


We recently reported an association of educational attainment with polygenic risk scores (PRS) computed using the 50% of DNA not transmitted to offspring by their parents (Bates et al., Reference Bates, Maher, Medland, McAloney, Wright, Hansell and Gillespie2018), a finding also demonstrated elsewhere (Koellinger & Harden, Reference Koellinger and Harden2018; Kong et al., Reference Kong, Thorleifsson, Frigge, Vilhjalmsson, Young, Thorgeirsson and Stefansson2018). Those findings were based on the second GWAS meta-analysis on educational attainment published by the Social Science Genetic Association Consortium (EA2; Okbay et al., Reference Okbay, Beauchamp, Fontana, Lee, Pers, Rietveld and Benjamin2016). Here, we use the pseudo-control method (Cordell et al., Reference Cordell, Barratt and Clayton2004) and the recently available third phase of educational attainment scores (EA3; Lee et al., Reference Lee, Wedow, Okbay, Kong, Maghzian, Zacher and Cesarini2018) to replicate this finding with a stronger polygenic predictor. We employed the same sample and high-quality measure of educational attainment, testing the replication of the hypotheses that: offspring EA3 PRS would positively predict their educational attainment (H1); parents’ EA3 PRS would positively predict their attained SES (H2); and offspring’s educational attainment would be significantly associated with parental EA3 PRS computed from alleles not transmitted to their offspring (H3). Finally, this cultural transmission effect would operate via socioeconomic competence at creating the extended phenotype of environmental niches aiding offspring development (H4).
Methods
Subjects
The sample consisted of all 2335 adolescent twins and their genotyped parents available within the Brisbane Adolescent Twin Study (Wright & Martin, Reference Wright and Martin2004) for whom educational attainment data and genotype data are available (1333 female, mean age at testing 17.15 years, SD = 0.39; 1002 male, mean age at testing 17.2 years, SD = 0.41), along with genotype and SES data for their parents. Exclusion criteria for entry to the cohort were significant head injury, neurological or psychiatric illness, substance dependence or chronic use of medications with central nervous system effects. Subjects were genotyped on the 610K and GSA Illumina genome-wide SNP platform and imputed to the Haplotype Reference Consortium (version 1.1). The study was approved by the QIMR Berghofer Medical Research Institute’s Human Research Ethics Committee.
Phenotypic Measures
Educational attainment was assessed using the overall score for the Queensland Core Skills Test (QCST; Queensland Studies Authority, 2003): a 7-h comprehensive assessment completed in the final year of high school (age ∼17 years) and providing a global index of achievement, standardized to the state-wide sample means for each year. Childhood SES was assessed using the Australian Socioeconomic Index 2006 (AUSEI06) occupational status scale (McMillan et al., Reference McMillan, Beavis and Jones2009) and was ascertained for each parent ∼12 months prior to the twins taking the QCST. For each twin pair, parental SES was set to the maximum of their maternal and paternal values.
Genetic Measures
Non-transmitted parental genomes were created using PseudoCons (Cordell et al., Reference Cordell, Barratt and Clayton2004; Howey, Reference Howey2014). Polygenic risk scores for EA3 were calculated for the offspring and parental pseudo-control genomes (best-guess imputed genotypes, excluding SNPs with low imputation quality r 2 < .6 and MAF <1%) using PLINK1.9 (Purcell et al., Reference Purcell, Neale, Todd-Brown, Thomas, Ferreira, Bender and Sham2007) and the method described by Wray et al. (Reference Wray, Lee, Mehta, Vinkhuyzen, Dudbridge and Middeldorp2014). A standardized PRS was computed using 79,339 SNPs: all SNPs associated p ≤ .05 recomputed from the cohorts of the third educational attainment GWAS (Lee et al., Reference Lee, Wedow, Okbay, Kong, Maghzian, Zacher and Cesarini2018) leaving out the QIMR and 23andMe samples, and using European population LD information from the 1000 Genomes reference set (pruning criteria LD r 2 < .1 within windows of 10 Mb).
Multilevel modeling was completed using lme from the nlme package (Pinheiro et al., Reference Pinheiro, Bates, DebRoy and Sarkar2017) to construct multilevel models with QCST as the dependent variable, with predictors nested within family and zygosity. Analyses controlled for age at test and for sex.
Results
We tested the replication of the four hypotheses outlined above. Results for hypotheses 1–3 are tabulated in Table 1 for convenience. Replicating hypothesis 1, educational attainment in the offspring was significantly related to their own EA3 PRS in a multi-level model with offspring QCST as the dependent variable, with offspring EA3 PRS, age and sex as predictors (β = 0.223, 95% CI = 0.169, 0.277, t(606) = 8.045, p < .001; see Figure 1, panel A). This exceeds the beta found using EA2 PRS (β = 0.133; 95% CI = 0.078, 0.187, t(606) = 4.805, p < .001). Controlling for parental SES did not materially alter this effect. Updating hypothesis 2, a linear model predicting parental attained SES from parental EA3 PRS was significant (β = 0.29, 95% CI = 0.24, 0.33, t = 11.98, p < .001). This was again larger than was found using EA2 (β = 0.20, 95% CI = 0.15, 0.26). Regarding an ancillary hypothesis that subjects’ EA3 PRS would interact with their parents’ SES (Tucker-Drob & Bates, Reference Tucker-Drob and Bates2016) — that is, a GPRS × SES interaction — no support for this was found (β = −0.02, 95% CI = −0.07, 0.04, t(593) = −0.57, p = .569), a result that comports with twin data from Australia (Bates et al., Reference Bates, Hansell, Martin and Wright2016; Grasby et al., Reference Grasby, Coventry, Byrne and Olson2017).
Table 1. Comparison of Effect Sizes between EA2 and EA3 Polygenic Risk Scores as Predictors of Offspring Educational Attainment and Parental SES

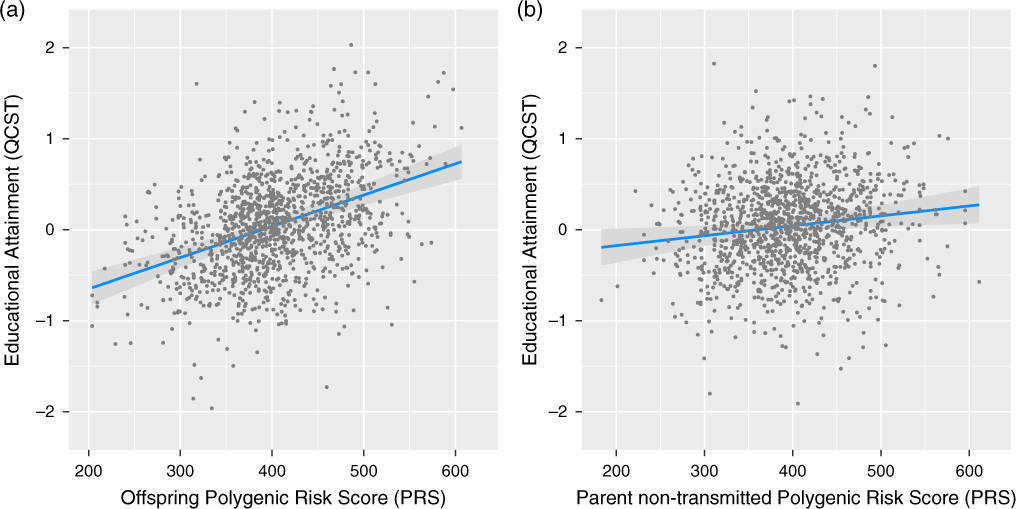
Fig. 1. Prediction of offspring educational attainment from offspring EA3 polygenic risk scores (A) and by parental non-transmitted EA3 polygenic risk scores (B).
Hypothesis 3, that offspring attainment would be significantly associated with parental EA3 PRS computed from alleles not transmitted to their offspring, reflecting cultural transmission, was updated using a multilevel model, with offspring EA3 PRS, parental non-transmitted EA3 PRS and age and sex as predictors. The effect was, again, replicated with increased magnitude (from β = 0.058, 95% CI = 0.002, 0.114 to β = 0.068, 95% CI = 0.014, 0.123, t(605) = 2.45, p = .014; see Figure 1, panel B). Supporting hypothesis 4, which predicts that the non-transmitted effect operates via parental socioeconomic competence, controlling parent SES wiped out the non-transmitted allele effect in the model (β = 0.00, 95% CI = −0.05, 0.06, t(594) = 0.17, p = .862).
Discussion
The findings of our initial investigation were replicated. In addition to increased magnitude of associations of offspring educational attainment with offspring EA3 PRS, the effect of parent genotype on a parental competence phenotype supporting child cognitive development was replicated, with higher attainment in offspring associated with non-transmitted elements of the EA3 PRS, and an effect size around one-third the magnitude of children’s own EA genetic score. The finding supports the view of EA3 gene polymorphisms as underlying a social competence phenotype of which education is just one component. The results also confirmed that the effects of parenting on attainment appear to mediate one or more of the aspects associated with the SES that parents provide their offspring. The useful foci for follow-up research include identifying specific physical and behavioral traits that are causal for improving offspring attainment. Examples of measurable mediators include wealth and housing (Duncan et al., Reference Duncan, Morris and Rodrigues2011; Galobardes et al., Reference Galobardes, Shaw, Lawlor, Lynch and Davey Smith2006), household chaos (Matheny et al., Reference Matheny, Wachs, Ludwig and Phillips1995), as well as parental responsiveness to offspring needs (Bradley & Corwyn, Reference Bradley and Corwyn2002). The method can also be extended to other outcomes, including psychiatric traits in which parental genotypes may be operating as an extended phenotype influencing offspring.
Acknowledgments
We are greatly appreciative of the support from the twins and their families, and their willingness to participate in our study. We thank Marlene Grace, Ann Eldridge and Natalie Garden for participant recruitment and data collection at QIMR Berghofer Medical Research Institute. We also thank the Queensland Curriculum and Assessment Authority for the provision of QCST data and statistics. Any analyses of QCST data constitutes our own assessment of the data and is not a representation of the agency. This collaborative research was supported by the Human Frontier Science Program (grant number RG0154/1998B) and grants from the Australian Research Council (A79600334, A79906588, A79801419, DP0212016, DP0664638, DP1093900) and the Netherlands Organisation for Scientific Research (NWO) 016.Veni.198.058. L.C-C. was supported by a QIMR Berghofer fellowship.



 Open access
Open access