mine, Nevada. About 50 elements are represented within the abundant collections of Neogondolella cornuta from Nevada. The species is not yet identified in B.C.
In Europe, similar elements are known from ?San Ulderico section, NE Italy (Mietto and Petroni, Reference Mietto and Petroni1979) (no upper view); ?Balaton Highland, Hungary (Szabo et al., Reference Szabo, Kovács, Lelkes and Oravecz-Scheffer1980; Kovács, Reference Kovács and Vörös2003) (no lower view); and Idrske Krnice, Slovenia (Kolar-Jurkovšek, Reference Kolar-Jurkovšek1983).
Description
The elongate segminiplanate P1 elements of this species have a length:breadth ratio of 5–6:1. The lateral platform margins are subparallel for much of the element length before broadening near the posterior end. The generally low blade–carina resembles that seen in Neogondolella cornuta with more discrete anterior blade denticles, a fused median carina, and slightly elevated, less-fused posterior carinal denticles. A very prominent terminal denticle is accompanied by one or more accessory denticles on the posterior margin. The pit is slightly shifted anterior in the keel, which extends beneath the postero-lateral platform expansion.
Etymology
Combination of the Latin quasi- as if, and cornuta, the species that co-occurs and shares its platform length:breadth ratio.
Remarks
Neogondolella quasicornuta n. sp. shares relative platform dimensions with both N. cornuta and N. excentrica excentrica, with which it often co-occurs. As in the much shorter N. quasiconstricta n. sp., the three associated species differ in their posterior platform configurations. The specimen from Balaton Highland is relatively longer than any others illustrated from Hungary, but no lower view is shown.
Acknowledgments
The Nevadan samples were collected initially by the senior author with the guidance of H. Bucher, who provided the ammonoid biostratigraphic framework illustrated in Figure 2. Early sampling in western Canada was similarly guided by T. Tozer. B. Nicoll and Tyrell Museum staff facilitated joint fieldwork in the Wapiti Lake area. P. Krauss and H. Taylor provided laboratory support. The work was undertaken during several Geological Survey of Canada projects, the most recent of which were GEM 2 and GEM-GeoNorth. V. Karádi, Z. Lyu, and S. Zhang are thanked for providing helpful reviews.
Declaration of competing interests
The authors declare none.

















