


In spring 1912 a small group of Scottish antiquaries excavated several inhumation graves near Balevullin on the Hebridean island of Tiree (Fig. 1). Although their work was never published, the proximity of these graves to a concentration of Early Iron Age settlement activity (MacKie Reference MacKie1965) led to a suspicion that the Balevullin burials may be of similar date. As a consequence, the sole surviving skeleton was included in a recent programme of AMS dating, targeting human remains associated with Iron Age domestic sites in Scotland (Tucker & Armit Reference Tucker and Armit2009). The results from three radiocarbon determinations demonstrated unequivocally, however, that the burial was of Neolithic date and had nothing to do with the later occupation.

Fig. 1 Location map
The early date was surprising firstly because this unusual individual had apparently suffered from rickets: a childhood disease caused by vitamin D deficiency, and more usually associated with the urban slums of the early modern period than the wide-open spaces of Neolithic Tiree. The disease would have marked this individual as physically different in life. Secondly, despite the profusion of monumental funerary architecture, inhumation cemeteries are virtually unknown in Neolithic Britain, making the Balevullin cemetery a significant and unexpected find. Was it that physical difference in life, in this case bone malformations thought to be associated with rickets, led to this unusual treatment in death?
Discovery and Excavation
Balevullin lies in the north-eastern part of Tiree in an area of coastal machair, light calcareous sands prone to dramatic wind and tidal erosion. The periodic exposure of ancient surfaces attracted antiquarian attention from the late 19th century onwards, most of which focussed on the retrieval of prehistoric artefacts (Beveridge Reference Beveridge1903). The early 20th century excavations which form the subject of this paper were led by A. Henderson Bishop, a wealthy Scottish businessman, pig-breeder and amateur archaeologist, although most of the fieldwork appears to have been carried out by his assistant, Mungo Buchanan; the well-known antiquarian Ludovic Mann appears also to have been involved (Ritchie Reference Ritchie2002, 48).
Bishop’s team concentrated their efforts on an imprecisely located area (NL 959 477) where the machair surface had been disturbed by wind erosion, exposing artefacts and structural remains. Excavation focussed on an Early Iron Age structure (MacKie Reference MacKie1965), but also involved surface collection over the surrounding area. As might be expected in such circumstances, the chipped stone assemblage seems to comprise a chronologically diverse assemblage, including Mesolithic and/or Neolithic material (Pirie Reference Pirie2006, 7).
The discovery of the burials was described in a letter by Bishop written many years later to Anne Robertson of the Hunterian Museum in Glasgow: ‘it was on this sandy area that Mr Ludovic Mann and his sister Mrs Andrew Farquhar, found a stone age burial and in 1913 unearthed the skeleton which Dr Tommy Bryce, then Prof of Anatomy, collared for his museum’ (Bishop Reference Bishop1952).
Although Bishop states that the excavation took place in 1913, Buchanan’s notes made at the time confirm that it actually happened in 1912. ‘Tommy Bryce’s museum’ was the Anatomy Museum at the University of Glasgow, and it was from there that the skeleton (Balevullin 1) was later transferred to the Hunterian Museum, within the same university, to be reunited with the rest of the Bishop Collection. Although Bishop makes no explicit reference to any other skeletons, their fate is implicit in the next section of the letter: ‘These inhumations led to trouble with the Islanders who accused me of digging up their Ancestors and taking them to London to put in a museum. It was reported to His Grace, of Argyle, and we were requested to leave without any remains or other treasures. When the flints and hammer-stones were shown to the local Factor we were absolved and were permitted to take the finds.’
It seems improbable in these circumstances that Bishop was able to remove any further human remains from Tiree. There is no indication of what subsequently became of them, but reburial by the local community seems the most likely outcome.
The Burials
Details of the Balevullin burials can be reconstructed from documents found among Bishop’s papers at the Hunterian Museum. The most relevant are a set of pencil notes by Mungo Buchanan (Reference Buchanan1912) describing the recovery of Balevullin 1 in some detail, with accompanying sketches and photographs (Fig. 2). The notes are dated 30 April 1912, and appear to have been made during or immediately after the excavation. An apparently later (but undated) typescript by Bishop (Reference Bishopnd) repeats much of this material, some of it verbatim, but includes additional information on Balevullin 2. The burials provisionally identified here as Balevullin 3 and 4 are each represented by a single photograph without accompanying text.
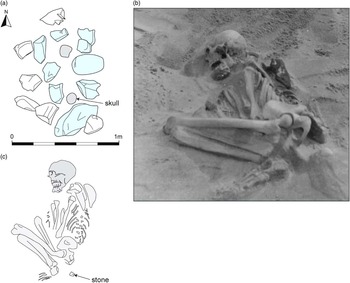
Fig. 2 Balevullin 1. a: the stone setting covering Balevullin 1, with parts of the cranium and pelvis visible at the level of the stones. b: photograph taken during excavation (Bishop Collection, Hunterian Museum). c: sketch plan of the skeleton. The plans have been re–drawn by Rachael Kershaw from annotated pencil sketches by Mungo Buchanan (Reference Buchanan1912)
Balevullin 1
Balevullin 1 lay under a simple stone setting which would have formed a visible, though hardly monumental, above-ground marker (Fig. 2a). Although the spread of stones on Buchanan’s plan measured approximately 1.4 m north–south by 1 m, some of those on the west and north sides may have been displaced from the centre of what was probably a fairly regular rectangular setting over the grave pit, measuring approximately 1.1 m north–south by 0.6 m. This seems unlikely to have been any higher or more substantial originally than it was on its discovery. It is possible that the central stones may have been dislodged by casual visitors to reveal the bones, or even by Mann and his sister when they discovered the site. In either case, the skull and pelvis were already exposed when excavations began. The skeleton was flexed, lying on its right side, with its head to the south (Fig. 2b). The folded position of the left wrist indicates that it may have been broken before or during burial, or else disturbed at a later date.
The only object found within the grave was ‘a white quartzite pebble or chuckie, a little larger than a walnut … about 3” [7.6 cm] from the lower edges of the pelvic bones’ (Bishop Reference Bishopnd). Given the complete absence of stones from the clean sand into which the burial had been dug this seems to be a deliberate inclusion and should probably be regarded as a funerary offering or else as an object belonging to the deceased and perhaps buried in their clothing or wrapping (Fig. 2c).
Three AMS 14C determinations obtained from Balevullin 1 by the Scottish Universities Environmental Research Centre have a combined two sigma range of 3340–3090 cal bc, placing the Balevullin 1 individual firmly within the Neolithic (Table 1).
Table 1 AMS Dating of the Sole Surviving Skeleton (Balevullin 1)

All dates from human bone with calibrated age ranges determined from the University of Oxford Radiocarbon Accelerator Unit calibration programme (Bronk Ramsey Reference Bronk Ramsey2005) using dataset and calibration curve from Reimer et al. (Reference Reimer, Baillie, Bard, Bayliss, Beck, Bertand, Blackwell, Buck, Burr, Cutler, Damon, Edwards, Fairbanks, Friedrich, Guilderson, Hughen, Kromer, McCormac, Manning, Bronk Ramsey, Reimer, Remmele, Southon, Stuiver, Talamo, Taylor, Plicht, van der and Weyenmeyer2004)
Balevullin 2
Records on the second Balevullin burial comprise only a brief description by Bishop (Reference Bishopnd) and a single photograph (Fig. 3a). Any notes or sketches made by Buchanan appear not to have survived. It lay ‘18 yards due south’ of the first and was again flexed on its right side, with the head to the south. Although there is no mention of a stone setting, the preservation of the burial was such that no above-ground marker could have been expected to survive. Indeed, parts of the skeleton, including the cranium, had disappeared, most likely through surface erosion, before the skeleton was excavated. According to Bishop the ‘arms were straight, pointing north …’.
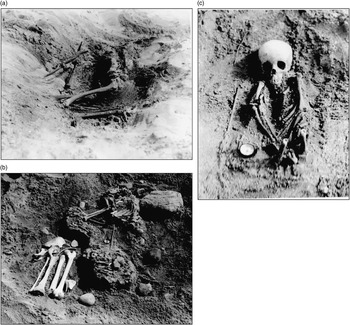
Fig. 3 a: Balevullin 2, b: Balevullin 3, c: Balevullin 4 (courtesy of the Bishop Collection, Hunterian Museum)
Balevullin 3
Alongside the photographs of Balevullin 1 and 2 is a print showing a third flexed inhumation (Fig. 3b). Although unlabelled, the burial environment appears identical to those of the previous two, and the filing of the print alongside the other Balevullin graves strongly implies that this is indeed a third burial from the same site. The lack of any mention in the documentary evidence can probably be explained by the rather abrupt termination of the excavations. Based on the photograph, this individual was apparently an adult, rather more complete than Balevullin 2, in a crouched or flexed position on the right side with the arms flexed in front of the torso. The lower limb bones, unfortunately, were not photographed in situ.
Balevullin 4
Balevullin 4 is again represented by a single photograph (Fig. 3c) without accompanying text, and the same caution must be exercised in its attribution. The skeleton appears to be that of a child between 4 and 7 years of age, lying supine with the arms across the body. Again the lower limb bones appear to have been disturbed: only the distal femora are present, and these point upwards and are broken off halfway along the shafts. It is impossible to determine whether this took place at the time of burial or during excavation.
Other burials
Buchanan’s notes include two sketches of dispersed surface skeletal material which appears also to derive from Balevullin. This suggests that the cemetery may originally have been rather more extensive than the four burials that can be reasonably securely identified with it.
Osteological Analysis
Skeleton 1 is substantially complete (68%) with good surface preservation. Assigning sex proved problematic due to the unusual shape of the pelvis, which is high and narrow, although with an oval aperture and flat sacrum. Several features of the pelvis and skull, however, suggest that this individual was probably female. Age at death was estimated according to the condition of the pubic symphysis and auricular surface (Buikstra & Ubelaker Reference Buikstra and Ubelaker1994; Lovejoy et al. Reference Lovejoy, Owen, Meindl, Pryzbeck and Mensforth1985), and the degree of molar teeth wear (Miles Reference Miles1962), at 25–30 years. Stature was calculated to be 145–150 cm (4’9”–4’11”) based on the length of the single intact femur (Trotter & Gleser Reference Trotter and Gleser1958); notably short even by prehistoric British standards (Roberts & Cox Reference Roberts and Cox2003, 67, 86). No serious dental pathology was noted on the 16 teeth present, although slight calculus and linear dental enamel hypoplasia were present.
The most striking skeletal pathology noted in this skeleton was on the sternum, which exhibits a severe deformation known as pectus carinatum or pigeon chest (Fig. 4a). The facets for the articulating ribs also appear deformed, those for the 3rd–5th ribs being expanded and showing evidence of degeneration in the form of surface pitting, presumably as a result of the sternum deformation. The manubrium and body of the sternum are completely fused, which is unusual in an individual of this age, and most likely a stabilising reaction to the deformation of this bone. Although the 1st and 2nd ribs appeared normal, the 3rd–8th form an unusual angle. The costal ends of the ribs, particularly ribs 5 through to 8, are slightly flared. This deformation is undoubtedly related to that of the sternum, forming the typical ‘pigeon-breast’ deformation of vitamin D deficiency (Ortner Reference Ortner2003, 397, 400). Although the tibiae and fibulae are missing (they are visible in excavation photographs), the humeri do exhibit deformation, with the shafts bowed and rotated (Fig. 4b). The single surviving femur, although not severely deformed, exhibits an unusual angle of the neck (coxa vara) and slight anterior–posterior bowing.
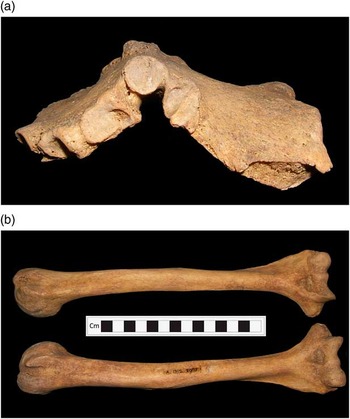
Fig. 4 Balevullin 1. a: sternum, b: humeri (photographs: Fiona Shapland)
The deformation of the sternum and ribs are diagnostic signs of vitamin D deficiency, and the more subtle changes to the humeri and femora support this diagnosis (Brickley & Ives Reference Brickley and Ives2008, 104–5). In particular, the anterior–posterior deformation of the sternum may be compared to classic cases of ‘pigeon-breast’ deformity noted in many rickets cases (Fig. 5). Although similar deformation of the sternum may be caused by genetic conditions such as hypophosphatasia or chondrodystrophy these are very rare, and childhood rickets is considered to be by far the most likely cause (Gupte Reference Gupte2002, 493). Osteomalacia (vitamin D deficiency in adulthood) may produce similar skeletal deformities, but several aspects of this case make rickets more likely. The bones were found to be robust and heavy, whilst active osteomalacia results in lighter, weaker bones (Ortner Reference Ortner2003, 398), and no pseudofractures, a typical sign of osteomalacia (Brickley & Ives Reference Brickley and Ives2008, 127–9), were noted. Some of the particular pathological changes, such as deformation of the humeri, are particularly associated with rickets during infancy (Ortner Reference Ortner2003, 397–9). All of the changes found in this skeleton are typical of residual childhood rickets (Brickley & Ives Reference Brickley and Ives2008, 107–8). Given the relatively young age at death of this individual, it seems most likely that all these deformations took place within the first 18 years of life, and that substantial healing and remodelling took place before death. This suggestion is supported by the stable isotope data (see below). Repeated periods of vitamin D deficiency over early and later childhood are a strong possibility given the different skeletal changes noted.

Fig. 5 An example of sternal deformation caused by vitamin D deficiency (courtesy of the Hunterian Museum at the Royal College of Surgeons, London)
Prior to the identification of the Balevullin example, the earliest cases of rickets identified in Britain did not pre-date the Roman period (Brickley & Ives Reference Brickley and Ives2008, 140), making this individual more than 3000 years earlier than the next dated case. Looking globally, there are a few possible cases from the Russian Palaeolithic, and from the later Neolithic Balkans, South Africa (Brickley & Ives Reference Brickley and Ives2008, 134–5) and Denmark (Brothwell Reference Brothwell1969, 270), but many of these are tentative diagnoses and the Balevullin case remains highly unusual at this early date.
Isotopic Analysis
A series of isotopic analyses were carried out to investigate the diet and potential origins of the Balevullin 1 skeleton, using a second premolar tooth and a sample of rib (Tables 2 and 3). Samples of core enamel, cleaned of all surfaces and adhering dentine, were removed for strontium, oxygen, and carbon isotope analysis following the procedure detailed in Montgomery (Reference Montgomery2002). The remaining tooth was bisected and one half subjected to carbon and nitrogen isotope analysis of incremental dentine sections following the procedure of Beaumont et al. (Reference Beaumont, Gledhill, Lee-Thorp and Montgomery2013) in order to investigate more closely diet over the period of childhood when active rickets was inferred from the osteological evidence. As dentine does not turnover or remodel in the manner of bone, isotopic signals from early childhood are preserved (Beaumont et al. Reference Beaumont, Gledhill, Lee-Thorp and Montgomery2013).
Table 2 Isotopic Analyses of the Balevullin 1 Skeleton

*Measured at University of Bradford; **Measured at NIGL; ‡Calculated using Daux et al. (Reference Daux, Lécuyer, Héran, Amiot, Simon, Fourel, Martineau, Lynnerup, Reychler and Escarguel2008) Eq. 6; #Calculated using the equation of Chenery et al. (Reference Chenery, Pashley, Lamb, Sloane and Evans2012); #Calculated using the equation of Chenery et al. (Reference Chenery, Pashley, Lamb, Sloane and Evans2012); Strontium data are reproduced from Evans et al. (Reference Evans, Chenery and Montgomery2012)
Table 3 Carbon and Nitrogen Isotope Analysis of Incremental Dentine Sections from 2nd Premolar of the Balevullin 1 Skeleton

The isotope ratios of nitrogen, oxygen, and carbon are expressed relative to international standards as δ15N, δ18O, and δ13C and units are per mil (‰). Strontium isotope ratios are expressed as 87Sr/86Sr. Samples were measured at the University of Bradford Stable Light Isotope Laboratory (δ18O and δ13C of enamel carbonate, and δ15N and δ13C of bone and dentine collagen), and the NERC Isotope Geosciences Laboratory (NIGL) at the British Geological Survey, Nottingham (87Sr/86Sr and δ18O enamel phosphate and δ18O and δ13C of enamel carbonate).
Methods
Further details of the methods used are provided in the online supplementary material.
Strontium Isotope Analysis of Enamel
The enamel sample was transferred to the clean laboratories at NIGL where strontium was extracted according to established laboratory procedures (Evans et al. Reference Evans, Stoodley and Chenery2006).
Oxygen and Carbon Isotope Analysis of Enamel
The enamel surface was removed using a diamond dental burr and discarded. Samples of powdered enamel (~5–15 mg) were produced for oxygen isotope analysis of both the carbonate and phosphate fractions at the University of Bradford. The values in Table 2 are the resulting mean values and the analytical error, based on the reproducibility of the standards, was ±0.2‰ (1σ) or better for both δ18Ocarb and δ13Ccarb.
At NIGL, oxygen isotope ratios were produced for both the enamel phosphate and enamel carbonate fractions. A batch control standard (ACC-1, Aldrich) converted to Ag3PO4 had an analytical precision of ±0.12‰ (2σ, n=5).
For enamel carbonate, δ18Ocarb and δ13Ccarb values were measured using a GV Isoprime duel inlet mass spectrometer. In-house carbonate reference material (KCM) was calibrated against NBS19 reference material to normalise the δ18Ocarb values to the VPDB scale. The reproducibility of KCM was ±0.06‰ (1σ; n=9). The resultant δ18Ocarb values were converted to the SMOW scale using Coplen (Reference Coplen1988).
Carbon and Nitrogen Isotope Analysis of Dentine and Bone
Collagen was extracted from dentine and rib bone. Samples were measured at the University of Bradford in duplicate, and laboratory (fish gelatin and bovine liver) and international (IAEA 600, CH6, CH7, N1 and N2) standards were interspersed throughout the run. The analytical error was ±0.2‰ (1σ) or better for both δ13C and δ15N and all samples met accepted quality control parameters for identifying good quality collagen (van Klinken Reference Van Klinken1999). Mean values are presented in Tables 2 and 3.
Results
The strontium isotope ratio obtained from the tooth enamel (0.70934) is a commonly found value in Britain and can indicate food sourced in a region of young Mesozoic or limestone rocks, soils in areas of high rainfall, or coastal regions influenced by sea-splash (Evans et al. Reference Evans, Montgomery, Wildman and Boulton2010). It is not in itself particularly diagnostic. The high strontium concentration of 355 ppm, however, is rare in Britain: concentrations over 300 ppm are almost entirely restricted to humans excavated from machair regions of the Northern and Western Isles (Evans et al. Reference Evans, Chenery and Montgomery2012). This combination of a seawater ratio and high concentration is therefore highly characteristic of the strontium profile of human populations occupying coastal regions or small islands in the North Atlantic irrespective of the geology (Fig. 6). There are few Neolithic individuals from the Western Isles against which to directly compare the Balevullin skeleton but one from Haugabost, Harris, produced a virtually identical combination of strontium isotope ratio and concentration, ie, 0.70934 and 307 ppm. This strontium profile, which is observed in archaeological skeletons in the North Atlantic from the Neolithic through to the medieval period, has been suggested to reflect the environmental and cultural practices of people who inhabit small, maritime islands in the North Atlantic and their consumption of plants grown on soils improved and fertilised with seaweed and subject to persistent salt deposition from sea-splash (Montgomery et al. Reference Montgomery, Evans and Cooper2007a; Montgomery Reference Montgomery2010).
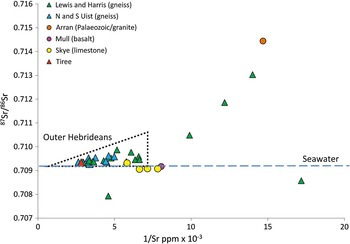
Fig. 6 A plot of enamel strontium isotope and concentration for skeletons excavated from the Inner and Outer Hebrides. The majority of individuals from the Outer Hebrides and the skeleton from Tiree fall within the triangular field: as the strontium concentration increases, the ratio approaches that of seawater. Individuals outside this triangle are deemed to be immigrants to the Outer Hebrides. Analytical uncertainty for 87Sr/86Sr estimated at ±0.002‰ (2σ) is within symbol (data source: Montgomery et al. Reference Montgomery, Evans and Cooper2007a and b; Evans et al. Reference Evans, Chenery and Montgomery2012)
Oxygen isotopes from the carbonate and phosphate fractions of the tooth enamel produced broadly consistent values (17.7‰ for enamel phosphate, 26.3‰ and 26.8‰ for enamel carbonate). Conversion to precipitation of these three results (Table 2) produced a range of values between −6.4‰ and −7.3‰ and highlights the significant uncertainty associated with using such equations and the necessity to be circumspect when interpreting oxygen isotope data (Pollard et al. Reference Pollard, Pellegrini and Lee-Thorp2011). Nonetheless, the measured phosphate value of 17.7‰ is within the range obtained for prehistoric individuals from Skye (Montgomery et al. Reference Montgomery, Evans and Chenery2007b), South Uist (Parker Pearson et al. Reference Parker Pearson, Chamberlain, Craig, Marshall, Mulville, Smith, Chenery, Collins, Cook, Craig, Evans, Hiller, Montgomery, Schwenninger, Taylor and Wess2005), Orkney (Montgomery et al. Reference Montgomery, Schutkowski, Evans and Chenery2007c), Mull, Arran, Lewis, and Harris (Evans et al. Reference Evans, Chenery and Montgomery2012). For example, the Neolithic individual from Haugabost produced an oxygen isotope ratio of 18.0‰ which lies within analytical error of the Balevullin skeleton. The value of 17.7‰ also falls within the mean of 18.2±0.5‰ (1σ) proposed by Evans et al. (Reference Evans, Chenery and Montgomery2012) for human populations inhabiting the higher rainfall (ie western and northern) areas of Britain. The oxygen isotope data are, therefore, consistent with origins on the western or northern seaboard of Britain.
Carbon and nitrogen isotope analysis of the rib (δ13Ccoll=−20.4‰ and δ15N=11.3‰), and carbon isotope analysis of the enamel (mean δ13Ccarb=−14.8‰) indicates a diet based predominantly on C3 plants and protein from terrestrial sources, with little evidence for the consumption of marine resources (Figs 7 & 8). Figure 7 compares the δ13Ccarb value obtained from enamel which represents whole diet (including carbohydrate), with the mean δ13Ccoll value obtained from dentine within the tooth crown (mean crown δ13Ccoll=–20.2‰) which derives largely from consumed protein. These two tissues have overlapping formation times between the approximate ages of 2.5 and 8 years (AlQahtani Reference AlQahtani2009) and together provide strong evidence for a terrestrial diet during this period. The dietary isotopes are thus consistent with the estimated upper δ13Ccoll limit proposed by Bonsall et al. (Reference Bonsall, Cook, Pickard, McSweeney and Bartosiewicz2009) for Neolithic terrestrial diets in the north-west Atlantic region of −20.0‰ and comparable with other Neolithic individuals from the western seaboard (Schulting & Richards Reference Schulting and Richards2002; Jay et al. Reference Jay, Parker Pearson, Richards, Nehlich, Montgomery, Chamberlain and Sheridan2012). Depending on metabolic rates and collagen turnover, the isotope ratios of the ribs of adults may represent an average of several years dietary inputs (Hedges et al. Reference Hedges, Clement, Thomas and O'Connell2007), and would thus not be expected to record any anomalous or sporadic short-term episodes of stress or dietary change during childhood.

Fig. 7 A plot of δ13C values for enamel carbonate (whole diet) against the mean value of co-genetic crown dentine collagen (dietary protein). Errors bars indicate analytical precision of ±0.2‰ (1σ) and are within symbol for enamel carbonate. Regression lines for C3, C4 and marine diets are reproduced from Kellner and Schoeninger (Reference Kellner and Schoeninger2007)
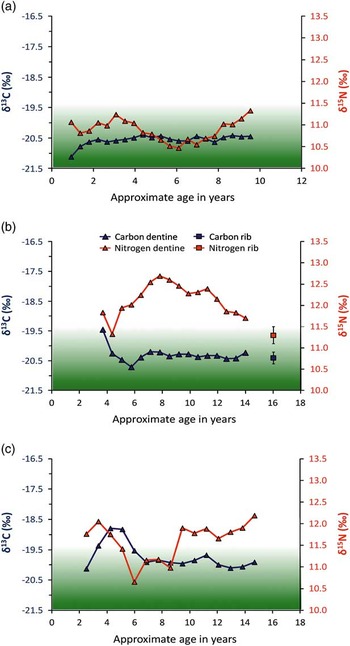
Fig. 8 13C and δ15N life-histories produced from incrementally sampled dentine collagen for: a: SUMB-41, b: Balevullin 1, c: SUMB-44. Profiles a and c, and the shading are redrawn from Montgomery et al. (Reference Montgomery, Beaumont, Jay, Keefe, Gledhill, Cook, Dockrill and Melton2013); the latter estimates the zone for Neolithic terrestrial diets in the Northern and Western Isles
Bulk bone, enamel, or dentine measurements, however, are averaged values over an often uncertain number of years and offer poor temporal resolution (Beaumont et al. Reference Beaumont, Gledhill, Lee-Thorp and Montgomery2013; Montgomery et al. Reference Montgomery, Beaumont, Jay, Keefe, Gledhill, Cook, Dockrill and Melton2013). To investigate whether there was any underlying childhood dietary evidence that might assist in the understanding of the environmental or physiological factors that contributed to the palaeopathological evidence for the Balevullin skeleton, a δ13C and δ15N dietary life-history profile between the ages of 3.5 and 14.5 years was produced from the incremental dentine sections (Fig. 8, plot b). For comparison, childhood dietary profiles from two other adult individuals from the broadly contemporary Neolithic cist at Sumburgh, Shetland Isles (Montgomery et al. Reference Montgomery, Beaumont, Jay, Keefe, Gledhill, Cook, Dockrill and Melton2013) are included in Figure 8 (plots a and c). Although there is little variation in δ13C of the Balevullin skeleton between the ages of 6 years and adulthood (as indicated by the rib value), the δ13C profile falls between the ages of 3.5 and 6 years and, coupled with a similar fall in δ15N from 3.5 to 4 years, this suggests a reduction in marine protein during this period. An alternative interpretation is that the fall in both δ13C and δ15N represents relatively late weaning at around the age of 3.5 years, which is by no means impossible (Stuart-Macadam & Dettwyler Reference Stuart-Macadam and Dettwyler1995). The dentine δ15N values are higher than those from the adult rib sample: mean=12.1±0.4‰ (1σ, n=16) compared to 11.3‰ for the rib (Fig. 8, plot b) and the main trend is a rise from age 4 to around 8 years, followed by a fall until the completion of tooth root formation at around 14.5 years of age (this falling trend continues in the bone sample). If the high δ15N value was the result of a period of marine protein consumption, the δ13C profile would also be expected to rise as can be seen in SUMB-44 between the ages of 2 and 6 years (Fig. 8c). Equally, a Neolithic largely terrestrial diet in Shetland appears to result in both profiles remaining within the shaded zone for SUMB-41 (Fig. 8a). As discussed above, bone collagen values for both δ15N and δ13C would be expected to be within this zone for the majority of Neolithic individuals in Britain consuming a terrestrial-protein diet as for example at the Neolithic site at Carding Mill Bay where individual δ15N bone collagen values do not exceed 10.0‰ and δ13C are below −21.3‰ (Schulting & Richards, Reference Schulting and Richards2002).
Given the consistently flat δ13C profile until the age of 14.5 years, this rise in δ15N may reflect a dietary or physiological change, such as an increased consumption of high-trophic level terrestrial protein (eg, omnivorous rather than herbivorous animal protein or human milk) or nutritional and/or physiological stress (Fuller et al. Reference Fuller, Fuller, Harris and Hedges2006; Beaumont et al. Reference Beaumont, Gledhill, Lee-Thorp and Montgomery2013). The high strontium concentration found in the tooth enamel would perhaps mitigate against a high-meat diet because the amount of strontium in skeletal tissues is lower in carnivores than herbivores in a foodchain due to removal (biopurification) of strontium at each trophic level (Burton et al. Reference Burton, Price and Middleton1999; Burton & Price Reference Burton and Price2000): if an individual was consuming a largely meat-based diet or meat from omnivores or carnivores, they would be expected to have low strontium concentrations. The separation of δ15N from the δ13C profile during the childhood of the Balevullin skeleton is therefore unusual in the context of contemporary individuals from a small, agriculturally marginal NW Atlantic island. A similar divergence of δ15N from δ13C can be seen in the profile of SUMB-44 between the ages of 10 and 15 years and nutritional or health stress was postulated as a cause given that at this age, breastfeeding is not a plausible driver and for the rest of the population δ15N remained within the shaded zone when δ13C were also indicating a terrestrial diet (Montgomery et al. Reference Montgomery, Beaumont, Jay, Keefe, Gledhill, Cook, Dockrill and Melton2013). During periods of fasting, fevers, and malnutrition, catabolism can occur. Body tissues are recycled with a resulting rise in δ15N without a corresponding rise in δ13C (Mekota et al. Reference Mekota, Grupe, Ufer and Cuntz2006; Duška et al. Reference Duška, Tůma, Mokrejš, Kuběna and Anděla2007). The hypothesis that Ballevullin 1 was subject to such a period of stress is supported by the lower adult bone collagen δ15N value.
Overall, the isotopic results are all consistent with a childhood spent in the Hebrides for Balevullin 1 but indicate she experienced some dietary or physiological change between the approximate ages of 4 and 14 years possibly following, or due to, the removal of marine protein or breast milk from the diet.
Discussion
Prior to the present work, our understanding of Neolithic funerary activity in Argyll in the Inner Hebrides has been largely confined to the evidence from monumental chambered cairns. Although numerous examples are known from the region, however, none has so far been identified on Tiree and the distribution on adjacent islands and the nearby mainland is extremely sparse (Ritchie Reference Ritchie1997, fig. 5.1). Neolithic activity is known from recent re-examination of chipped stone assemblages, often from antiquarian collections including those of Ludovic Mann (Mithen et al. Reference Mithen, Pirie, Smith and Wicks2007), but no sites of the period have been systematically excavated.
Aside from Balevullin, only one other Neolithic flat inhumation cemetery is presently known in Britain, at Barrow Hills, Radley in Oxfordshire (Barclay & Halpin Reference Barclay and Halpin1999). Here the contracted burials of two adults (male and female) and a child were arranged on various orientations, with grave goods limited to a few flint flakes. An AMS date of 3380–3090 cal bc (Barclay & Halpin Reference Barclay and Halpin1999, 33), from the adult male, is consistent with that from Balevullin. Radiocarbon determinations from the other two skeletons produced similar results, but with wider standard deviations. Slightly more common are individual inhumations, including an unaccompanied child inhumation from Northton in the Isle of Harris, where the overlying midden has been dated by a single AMS determination to 3350–2890 cal bc (Simpson et al. Reference Simpson, Murphy and Gregory2006, 44). Elsewhere in Britain, individual inhumations are occasionally found in association with Neolithic funerary monuments, (eg, Saville Reference Saville1990; Morgan Reference Morgan1959) and in the ditches surrounding causewayed camps (eg, Smith Reference Smith1965; Mercer & Healy Reference Mercer and Healy2008). Gibson (Reference Gibson2007) has also discussed the occurrence of Neolithic inhumation burials under round barrows. Isolated inhumations, however, represent a seemingly sporadic minority rite.
There is nothing in the scattered Neolithic inhumations to suggest that these individuals were of high status. Children seem unusually well-represented, as at Balevullin (individual 4), Radley and Northton, while the unusual physical condition of the Balevullin 1 adult skeleton might even hint at some low status or marginal position for some of these individuals. The graves are non-monumental and generally devoid of grave goods. Energy expenditure is relatively minimal, even at Balevullin 1 where stones marked the grave. Inhumation was clearly not a normative rite and we should not assume that it was intended to honour the deceased or ensure their passage to the otherworld. Taylor (Reference Taylor2002) has suggested, for example that inhumation may in certain circumstances involve deliberate removal of certain individuals from the usual funerary rites and be intended to prevent the translation of their souls; perhaps through fear of troublesome ghosts, or as a punitive act.
The question remains as to how anyone could have contracted rickets on Neolithic Tiree. Vitamin D deficiency should not be a problem for anyone exposed to a rural outdoor lifestyle and able to metabolise vitamin D – certain genetic conditions can prevent the efficient production of vitamin D, but these are extremely rare (Malloy & Feldman Reference Malloy and Feldman2010). It is possible that Balevullin 1 may have experienced a period of famine as a result of conflict or crop failure which may have produced the observed changes in her δ15N dentine collagen profile, although no evidence is available from contemporaneous individuals to test this hypothesis. The strong implication, however, is that this individual, almost certainly a woman and probably local, was either kept indoors for much of their childhood, or else habitually wore clothing or costume that prevented adequate absorption of sunlight: the latter especially is a common cause of rickets in certain modern cultures (Hatun et al. Reference Hatun, Islam, Cizmecioglu, Kara, Babaoglu, Berk and Sevim Gökalp2005). There may of course have been a vicious cycle involved here, since initial illness might have led to confinement which would have progressively exacerbated the condition. The lack of any significant marine component in the diet may also have been a contributory factor, since consumption of oily fish could have provided an alternative source of vitamin D even for an individual kept indoors or covered up. Neolithic communities in the region appear to have engaged in dairy farming from the beginning of the period (Cramp et al. Reference Cramp, Jones, Sheridan, Smyth, Whelton, Mulville, Sharples and Evershed2014), and a lack of calcium, which can contribute to the development of skeletal rickets deformities (Brickley & Ives Reference Brickley and Ives2008, 93), is therefore also unlikely unless the individual was denied access to cow’s milk. The putative aetiology for the rickets must include the presence of a period of nutritional stress as suggested by the incremental dentine δ15N profile. Although the original osteological report identifies the presence of linear enamel hypoplasia, recent images of the teeth do not show this to be present on teeth which were forming at the time of the putative nutritional stress.
We can speculate on various scenarios that might have resulted in confinement or concealment. She might, for instance, have had some special religious role that confined her to a dark interior space, or entailed a costume that covered her body and shielded her face; alternatively she might simply have been a domestic slave seldom able to venture outdoors. There are of course innumerable possibilities, but these at least serve to demonstrate that neither high nor low status is necessarily entailed by her condition. Given the dearth of comparable skeletal material, however, it is too early to tell whether her situation was unique or whether rickets was a recurrent product of some cultural practice peculiar to the region.
Acknowledgements
We would like to thank Sally-Anne Coupar of the Hunterian Museum, Glasgow, for facilitating the analysis and for providing images of the dentition, and Dr Anne Pirie for providing her unpublished reports on the Bishop Collection. Historic Scotland funded the AMS dates and we are especially grateful to Rod McCullagh and Professor Gordon Cook at SUERC for support in obtaining additional determinations to verify the initial results. Dr Jo Buckberry and Professor Megan Brickley provided helpful comments on the osteological work. We would also like to thank Andy Gledhill at the University of Bradford, and Dr Jane Evans and Carolyn Chenery at NIGL, Keyworth for producing the isotope data. Figures 2 and 3 were prepared by Rachael Kershaw.
Online Supplementary material
Details of the methods employed in staple isotope analysis. To view supplementary material for this article, please visit http://dx.doi.org/10.1017/ppr.2015.7












