


Introduction
Monitoring wildlife populations is important for conservation management but is often difficult to achieve effectively, particularly for rare and cryptic species (e.g. Gese, Reference Gese, Gittleman, Funk, Macdonald and Wayne2001). Monitoring carnivores is challenging because they are often threatened and exist at low densities and in fragmented populations (Gese, Reference Gese, Gittleman, Funk, Macdonald and Wayne2001). Carnivores suffer from many, mostly anthropogenic, threats, including direct persecution, habitat loss and competition with humans for prey. Lack of information on the current status of many carnivores may be hindering effective conservation action (Gittleman et al., Reference Gittleman, Funk, Macdonald and Wayne2001).
The Scottish wildcat is a subpopulation of the European wildcat Felis silvestris silvestris and is Britain's only surviving native felid (Macdonald et al., Reference Macdonald, Daniels, Driscoll, Kitchener and Yamaguchi2004). Although widely distributed (Africa, Asia, Europe) and categorized as Least Concern (Driscoll & Nowell, Reference Driscoll and Nowell2009), this species is subject to several threats globally, resulting in local extinctions and population fragmentation, especially in Europe (Nowell & Jackson, Reference Nowell and Jackson1996). Recent estimates, from the proportion of cats with wildcat pelage from a 1990s sample, indicate the Scottish population may be Critically Endangered, with < 400 genetically pure individuals remaining (Kitchener et al., Reference Kitchener, Yamaguchi, Ward and Macdonald2005; Driscoll & Nowell, Reference Driscoll and Nowell2009). Once widespread across Britain, habitat loss (Nowell & Jackson, Reference Nowell and Jackson1996), persecution (Langley & Yalden, Reference Langley and Yalden1977; Tapper, Reference Tapper1992; Kitchener, Reference Kitchener1995) and hybridization with feral domestic cats Felis catus (McOrist et al., Reference McOrist, Boid, Jones, Easterbee, Hubbard and Jarrett1991; Hubbard et al., Reference Hubbard, McOrist, Jones, Boid, Scott and Easterbee1992; Beaumont et al., Reference Beaumont, Barratt, Gottelli, Kitchener, Daniels, Pritchard and Bruford2001; Daniels et al., Reference Daniels, Beaumont, Johnson, Balharry, Macdonald and Barratt2001) have now restricted wildcats to northern Scotland (Balharry & Daniels, Reference Balharry and Daniels1998; Daniels et al., Reference Daniels, Balharry, Hirst, Aspinall and Kitchener1998; Davies & Gray, Reference Davies and Gray2010).
Hybridization is currently considered the greatest threat to this species (Nowell & Jackson, Reference Nowell and Jackson1996; Macdonald et al., Reference Macdonald, Daniels, Driscoll, Kitchener and Yamaguchi2004, Reference Macdonald, Yamaguchi, Kitchener, Daniels, Kilshaw, Driscoll, Macdonald and Loveridge2010). Documented since the 18th century (Berwick, Reference Berwick1920), hybridization has potentially occurred since domestic cats arrived in Britain 2,000–3,000 years ago (Clutton-Brock, Reference Clutton-Brock1987; Serpell, Reference Serpell, Turner and Bateson2000). Extensive introgressive hybridization has led to difficulties in distinguishing Scottish wildcats from some wildcat × feral cat hybrids, complicating enforcement of protective legislation, hindering monitoring and making management potentially ineffective (Macdonald et al., Reference Macdonald, Daniels, Driscoll, Kitchener and Yamaguchi2004, Reference Macdonald, Yamaguchi, Kitchener, Daniels, Kilshaw, Driscoll, Macdonald and Loveridge2010). Kitchener et al. (Reference Kitchener, Yamaguchi, Ward and Macdonald2005) identified seven principal and eight subsidiary pelage characters to distinguish Scottish wildcats from hybrids and feral domestic cats, providing an objective method for identifying wild-living cats in the field. Monitoring methods for wildcats to date include road traffic accident surveys, live trapping, interviews and questionnaires, or combinations of these (Easterbee et al., Reference Easterbee, Hepburn and Jefferies1991; Balharry & Daniels, Reference Balharry and Daniels1998; Daniels et al., Reference Daniels, Balharry, Hirst, Aspinall and Kitchener1998; Davies & Gray, Reference Davies and Gray2010). Although these methods generate useful data, each has limitations. For example, road traffic accident surveys may be biased towards hybrids and feral cats, which scavenge more frequently on roads as a result of competition with larger wildcats (A. Kitchener, unpubl. data). Live trapping is time consuming, and requires experience and licensing under British law. Information from questionnaires and interviews depends on respondent experience and this method is liable to observer error (Davies & Gray, Reference Davies and Gray2010). Also, except for the survey of Davies & Gray (Reference Davies and Gray2010), most data on Scottish wildcats were collected before the development of the current pelage identification method. Given the limitations and biases of existing methods, exploring other survey methodologies, such as camera trapping, is desirable.
Camera trapping can provide much useful data (e.g. Karanth & Nichols, Reference Karanth and Nichols1998, Reference Karanth and Nichols2002; Carbone et al., Reference Carbone, Coulson, Christie, Conforti, Seidensticker and Franklin2001), including population-density estimates for monitoring studies (Yoccoz et al., Reference Yoccoz, Nichols and Boulinier2001; Martin et al., Reference Martin, Kitchens and Hines2007). Presence and abundance of European wildcats have been determined from camera trapping, which has also allowed individual identification (Monterroso et al., Reference Monterroso, Sarmento, Ferreras, Alves and Herrmann2005; Anile et al., Reference Anile, Bizzarri and Ragni2009; Sarmento et al., Reference Sarmento, Cruz, Eira and Fonseca2009). This study aimed to (1) determine the feasibility of camera trapping for surveying and monitoring Scottish wildcats, (2) compare the success of baited, scented and unbaited camera traps, and (3) develop a camera trapping protocol for future surveys. Population-density estimates under different models were generated using spatially explicit capture–recapture analysis.
Study area
The study was carried out on Seafield and Strathspey Estates in north-east Scotland (Fig. 1), partly within the Cairngorms National Park, of which 57.6 km2 is designated a Site of Special Scientific Interest. Comprising a mixture of heather moorland, Scots pine Pinus sylvestris plantations, birch woodland (Betula sp.) and rough grazing, the site supports diverse wildlife, including Scottish wildcats. Traditionally used for red grouse Lagopus lagopus scotica shooting and deer stalking, predator control was important for estate management. Although the estate is now primarily used for tourism and deer stalking, predator control continues, mainly to protect capercaillie Tetrao urogallus leks (Seafield & Strathspey Estates, 2001). The estate was selected because putative wildcats had been seen by gamekeepers and wild-living cats were present regularly (Estate manager, pers. comm., 2009).
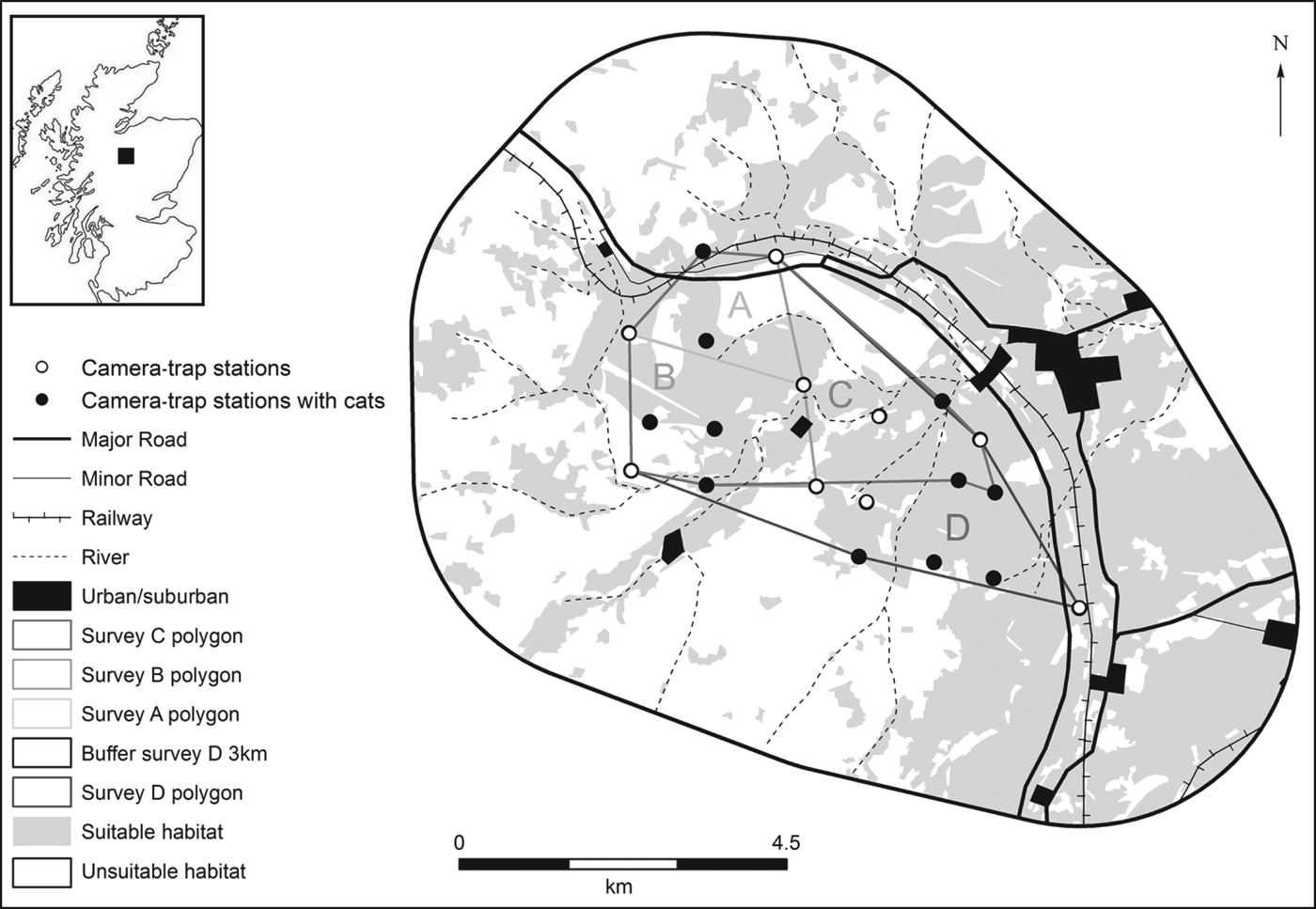
Fig. 1 The study site at the Seafield and Strathspey Estates, showing the locations of the camera-trap stations and their associated habitat: suitable (woodland, scrub and pasture/grassland) and unsuitable (arable, urban/suburban, heather moorland, bog and montane). The rectangle on the inset indicates the location of the main figure in north-east Scotland. A, B, C and D refer to the subsets of camera traps used in the spatially explicit capture–recapture analysis examining how a reduction in survey area affects density estimates.
Methods
Twenty camera trap stations were placed in a 4 × 5 grid. Based on the minimum home range of female Scottish wildcats (Corbett, Reference Corbett1979; Daniels et al., Reference Daniels, Beaumont, Johnson, Balharry, Macdonald and Barratt2001), stations were located 0.8–1.5 km apart so that individuals with the smallest recorded home range had a probability of > 0 of encountering a station (Karanth & Nichols, Reference Karanth and Nichols1998). Two Cuddleback Capture 3.0 (Cuddleback Digital, Green Bay, USA) camera traps were used per station. The first station was located where a cat with wildcat-type pelage had recently been trapped alive and released following standard estate predator control practices. The remaining 19 stations were arranged around this site. Stations were located where either wild-living cat signs (footprints, scats, dens, scrape marks) were present or where there were signs of pine martens Martes martes (which have similar habitat and prey requirements to the Scottish wildcat; Balharry, Reference Balharry1993; Birks et al., Reference Birks, Messenger and Halliwell2005) or rabbits Oryctolagus cuniculus (e.g. burrows, sightings, footprints, latrines). Camera traps were set across obvious animal trails 2.1–10.8 m apart, facing each other (but slightly staggered, to avoid flashes interfering with photographs from opposite cameras), to ensure both sides of wildcats were photographed for individual identification. Some camera traps were angled or moved slightly to face baits or lures in surveys using these attractors. Camera traps were attached to suitable trees or fence posts 20–150 cm above ground level, to achieve the best angle for photographing pelage characteristics.
There were three successive camera trap surveys. Survey 1 during February–March 2010 (900 trap nights) did not use bait or lures. Survey 2 during March–April 2010 (560 trap nights) used pheasant/partridge bait, which was attached to trees or posts at heights of 60–80 cm to encourage cats to stretch up and expose the dorsal, neck and shoulder regions for pelage assessments. Survey 3 during April–May 2010 (500 trap nights) used valerian tincture as a scent lure. Following studies on European wildcats (e.g. Weber, Reference Weber2008), lures comprised rough-surfaced wooden stakes (c. 60 cm length) in the ground between the camera traps. The upper 1/3 of the stake was coated with undiluted valerian tincture (AVogel/Holland & Barrett), extracted from dried roots of valerian Valeriana officinalis; it is believed to have a similar effect on cats as catnip Nepeta cataria.
Wild-living cats caught on camera were identified as wildcat, hybrid or domestic/feral cat based on seven key pelage characters, following Kitchener et al. (Reference Kitchener, Yamaguchi, Ward and Macdonald2005). Scoring was 1 (domestic), 2 (hybrid) and 3 (wildcat) for each pelage character. Any individual with a total score of ⩾ 14 and no scores of 1 for any character was considered a wildcat unless other data conflicted with this (e.g. white paws, white patches on flanks or back; Kitchener et al., Reference Kitchener, Yamaguchi, Ward and Macdonald2005). Hybrids could score 3, 2 or 1 for any of the characters and domestic cats had no scores of 3 for any characters.
Camera-trapping density estimates were compared to similar data collected during road traffic accident surveys and from sightings by estate staff. Roads and estate tracks surrounding and crossing the survey area were checked for cat carcasses every 10–14 days, while camera traps were checked. Estate staff were regularly questioned about cat sightings during and after surveys.
Data analysis
Data were managed using Camera Base (Tobler, Reference Tobler2010), displayed in ArcGIS v. 9.3 (ESRI, Redlands, USA) and analysed using SPSS v. 21.0 (SPSS, Chicago, USA). The Kolmogorov-Smirnov non-parametric test was used if residuals were not distributed normally. SPACECAP v. 1.0.6 (Singh et al., Reference Singh, Gopalaswamy, Royle, Kumar and Karanth2010), with R v. 2.15.2 (R Development Core Team, 2012), was used to generate estimates of population density.
Trap nights were calculated as the total number of 24-hour periods that all stations were active (Table 1). Capture events were the capture of an individual at a trap station within a 24-hour period. If the same individual was re-photographed within the 24-hour period this was considered one capture event. If cats were not individually identifiable, or were difficult to identify, and re-captured within a short time period (an arbitrary 30-minute interval), we assumed these were the same individuals and recorded them as single capture events. Capture rates for wildcats, hybrids and feral cats were calculated as number of capture events per 100 trap nights.
Table 1 No. of capture events, individuals caught and no. of trap stations at which the Scottish wildcat Felis silvestris silvestris was captured at the Seafield and Strathspey Estates in north-east Scotland (Fig. 1).

* One hybrid was caught in both Survey 1 and Survey 2.
SPACECAP is designed to estimate animal population densities using photographic data and closed capture–recapture models in a Bayesian Framework. SPACECAP takes into account capture locations of individuals, thus incorporating spatially explicit capture heterogeneity into analyses to achieve more precise and accurate estimates (Royle et al., Reference Royle, Nichols, Karanth and Gopalaswamy2009). This method has advantages over traditional mark–recapture methods (e.g. CAPTURE; Otis et al., Reference Otis, Burnham, White and Anderson1978) because it avoids having to convert abundance estimates into population-density estimates using effective sampling area approaches, which may inaccurately represent distances moved by individuals, often resulting in violation of the closure assumption (Royle et al., Reference Royle, Nichols, Karanth and Gopalaswamy2009). SPACECAP also recognizes that individual trap encounter histories are the outcome of two processes: distribution of individuals and an encounter process that describes whether or not individuals are encountered by traps as a function of their location (Singh et al., Reference Singh, Gopalaswamy, Royle, Kumar and Karanth2010). The programme also deals with problems posed by individual heterogeneity in capture probabilities and offers non-asymptotic inferences, which are more appropriate for small samples of capture data typical of photo-capture studies (Singh et al., Reference Singh, Gopalaswamy, Royle, Kumar and Karanth2010).
In SPACECAP the surveyed area contains camera traps and an extended area around it called the ‘space state’. The space state is defined in ArcGIS as a fine mesh of equally spaced points (here, 500 × 500 m), representing all possible home-range centres of all animals in the survey area (Singh et al., Reference Singh, Gopalaswamy, Royle, Kumar and Karanth2010). It should be large enough to ensure that it contains all individuals potentially caught by the camera traps. In this case a 3 km buffer was added to a rectangle encompassing the outermost camera traps. Where these points fell on suitable habitat they scored 1, and for unsuitable habitat (water, urban/suburban areas, roads) they scored 0.
SPACECAP was initially run using the spatial capture–recapture model with no trap response and a bivariate normal model, for 50,000 iterations with a burn-in period of 1,000. The thinning rate = 1 (default value) and the data augmentation value = 100 under the basic model (c. 8 times the total number of individuals captured). The model was also run with the trap response present, which implements a trap-specific behavioural response under which the encounter probability (p2) in that trap increases or decreases after initial capture at that trap (p1). Spatially explicit capture–recapture models were run to examine how population-density estimates varied under different scenarios. The accuracy of the models was assessed using Bayesian P-values (adequate models with P close to 0.5, and inaccurate models with P closer to 0 or 1).
Post-hoc power analysis was carried out in G*Power v. 3.1.7 (Faul et al., Reference Faul, Erdfelder, Lang and Buchner2007) to examine whether camera trapping could detect changes in population density over an arbitrary four survey periods. Statistical power of 0.8 was considered sufficient. Power at increases and decreases in population density of 75, 50, 25 and 10% and the effect of decreasing the posterior standard deviation of means to 75, 50, 25 and 10% of original values were modelled to determine their impacts on whether camera trapping could detect changes in population density.
Population-density estimates for different camera-trap array sizes were also modelled. The trapping grid was divided into four subsets of camera traps; A, B, C and D, running sequentially from the north to south of each survey site (Fig. 1). Each consecutive subset encompassed the cameras contained within the previous one plus an additional five camera-trap stations. The effective areas covered by each subset's space state (outer rectangle plus 3 km buffer) were A (68 km2), B (80.75 km2), C (104.5 km2) and D (138 km2). The percentage of home-range centres falling on unsuitable habitat was comparable: A (4.5%), B (3.3%), C (4.3%) and D (3.8%; t = 0.629, P = 0.59, df = 3). The percentage of suitable land cover (mixture of woodland, scrub and grassland) in each subset did not differ significantly: A (85.95%), B (80.58%), C (80.89%) and D (76.25%; t = 3.719, P = 0.07, df = 3). Coniferous woodland (recently felled coniferous woodland, young plantation, mature) comprised 45–60% of habitat in each subset. The effect of survey length on population-density estimates was also modelled, using sampling periods of 80, 70, 60, 50, 40, 30, 20 and 10 days (Table 2).
Table 2 The variation in the parameters generated by SPACECAP when the data were modelled over survey lengths of 10–80 days, with number of cats captured, the Bayesian P-value (which indicates the accuracy of each model: adequate models have P close to 0.5 and inaccurate models have P closer to 0 or 1), mean movement parameter (σ), mean encounter frequency (λ0), mean number of individuals (n), and mean density of wild-living cats per 100 km2.

To examine whether camera-trap spacing was sufficient, a measure of home-range radius was calculated. Under the bivariate normal model for animal movement, the movement parameter σ from SPACECAP can be converted into an estimate of home-range radius. This estimate provides a useful check on the validity of camera-trap spacing by converting it into a measure of the diameter of the home range (e.g. Karanth & Nichols, Reference Karanth and Nichols2002).
Results
Cameras were active for a total of 1,954 trap nights, and successfully detected wildcats in an area where they were thought to occur. No cats were captured using valerian lures, and therefore Survey 3 was excluded from the analysis. No photographs of cats were obtained during periods of heavy snowfall, despite other species being photographed. No cat carcasses were found along the roads surveyed during the study, and gamekeepers did not see any cats during the surveys.
Thirty photographs of wild-living cats were obtained and cats were detected at 11 of the 20 trap stations. A minimum of 13 different individual wild-living cats were captured at least once during Surveys 1 and 2 (Table 1). From pelage characters (Plate 1) four were classified as wildcats, nine as hybrids and none as feral cats. Although not all characters were visible in all photographs cats classified as hybrids (total pelage characteristic scores ⩽ 14) had scores of 1, 2 or 3 for one or more of the seven pelage characteristics and had a mean total score of 9 ± SD 4. The four wildcats (total scores > 14) did not score 1 for any visible characters and had a mean total score of 16.5 ± SD 3.

Plate 1 Wild-living cats (two wildcats and two hybrids) photo-trapped, showing some of the pelage characteristics and pelage variation.
The mean time to first photo-capture was 624 ± SD 457 trap nights for all wild-living cats and 629 ± SD 605 trap nights for wildcats only (Fig. 2). Capture rate per 100 trap nights increased with bait, but not significantly (Mann Whitney U test: Z = 0.000, P = 1.0; Z = 0.408, P = 0.67; Z = 0.152, P = 0.91 for all cats, wildcats and hybrids, respectively; Table 1). In most cases use of bait also increased the number of photographs of individuals, making identification easier: 1 photograph per capture event without bait compared to a mean of 1.75 ±SD 2 photographs per capture event with bait (n = 21; range 1–5). In only three capture events were photographs obtained from both of the camera pair.

Fig. 2 Cumulative number of individual wild-living cats Felis silvestris silvestris photo-trapped across the three surveys.
The initial model using Surveys 1 and 2 combined over the complete survey length of 80 days generated an unrealistically high movement parameter (σ). Therefore no further results are shown for this survey length. At a survey length of 70 days population density was estimated to be 68.17 ± SD 9.47 individual wild-living cats per 100 km2, with a 95% posterior interval of 49–79 individuals per 100 km2 and an encounter probability (λ0) of 0.026, σ = 0.34 km, giving a 95% home-range radius of 0.84 km and estimated home range of 2.44 km2. The posterior mean population density per 0.25 km2 (home range centre) is shown in Fig. 3. Population density was greatest adjacent to a large rabbit population and on the edges of grassland/woodland habitat.

Fig. 3 Posterior mean cat density per 0.25 km2 across the space state (see text for details). Home range centres are centred in each square. Circles indicate camera trap stations.
Power analysis (Fig. 4) shows this level of confidence of the mean density (SD = ± 9.47) is sufficient to detect ⩾ 25% increase or decrease in mean population density with statistical power > 0.8. Modelling a reduction in posterior standard deviation of mean population density to ⩽ 25% of the original value increases power sufficiently (⩾ 0.8) to detect a 10% change in population size.

Fig. 4 Power analysis graph showing how the power of camera trapping to detect an increase or decrease in mean density of individual cats per 100 km2 over four surveys increases as the SD decreases from 100 to 10% of the original value.
Running the trap response model increased the population density estimate to 98.7 ± SD 19.22 individuals per 100 km2, with posterior intervals of 62–124 individuals per 100 km2. The encounter probability also increased (λ0 = 0.01), with a positive trap response observed (p1 = 0.01, p2 = 0.33), σ = 0.45 km, giving a 95% home-range radius of 1.1 km and home-range size of 3.8 km2. Bayesian P-value = 0.3, indicating the trap response model is less accurate than the original model without a trap response.
Population density estimates were robust to changes in survey length, with no significant difference in population densities for surveys lengths of 20–70 days (t = −0.866, P = 0.426, df = 6) although 95% confidence intervals were significantly higher (t = 11.523, P < 0.01, df = 6) and lower (t = −4.657, P = 0.06, df = 6) for shorter survey lengths (Table 2). The encounter probability λ0 was 0.02–0.05 and the movement parameter σ 0.3–0.5 km at different survey lengths. A 10-day survey length resulted in an unrealistically large estimate for σ, as did reducing the survey area, even by 25%.
Discussion
Camera trapping detected wildcats and wild-living cats when none were recorded in road traffic accident surveys or observed by estate staff. Furthermore, with camera trapping cats could be individually identified and classified from pelage characters, facilitating estimation of population densities.
There are few estimates of the population density of Scottish wildcats: 0.1 individuals per 10 km2 from radio-tracking in west Scotland (Scott et al., Reference Scott, Easterbee and Jefferies1993) and 3 per 10 km2 using radioactive scat surveys in the east (Corbett, Reference Corbett1979). In both studies it is unknown whether the estimates included mixtures of wildcats and hybrids, although this is probable given that both occurred before Kitchener et al. (Reference Kitchener, Yamaguchi, Ward and Macdonald2005) proposed the use of pelage criteria. Population density in our study was estimated to be 6.8 individuals per 10 km2. This estimate is based on all wild-living cats from Surveys 1 and 2 combined and therefore may be an underestimate as the use of bait produced higher capture rates during survey 2. The use of bait across the whole survey period may therefore have resulted in an increased number of captures overall and a corresponding greater population density estimate.
Running the spatially explicit capture–recapture model with a trap response resulted in a positive trap-specific behavioural response, with a greater encounter frequency resulting in a larger population density estimate, indicating baited cameras were revisited by individuals. Also, although not significant, baiting increased capture rate and overall numbers of capture events. Occasionally, baiting resulted in multiple photographs of the same individual, leading to more accurate identification. In particular, placing bait so cats had to stretch up to reach them, with one camera facing the bait, enabled several pelage characteristics to be seen more clearly, including neck stripes, shoulder stripes and dorsal line. Without bait these characters were often difficult to see. Using two cameras per station thus aids identification of individuals, facilitating more accurate estimates of population density (Jackson et al., Reference Jackson, Roe, Wangchuck and Hunter2005).
Valerian lures did not appear to attract any cats, despite their successful use in some studies of European wildcats (Hintermann & Weber, Reference Hintermann and Weber2008; Weber, Reference Weber2008), although not in all (Anile et al., Reference Anile, Bizzarri and Ragni2009). Hintermann & Weber (Reference Hintermann and Weber2008) suggested using hair-lure surveys during November–April for optimum results. Outside this period, valerian did not seem to attract cats. Survey 3 occurred during April–May, which may thus not have been optimal for using valerian lures. Studies have shown that a felid's response to catnip is genetically determined (Bradshaw, Reference Bradshaw1992); if a response to valerian is also genetic, all individuals in the study area, or Scottish wildcats (and possibly hybrids) in general, may be unable to detect it. However, even if valerian failed to attract cats some would be expected at camera traps by chance (as cats were photo-trapped in Survey 1 without lures). Lack of detection of cats in Survey 3 could be because (1) Survey 3 occurred when wildcats were giving birth, and thus female cats recorded in the previous two surveys may have had more restricted activity and ranges, whilst kittens were young, (2) male cats may have changed their ranges (Daniels, Reference Daniels1997) following changes in resource availability, (3) valerian may have deterred cats in the area. Further research could identify an effective scent lure.
Population density estimates were generally robust to reduced survey lengths, except for short (10 days) and long (80 days) periods, which both produced unrealistically high movement parameter estimates. A 10-day survey may be unsuitable because of the resulting small sample size and an 80-day survey because populations may not be closed over a longer period. Generally, upper and lower 95% confidence intervals were significantly greater when < 13 individuals were captured. Although cats were photo-captured after only 40 trap nights (within 2 nights of all stations being active), the effort required to detect sufficient individuals for reliable population estimates using SPACECAP (10–13; A. Golpanswany, pers. comm., 2012) was considerably greater, with at least 990 trap nights required for the minimum of 10 individuals. Generally, spatially explicit capture–recapture methods need ⩾ 20 recaptures for precise estimates of population density (Efford et al., Reference Efford, Borchers, Byrom, Thomson, Cooch and Conroy2009), which was only achieved by combining Surveys 1 and 2. The overall capture rate of wildcats per 100 trap nights (0.5) is similar to that of European wildcats in Portugal (0.51; Sarmento et al., Reference Sarmento, Cruz, Eira and Fonseca2009).
Reducing survey area size by even 25% generated unrealistically large values for the movement parameter. This suggests reducing survey area and/or using < 20 camera stations for wildcats may not produce accurate population-density estimates using spatially explicit capture–recapture. For monitoring, detecting changes in population trends is achievable if population density estimates are sufficiently accurate. Although both posterior standard deviations and posterior 95% confidence intervals decreased with increased survey lengths, even the population estimate from combining Surveys 1 and 2 still had a large confidence interval (49–79) and posterior standard deviation (9.47), which will not detect with confidence changes of ⩽ 10% in population density unless the posterior standard deviation is reduced. Further surveys should therefore attempt to reduce the standard deviation and confidence interval to maximize monitoring success. To estimate population density most effectively, camera-trap surveys using a minimum of 20 stations should operate for 60–70 days.
The estimated 95% home range radius was 0.84 km2, giving a home range of c. 2.44 km2. Under the trap response, the estimate of home range was c. 3.8 km2. Both estimates were higher than expected based on previous studies (e.g. Corbett, Reference Corbett1979; Daniels et al., Reference Daniels, Beaumont, Johnson, Balharry, Macdonald and Barratt2001) indicating that, in this study area at least, wildcat home ranges could be greater than expected. To take these larger home ranges into account, camera traps should be placed slightly further apart (c. 1.5 km), or more camera traps used, to increase the survey area and increase the number of individuals likely to be detected during the survey period, thereby improving density estimates.
Future studies using this method for wildcats should aim to increase capture probabilities, to improve robustness of population density estimates. Capture probabilities can be improved by using bait, increasing the number of camera traps to cover larger survey areas, using two cameras at each station to increase probability of photographing wildcats, and placing cameras ⩽ 1.5 km apart to maximize the capture probability of individuals. Increasing survey length is not recommended, because population closure cannot be ensured (Silver, Reference Silver2004). Also, different camera trap models have different capabilities, resulting in different detection abilities. Therefore, using higher specification camera traps (e.g. faster trigger speeds, greater detection distances, improved battery life) may increase capture probabilities. It is noteworthy that we did not photo-trap cats when snow was > 100 mm deep, even though other species were captured. Studies have shown that locomotion through deep snow is often difficult for European wildcats (Mermod & Liberek, Reference Mermod and Liberek2002), influencing what habitat they use during periods of heavy snowfall, and Scottish wildcats are probably affected similarly. Camera trapping wildcats in heavy snow will not therefore yield reliable results.
To summarize, our study demonstrates that camera trapping is effective for detecting Scottish wildcats and could be used for other wildcat populations, particularly where hybridization is less extensive and differences in pelage patterns between wildcats and domestic cats are more pronounced (Ragni & Possenti, Reference Ragni and Possenti1996). Camera trapping may prove particularly useful for identifying areas where wildcats, hybrids and domestic/feral cats co-exist, facilitating effective targeted management of wildcat populations. However, long-term monitoring requires increased capture success to detect changes in population trends with confidence.
Acknowledgements
We thank the staff of Seafield and Strathspey Estates for their assistance and cooperation, and David Hetherington (Cairngorms Wildcat Project), Keith Duncan and Mairi Cole (Scottish Natural Heritage) for assistance in finding a suitable study site. We also thank Neil Anderson for providing information on wildcats in the area and Mairi Cole and Jenny Bryce (Scottish Natural Heritage) for their comments. Paul Johnson acknowledges the support of the Whitley Trust and the John Ellerman Foundation. We thank Scottish Natural Heritage for jointly funding this project, along with the support of a grant to DWM from The European Nature Trust and the Recanati-Kaplan Foundation.
Biographical sketches
Kerry Kilshaw has researched the Scottish wildcat and other small carnivores in a range of environments. Paul Johnson is involved in conservation projects that span taxa from invertebrates to large carnivores. Andrew Kitchener is principal curator of vertebrates at National Museums Scotland and he has broad interests that include hybridization between native and introduced mammal species, geographical variation and biogeography, the effects of captivity on mammal and bird skeletal morphology, including ageing and pathology, and faunal change and zooarchaeology in Scotland. David Macdonald, founder and Director of WildCRU, has a background in behavioural ecology with an emphasis on carnivores, with a recent focus on environmental policy, economics and research strategy.







