


This paper contains supplementary material that can be found online at http://journals.cambridge.org
Introduction
Tropical dry forest, also known as seasonally dry tropical forest, can be loosely defined as forest in frost-free regions with 500–2,000 mm of precipitation annually and a pronounced dry season of 4–7 months (Walter, Reference Walter1971; Murphy & Lugo, Reference Murphy and Lugo1986; Miles et al., Reference Miles, Newton, DeFries, Ravilious, May and Blyth2006). Isolated oceanic islands in the Pacific contain tropical dry forest and a number of endemic plant species with small geographical ranges (Mueller-Dombois & Fosberg, Reference Mueller-Dombois and Fosberg1998). There have been high levels of deforestation and degradation of tropical dry forests on these islands, which have a long history of anthropogenic disturbance (Rolett & Diamond, Reference Rolett and Diamond2004). New Caledonia, Fiji, the Marquesas and Hawaii have been deforested for agriculture, and forests have been degraded by fire, grazing and invasion of non-native species of plants and animals (Cuddihy & Stone, Reference Cuddihy and Stone1990; Cabin et al., Reference Cabin, Weller, Lorence, Cordell and Hadway2002; Meyer, Reference Meyer2004). According to the definition of Fisher & Lindenmayer (Reference Fisher and Lindenmayer2007) the majority of remaining native dry forest fragments in the Pacific can be considered relictual because they have been reduced to <10% of their original cover, and almost all remaining fragments are <100 ha. Thus, they provide a good opportunity to examine methods used to identify patterns of species richness in fragmented systems and compare the conservation status of these forests with other regions (Gordon et al., Reference Gordon, Hawthorne, Reyes-García, Sandoval and Barrance2003).
There is a growing interest in identifying tropical forests and forest fragments in regions that have a high conservation priority at a global spatial scale (Miles et al., Reference Miles, Newton, DeFries, Ravilious, May and Blyth2006; Schmitt et al., Reference Schmitt, Burgess, Coad, Belokurov, Besançon and Boisrobert2009). However, there are few comparative data on the relationship between various metrics and stand species richness across different geographical regions (Gordon et al., Reference Gordon, Hawthorne, Reyes-García, Sandoval and Barrance2003) and there is an ongoing debate about the importance of species richness as a metric for conservation planning. Levels of species richness in stands of tropical forests have been identified as an important metric for studying the conservation value of forests and nature reserves (Arroyo-Rodriguez et al., Reference Arroyo-Rodríguez, Pineda, Escobar and Benítez-Malvido2008). However, some researchers maintain that the number of range-restricted or endemic species and presence or densities of threatened species are equally important metrics for prioritizing conservation areas (Kerr, Reference Kerr1997; Chapman et al., Reference Chapman, Underwood and Clarke2009).
Biodiversity hotspots are regions that contain exceptional concentrations of endemic species whose habitat is threatened by human activity (Myers et al., Reference Myers, Mittermeier, Mittermeier, da Fonseca and Kent2000; Myers, Reference Myers2003; Conservation International, 2009). The resources available for conservation in biodiversity hotspots are inadequate to protect all species and therefore it would be more effective to concentrate conservation efforts in areas with the highest levels of biodiversity and the highest numbers of threatened species. Gentry transects provide a standard and repeatable method that can be used to compare levels of species richness and risk of extinction (Gentry, Reference Gentry, Bullock, Mooney and Medina1995; Phillips & Miller, Reference Phillips and Miller2002), and geographical information system (GIS) polygons of tropical dry forest regions and satellite imagery (MODIS 500 m resolution) have been used to identify the extent of dry forests at a global spatial scale (Miles et al., Reference Miles, Newton, DeFries, Ravilious, May and Blyth2006; Schmitt et al., Reference Schmitt, Burgess, Coad, Belokurov, Besançon and Boisrobert2009; Portillo-Quintero & Sánchez-Azofeifa, Reference Portillo-Quintero and Sánchez-Azofeifa2010), although there are few comparative plot or forest extent data for biodiversity hotspots in the Pacific (Miles et al., Reference Miles, Newton, DeFries, Ravilious, May and Blyth2006; Pau et al., Reference Pau, Gillespie and Price2009; Pennington et al., Reference Pennington, Lavin and Oliveira-Filho2009; Schmitt et al., Reference Schmitt, Burgess, Coad, Belokurov, Besançon and Boisrobert2009). There have been significant advances in the spatial resolution of GIS and remote-sensing data, which can be used to assess the extent and conservation status of dry forests in biodiversity hotspots. WWF's ecoregions approach has provided comparative data on large units of land and water containing geographically distinct assemblages of species, natural communities and environmental conditions, at a global spatial scale (Olson et al., Reference Olson, Dinerstein, Wikramanayake, Burgess, Powell and Underwood2001), and there have been advances in the spatial resolution of land-cover classifications such as the European Space Agency's GlobCover dataset, which provides 22 land-cover classes (six terrestrial forest types) at a 300 × 300 m pixel resolution (ESA, 2010).
Our research had three primary objectives. Firstly, we examined the relationship between total species richness, native species richness, number of endemic species and number of threatened woody plant species (trees, shrubs, lianas) at the stand level. In particular, we tested the hypothesis that total species richness at the stand level is positively associated with native species richness, endemic species richness, and presence and density of threatened species across all four regions (New Caledonia, Fiji, Hawaii and the Marquesas). Secondly, we assessed the conservation priorities of tropical dry forest sites in the Pacific based on the presence and density of threatened species and levels of endemism. Thirdly, we calculated the extent of tropical dry forests and protected areas in the New Caledonia and Polynesia–Micronesia biodiversity hotspots to assess the conservation priorities in the region.
Study area
This research was undertaken on four oceanic island archipelagoes (New Caledonia, Fiji, the Marquesas and Hawaii) within the New Caledonia and Polynesia–Micronesia biodiversity hotspots (Myers et al., Reference Myers, Mittermeier, Mittermeier, da Fonseca and Kent2000; Fig. 1). Study sites were selected based on Landsat ETM+ data from 2005, WorldClim climate data extrapolated from climate stations (WorldClim, 2009), and Shuttle Radar Topography Mission data, to identify the best-preserved forest fragments in regions with 500–2,000 mm of annual precipitation and a pronounced dry season (Hijmans et al., Reference Hijmans, Cameron, Parra, Jones and Jarvis2005). We conducted field visits to potential sites to ensure that they hosted native tree species. We surveyed a total of 37 dry forest sites (Fig. 2). Seven sites were surveyed on the western side of Grand Terre, New Caledonia. Nine sites were surveyed in western Fiji: Yasawa group (4), Mamanuca group (1), Viti Levu (2) and islands near Viti Levu and Vanua Levu (2). Six sites were surveyed on the two largest islands in the Marquesas: Nuku Hiva (3) and Hiva Oa (3). Fifteen sites were surveyed on six Hawaiian islands: Kauai (4), Oahu (2), Molokai (1), Lanai (3), Maui (2), and the island of Hawaii (3). Detailed overviews of each region and our study areas are available at Gillespie (Reference Gillespie2013).

Fig. 1 Pacific archipelagos with tropical dry forests.

Fig. 2 Locations of our study sites in tropical dry forests on the Pacific islands of Hawaii, New Caledonia, the Marquesas and Fiji.
Methods
Data on species richness, floristic composition and forest structure at each study site were collected following Gentry (Reference Gentry1988). Belt transects of 1,000 m2 (0.1 ha) were established at each site. Each survey consisted of 10 transects (2 × 50 m), spaced 10 m apart, in which all plants of diameter at breast height ⩾ 2.5 cm rooted in the sample area were recorded (Gentry, Reference Gentry1988). In New Caledonia, Fiji, and Hawaii, plants were identified to species or morphospecies at the local herbaria: Institut de recherche pour le développement (IRD) in Nouméa, New Caledonia; South Pacific Regional Herbarium (SUVA) at the University of the South Pacific in Suva, Fiji; and Bishop Museum in Honolulu, Hawaii (BISH). All species from the Marquesas were identified according to Wagner & Lorence (Reference Wagner and Lorence2003). At each site species were identified as native, endemic to the regional archipelago or non-native, and threatened species were categorized as Critically Endangered, Endangered or Vulnerable according to the IUCN Red List (IUCN, 2012).
We collected four GIS layers to identify the extent of ecoregion, dry forest and protected forest in Pacific biodiversity hotspots. Ecoregion polygons were acquired from WWF (2009) for dry forest in the New Caledonia and Polynesia–Micronesia biodiversity hotspots (Conservation International, 2009). We acquired the European Space Agency's GlobCover v. 2.1 dataset (ESA, 2010), which contains three terrestrial forest land-cover classifications that correspond to tropical dry forest: (1) > 5 m tall closed to open (>15%) broadleaved evergreen or semi-deciduous forest; (2) closed (>40%) broadleaved deciduous forest; (3) open (15–40%) broadleaved deciduous forest. Protected area polygons were acquired from the World Database on Protected Areas (IUCN/UNEP, 2009). Only terrestrial protected areas in categories I to VI were included (IUCN, 2009).
Species richness data (total richness, native richness and endemic richness) had a normal distribution within each region, based on a one-sample Kolmogorov–Smirnov test, and therefore a Pearson product correlation was used to identify whether total species richness was associated with native richness or endemic richness. The number and density of Red-Listed species did not have a normal distribution, and thus we used a Spearman's rank correlation between total species richness and number and density of threatened species in 0.1 ha plots. We ranked conservation priorities of sites, based on the number and density of Red-Listed species and the number of endemic species in the 0.1 ha plots. Ecoregions within the Polynesia–Micronesia and New Caledonia biodiversity hotspots that contain dry forest were overlain on GlobCover data to calculate forest extent in each ecoregion. Protected area polygons were then overlain to identify the extent of forest in protected areas.
Results
A total of 265 species or morphospecies were identified from 8,769 stems at the 37 sites. Forests in New Caledonia and Fiji contained the highest species richness (Table 1), and forests in New Caledonia and Hawaii had the highest number and proportion of endemic and threatened species and the highest number of threatened individuals (Table 1; Supplementary Table 1). The dry forests of Fiji and the Marquesas had a high proportion of native species and a lower number and density of threatened species than New Caledonia and Hawaii.
Table 1 Number (and percentage) of native, endemic and non-native species, number of Red-Listed species (Critically Endangered, Endangered or Vulnerable), and number of individuals of Red-Listed species of woody plants ⩾ 2.5 cm diameter at breast height rooted within Gentry transects at our study sites on the Pacific islands of New Caledonia, Fiji, Marquesas and Hawaii (Fig. 2).

There were significant positive correlations between total woody plant species richness and native species richness in all four Pacific dry forests (Table 2). Total species richness was significantly correlated with endemic species richness in New Caledonia, Fiji and Hawaii. There were no correlations between total species richness and number of threatened species or density of threatened species in any of the forests.
Table 2 Correlations of total species richness with native and endemic species richness (Pearson product correlation) and number and density of Red-Listed species (Spearman's rank correlation) in 0.1 ha plots in tropical dry forest in New Caledonia, Fiji, Marquesas and Hawaii (Fig. 2). Blank cells indicate that no Red-Listed species were encountered in plots.

*P < 0.05; **P < 0.01; ***P < 0.001
We ranked the conservation priorities of the tropical dry forest sites (Supplementary Table 1). New Caledonia had the most sites with a high number and density of threatened species and number of endemic species, followed by Hawaii, Fiji and the Marquesas (Supplementary Table 2). In New Caledonia, Metzdorf, Nekoro and Pindai have a high conservation priority, and in the Hawaiian Islands Kokee (on the island of Kauai) and Kaupulehu (on the island of Hawaii) have a high conservation priority. Vatia, on the island of Viti Levu, has the highest conservation priority in Fiji, although Naviti and Yanuca both had a high number of endemic species. In the Marquesas we did not encounter any threatened species, and Hatiheu and Hakaui, on Nuku Hiva, had the highest levels of endemism.
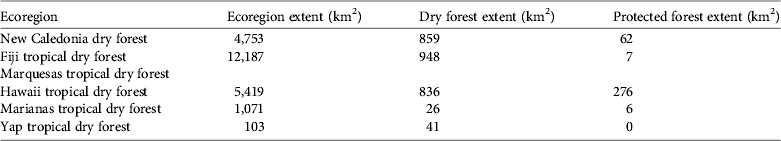
There are six tropical dry forest ecoregions (WWF, 2009) within the New Caledonia and Polynesia–Micronesia biodiversity hotspots (Table 3). Marquesas tropical dry forest is not recognized as an ecoregion and there are no land-cover data for this forest in the GlobCover dataset. Fijian tropical dry forest had the largest ecoregion extent, followed by Hawaii and New Caledonia tropical dry forest (Table 3). Forest extents within ecoregions were relatively similar for New Caledonia dry forest (18%), Hawaiian tropical dry forest (15%) and Fijian tropical dry forest (8%). Fiji had the smallest extent of forest in protected areas (< 1%), followed by New Caledonia (7%).
Table 3 Extent of dry forest (from GlobCover; ESA, 2010) and protected forest in ecoregions with tropical dry forest in the New Caledonia and Polynesia–Micronesia biodiversity hotspots. Blank cells indicate that no data are available.
Discussion
Relationships between species richness, endemism and endangerment
There were correlations between total species richness, native species richness and endemic species richness in fragmented stands of dry forest in the Pacific. However, there were no correlations between total species richness and number or density of Red-Listed species. This suggests that caution should be taken when assessing the conservation priority of fragments on the basis of total species richness within stands (Kerr, Reference Kerr1997; Chapman et al., Reference Chapman, Underwood and Clarke2009). For example, if simple measures of species richness were used to prioritize regions in the Pacific, Hawaii would have a low conservation priority. However, 90% of the 109 species of trees and shrubs in the dry forest of Hawaii are endemic and 45% are included on the IUCN Red List (Pau et al., Reference Pau, Gillespie and Price2009). Thus, studies of species richness patterns should also provide data on endemism and risk of extinction to accurately assess conservation priorities.
Gentry's transect method appears to be appropriate for quantifying patterns of species richness and forest structure in regions where few comparative field data exist. Applying this method, 48% of known dry forest trees and shrubs of Hawaii were encountered at 15 sites that covered a total of 1.5 ha (Pau et al., Reference Pau, Gillespie and Price2009) and 29% of all known dry forest woody plants in New Caledonia were encountered at six sites that covered a total of 0.6 ha (Jaffré et al., Reference Jaffré, Morat and Veillon1993). However, only 16% of the 49 Red-Listed dry forest species were encountered in Hawaii and 11% of 117 Red-Listed dry forest species were encountered in New Caledonia (Jaffré et al., Reference Jaffré, Morat and Veillon1993; Pau et al., Reference Pau, Gillespie and Price2009). This suggests that rare species may be missed and that extensive presence/absence data for remaining forest fragments are still needed.
Conservation assessment of tropical dry forests in the Pacific
Based on the number of threatened species and levels of endemism the highest-priority sites for dry forest conservation within the Pacific are in New Caledonia and Hawaii. The most extensive surveys of species in forest fragments have been carried out in New Caledonia, with comparative presence data for woody species available for > 71 sites (Jaffré et al., Reference Jaffré, Rigault and Munzinger2008). Gentry transects have been used to quantify the regeneration status of stands and threatened species, based on tree size class (Hequet & Rigault, Reference Hequet and Rigault2007). New Caledonia also has high levels of microendemism; i.e. subspecies that have evolved in select dry forest locations (Jaffré et al., Reference Jaffré, Rigault and Munzinger2008). These morphological variations are not as pronounced in other dry forest regions in the Pacific.
Presence/absence data are available for islands and the dry forest region of Hawaii (Wagner et al., Reference Wagner, Herbst and Sohmer1999; Pau et al., Reference Pau, Gillespie and Price2009) but there are few publicly available data on the distribution of native or threatened species for the dry forest region. For other dry forests in the USA (e.g. in South Florida), presence/absence data are available for all vascular plants for each fragment and standardized plot data are available for a majority of the remaining fragments (Gillespie, Reference Gillespie, Pennington, Lewis and Ratter2006; IRC, 2012). These types of occurrence data per fragment or reserve are useful for conservation assessments and monitoring the status of threatened species, especially given that 45% of dry forest tree species are on the federal endangered species list (Pau et al., Reference Pau, Gillespie and Price2009).
Dry forest fragments in Fiji occur in a highly modified landscape where frequent fires occur; only a small fraction of these remnants (0.7% of the forest cover) are in protected areas. Although species occurrence data exist for 10 dry forest sites in Fiji (Keppel & Tuiwawa, Reference Keppel and Tuiwawa2007) there are no published data for the Yasawa Islands, the driest region in Fiji. There is potential for conservation of forest fragments on these islands but systematic floristic inventories are needed. The IUCN categorization of tropical dry forest species in Fiji is not as complete as in New Caledonia, Hawaii and the Marquesas.
The tropical dry forests of the Marquesas contain some of the lowest levels of native species richness at the stand level because of the relatively small size and young age of the archipelago (1,050 km2; 3.7 million years old) and its isolation from continental land masses (Wagner & Lorence, Reference Wagner and Lorence2003). Decker (Reference Decker1973) highlighted the importance of protecting lowland dry and coastal forests in the Marquesas, which are threatened by overgrazing by feral ungulates (goats, sheep and pigs). Presence/absence data have been recorded for all of the Marquesas islands (Wagner & Lorence, Reference Wagner and Lorence2003) but extensive and systematic plant inventories are needed at dry, low elevations on smaller islands not included in our survey. Surveys of the dry lowlands of the Marquesas are also needed to identify the distribution of threatened small trees such as Abutilon sachetianum (Malvaceae; Vulnerable), Chamaesyce sachetiana (Euphorbiaceae; Vulnerable), and Rauvolfia sachetiae (Apocynaceae; Critically Endangered; Lorence & Butaud, Reference Lorence and Butaud2011; IUCN, 2012).
Comparisons with other tropical dry forests in biodiversity hotspots
We compared the species richness and the number of Red-Listed species categorized as Critically Endangered, Endangered or Vulnerable with other tropical dry forests in biodiversity hotspots in which Gentry's transect method has been employed and where 95% of the plants encountered were identified to species (Lott et al., Reference Lott, Bullock and Solis-Magallanes1987; Gentry, Reference Gentry, Bullock, Mooney and Medina1995; Gillespie et al., Reference Gillespie, Grijalva and Farris2000; Phillips & Miller, Reference Phillips and Miller2002). We found that species richness at the stand level was lower in the Pacific than in mainland biodiversity hotspots (Supplementary Table 3). Stands of dry forest in New Caledonia, Costa Rica, Tanzania, Thailand and Ecuador contain the highest number of threatened species. However, many trees in Costa Rica and Thailand are on the IUCN Red List because they are threatened by logging but they may yet be very common and have large geographical ranges (Phillips & Miller, Reference Phillips and Miller2002; Gordon et al., Reference Gordon, Hawthorne, Reyes-García, Sandoval and Barrance2003). In comparison, Red-Listed species in New Caledonia are restricted to a small number of sites (Jaffré et al., Reference Jaffré, Rigault and Munzinger2008) and New Caledonian dry forests have a high proportion of Red-Listed species compared to other regions in which Gentry's transect method has been employed. The dry forests of New Caledonia therefore appear to be a high priority for conservation compared to other tropical dry forests in biodiversity hotspots (Jaffré et al., Reference Jaffré, Bouchet and Veillon1998).
Dry forests in the Pacific and Caribbean have a smaller geographical extent and are included in fewer protected areas compared to other dry forests in biodiversity hotspots (Supplementary Table 4). Dry forests in Fiji, Ecuador and New Caledonia have the lowest percentage of protected area of forest. The dry forest cover in the Pacific is estimated to be < 1,000 km2 but the actual extent of native forest may be significantly less. Although ecoregions are useful for identifying the location and distribution of ecosystem types at a global spatial scale and the World Database on Protected Areas is useful for identifying protected areas within ecoregions, there are currently no remote-sensing or GIS data on native forests vs non-native forests (i.e. plantations, forests dominated by non-native tree species) for forest fragments in the Pacific outside of Hawaii, which recently produced a gap analysis or land cover map at a 30 × 30 m pixel resolution with non-native forest types (Gillespie et al., Reference Gillespie, Chu and Pau2008). Such data would be useful in the Pacific and other biodiversity hotspots that have a large extent of forest dominated by non-native species.
Implications for conservation
There are associations between total plant species richness and native and endemic species richness in the dry forests of the four Pacific island groups that we studied but we found no significant association between species richness and the number or density of Red-Listed species. This suggests that although Gentry's transect method provides important comparative data on patterns of species richness, the method does not correlate with the number or density of Red-Listed species. Conservation priorities within regions should be based on occurrence and density data for threatened species in remaining fragments. Globally, the dry forests of Hawaii and New Caledonia merit a high conservation priority because of their high endemism and high numbers of species at risk of extinction. New Caledonia and Fiji have a small extent of dry forest and protected forest compared with other biodiversity hotspots that contain dry forest. Although our study identified high priority areas for conservation of dry forest it also highlighted the limited floristic data available for many Pacific dry forests and the limitations of applying global datasets for the region (i.e. there are no land-cover data for the Marquesas). More comparative plot data, presence/absence data in fragments, and GIS data are required to adequately assess and protect the dry forests of the Pacific.
Acknowledgements
We thank Christophe Lambert (Environmental Service of the Southern Province) for permission to work in New Caledonia, and Gilles Dagostini and Jerôme Munzinger for help with plant identification at the IRD Herbarium at Nouméa. We thank the National Trust of Fiji and especially local people and village chiefs for permission to do field research in Fiji, and Laura L. Williams from the Division of Fish and Wildlife, who was extremely helpful in the Mariana Islands. We thank the federal and state agencies and the Nature Conservancy of Hawaii for granting us permission to do research in the Hawaiian Islands. We also thank SALVIAS for providing data from Gentry transects, and Cella Carr and two reviewers for their helpful comments and suggestions. This research was funded by the National Geographic Society, National Science Foundation BCS0455052, UCLA Department of Geography and the NASA Earth System Science Fellowship.
Biographical sketches
Thomas W. Gillespie is a geographer interested in using field data, geographical information systems, and remote-sensing data to predict patterns of species richness and rarity in tropical plants. Kristin O’Neill is a biogeographer interested in island ecosystems. Gunnar Keppel is an ecologist interested in biogeography, community ecology, islands, evolution and population genetics. Stephanie Pau is a biogeographer interested in understanding the dynamic spatial and temporal responses of ecosystems to climate variability. Jean-Yves Meyer is a plant ecologist, field botanist and conservation biologist, with expertise in invasive alien species. Jonathan P. Price is a biogeographer interested in understanding spatial patterns in biodiversity, including how these patterns have originated and how human activity has modified them. Tanguy Jaffré is a botanist interested in the biodiversity and conservation of the flora of New Caledonia.






