



Introduction
The four extant species of horseshoe crabs (Xiphosura) are often touted as an example of morphological conservancy. The fossil record of the horseshoe crabs stretches all the way back to the Ordovician and reveals a rather different picture (e.g., Rudkin et al., Reference Rudkin, Young and Nowlan2008; Van Roy et al., Reference Van Roy, Orr, Botting, Muir, Vinther, Lefebvre, El Hariri and Briggs2010; Lamsdell et al., Reference Lamsdell, Isotalo, Rudkin and Martin2022). This fossil record, which has seen a renewed interest in recent years (e.g., King et al., Reference King, Stimson and Lucas2019; Tashman et al., Reference Tashman, Feldmann and Schweitzer2019; Haug & Haug, Reference Haug and Haug2020; Lamsdell, Reference Lamsdell2020; Bicknell et al., Reference Bicknell, Błażejowski, Wings, Hitij and Botton2021, Reference Bicknell, Kimmig, Budd, Legg, Bader, Haug, Kaiser, Laibl, Tashman and Campione2022; Lamsdell et al., Reference Lamsdell, Teruzzi, Pasini and Garassino2021), displays a remarkable diversity, both in terms of morphological disparity and species richness (>80 extinct species) (Bicknell & Pates, Reference Bicknell and Pates2020; Lamsdell, Reference Lamsdell2020). Furthermore, whereas modern-day horseshoe crabs are restricted to marine environments, multiple extinct species inhabited brackish and freshwater environments (Lamsdell, Reference Lamsdell2016, Reference Lamsdell2020). A phylogenetic analysis has shown that the transition from salt to freshwater environments occurred multiple times (Lamsdell, Reference Lamsdell2016).
One extinct species considered to have inhabited freshwater environments is Limulitella bronnii (Schimper, Reference Schimper1853) from France (Lamsdell, Reference Lamsdell2016; Bicknell & Pates, Reference Bicknell and Pates2020). Here, we present the first well-preserved horseshoe crab specimen from the Middle Triassic (Anisian) strata of the Winterswijk quarry complex in the eastern Netherlands, also attributable to L. bronnii. We discuss the implications of this new find for our understanding of the paleoecology of the Winterswijk deposits.
Geological background
Whereas small exposures of Upper Triassic, Early Jurassic and Oligocene sediments have been reported from the Winterswijk quarry complex (51.967°, 6.781°) in the eastern Netherlands (Herngreen et al., Reference Herngreen, Van Konijnenburg - Van Cittert and Oosterink2005; Klompmaker & Van den Berkmortel, Reference Klompmaker and Van den Berkmortel2007; Klompmaker et al., Reference Klompmaker, Herngreen and Oosterink2010; Diependaal & Reumer, Reference Diependaal and Reumer2021), Middle Triassic (Anisian) carbonates dominate the exposures (Herngreen et al., Reference Herngreen, Van Konijnenburg - Van Cittert and Oosterink2005; Boekschoten & Van den Bosch, Reference Boekschoten and Van den Bosch2019; Van Hinsbergen et al., Reference Van Hinsbergen, Van Hinsbergen, Langereis, Dekkers, Zanderink and Deenen2019). These Lower Muschelkalk deposits belong to the Vossenveld Formation and form an approximately 40 m thick outcrop (Hagdorn & Simon, Reference Hagdorn and Simon2010; During et al., Reference During, Pietersen, Voeten, Abadi, Verdegaal-Warmerdam, Schulp, Smit, Reijmer and Van der Lubbe2019). These deposits consist of finely laminated micritic limestones interspersed with marly limestones, dolomite, clayey marls and dolomitic clay layers (Borkhataria et al., Reference Borkhataria, Aigner and Pipping2006; Klein et al., Reference Klein, Voeten, Lankamp, Bleeker, Sichelschmidt, Liebrand, Nieweg and Sander2015; Maxwell et al., Reference Maxwell, Diependaal, Winkelhorst, Goris and Klein2016; During et al., Reference During, Pietersen, Voeten, Abadi, Verdegaal-Warmerdam, Schulp, Smit, Reijmer and Van der Lubbe2019). The sediments were deposited at the edge of the epicontinental Germanic-basin at a paleolatitude of 14 ± 3°N, in paleogeographic conditions somewhat reminiscent of the Persian Gulf today (Van Hinsbergen et al., Reference Van Hinsbergen, Van Hinsbergen, Langereis, Dekkers, Zanderink and Deenen2019). They documented an environment that alternated between shallow marine and sabkha-like (hypersaline) intertidal mudflats (Oosterink & Winkelhorst, Reference Oosterink and Winkelhorst2013; Klein et al., Reference Klein, Voeten, Lankamp, Bleeker, Sichelschmidt, Liebrand, Nieweg and Sander2015; During et al., Reference During, Pietersen, Voeten, Abadi, Verdegaal-Warmerdam, Schulp, Smit, Reijmer and Van der Lubbe2019). The intertidal nature of the deposits is emphasised by the preservation of algal laminates, polygonal mud cracks, burrowing traces, reptile trackways and reptile swimming traces (Knaust, Reference Knaust2013; Oosterink & Winkelhorst, Reference Oosterink and Winkelhorst2013; Maxwell et al., Reference Maxwell, Diependaal, Winkelhorst, Goris and Klein2016; Schulp et al., Reference Schulp, Bleeker, Haarhuis, Van Spronsen, During, Goris, Kaskes, Matteman, Winkelhorst and Winkelhorst2017; During et al., Reference During, Pietersen, Voeten, Abadi, Verdegaal-Warmerdam, Schulp, Smit, Reijmer and Van der Lubbe2019; Marchetti et al., Reference Marchetti, Van der Donck, Van Hylckama Vlieg and During2019). It should be noted that whereas saline conditions likely persisted throughout the entire stratigraphic profile, a hypersaline depositional environment has been suggested for the top section (top 10 m) of the stratigraphic profile in particular (During et al., Reference During, Pietersen, Voeten, Abadi, Verdegaal-Warmerdam, Schulp, Smit, Reijmer and Van der Lubbe2019).
The Anisian deposits at Winterswijk have yielded many different types of fossils. Reptiles, such as nothosaurs, pachypleurosaurs, placodonts and tanystropheids (Klein et al., Reference Klein, Voeten, Lankamp, Bleeker, Sichelschmidt, Liebrand, Nieweg and Sander2015; Heijne et al., Reference Heijne, Klein and Sander2019; Spiekman et al., Reference Spiekman, Bleeker, Dorst, De Haan, Winkelhorst and Voeten2019), range in preservation from isolated bones to almost complete skeletons (Heijne et al., Reference Heijne, Klein and Sander2019). Furthermore, actinopterygian and chondrichthyan fish remains belonging to at least ten different genera have also been discovered (Haarhuis & Diependaal, Reference Haarhuis and Diependaal2019). An abundance of trace fossils has also been reported including invertebrate grazing traces, invertebrate burrowing traces, fish swimming traces, several different types of reptile trackways and reptile swimming traces (Knaust, Reference Knaust2013, Reference Knaust2015; Schulp et al., Reference Schulp, Bleeker, Haarhuis, Van Spronsen, During, Goris, Kaskes, Matteman, Winkelhorst and Winkelhorst2017; Marchetti et al., Reference Marchetti, Van der Donck, Van Hylckama Vlieg and During2019). In addition, many different invertebrate fossils have also been described, including mollusks (18 bivalve species, 2 gastropods and 1 ammonite), brachiopods (2 species), possible jellyfish and representatives of several clades of arthropods (e.g., Oosterink, Reference Oosterink1981; Akkerman & Mulder, Reference Akkerman and Mulder2012; Oosterink & Winkelhorst, Reference Oosterink and Winkelhorst2013; Klompmaker, Reference Klompmaker2019; Van Eldijk et al., Reference Van Eldijk, Heerschop, Haarhuis, Winkelhorst and Klompmaker2019). Amongst these arthropod finds are the remains of three species of lobsters, Clytiopsis argentoratensis Bill, Reference Bill1914, Oosterinkia neerlandica Klompmaker & Fraaije, Reference Klompmaker and Fraaije2011, and Audogaster cf. P. spinosa (Assmann, Reference Assmann1927), which constitute the oldest known lobsters from the Netherlands (Klompmaker & Fraaije, Reference Klompmaker and Fraaije2011). Furthermore, the isopod Gelrincola winterswijkensis Schädel et al., Reference Schädel, Van Eldijk, Winkelhorst, Reumer and Haug2020, and the cycloid Halicyne oosterinkorum Schweitzer et al., Reference Schweitzer, Feldmann and Schinker2019, are only known from Winterswijk and are both only known from single specimens (Schweitzer et al., Reference Schweitzer, Feldmann and Schinker2019; Schädel et al., Reference Schädel, Van Eldijk, Winkelhorst, Reumer and Haug2020). In addition, Winterswijk is the only Muschelkalk outcrop that has yielded a small assemblage of insect remains, including, amongst others, the elytron of a beetle (Coleoptera), a partial dragonfly wing (Odonata) and a complete forewing of a cockroach (Blattodea) (Van Eldijk et al., Reference Van Eldijk, Goris, Haarhuis, Lankamp, Winkelhorst, Reumer, Nel and Wappler2017). In addition, a single, poorly preserved horseshoe crab has previously been described from Winterswijk by Hauschke et al. (Reference Hauschke, Oosterink and Wilde2009). This specimen was tentatively assigned to the genus Limulitella (Hauschke et al., Reference Hauschke, Oosterink and Wilde2009), which was subsequently confirmed by Zuber et al. (Reference Zuber, Laaß, Hamann, Kretschmer, Hauschke, Van de Kamp, Baumbach and Koenig2017).
The occurrence of the bivalve Myophoria vulgaris (von Schlotheim, Reference von Schlotheim1820), the occurrence of the ammonite Beneckeia buchi (von Alberti, Reference von Alberti1834) and palynological evidence (Herngreen et al., Reference Herngreen, Van Konijnenburg - Van Cittert and Oosterink2005) firmly date the outcrops at Winterswijk to the Anisian stage of the Triassic (247.2–242 million years ago) (Akkerman & Mulder, Reference Akkerman and Mulder2012; Cohen et al., Reference Cohen, Finney, Gibbard and Fan2013). Originally, 39 stratigraphic horizons were identified in the outcrops at Winterswijk by Oosterink (Reference Oosterink1986). However, during expansion of the quarry complex, new layers were identified by the Working Group Muschelkalk Winterswijk, which were added to the profile and numbered according to the system established by Oosterink (Reference Oosterink1986) (see Van Eldijk et al., Reference Van Eldijk, Goris, Haarhuis, Lankamp, Winkelhorst, Reumer, Nel and Wappler2017; During et al., Reference During, Pietersen, Voeten, Abadi, Verdegaal-Warmerdam, Schulp, Smit, Reijmer and Van der Lubbe2019). Amongst these newly added horizons is an approximately 1.20 m thick layer, known as layer 43, in which the specimen we describe here, RGM.1333509, was discovered by one of the authors (HW). Layer 43 has previously yielded exceptionally well-preserved fossil fishes (Maxwell et al., Reference Maxwell, Diependaal, Winkelhorst, Goris and Klein2016), insect remains (Van Eldijk et al., Reference Van Eldijk, Goris, Haarhuis, Lankamp, Winkelhorst, Reumer, Nel and Wappler2017) and the only known specimen of the isopod G. winterswijkensis (Schädel et al., Reference Schädel, Van Eldijk, Winkelhorst, Reumer and Haug2020). Based on the occurrence of the bivalve Neoschizodus orbicularis (Bronn, Reference Bronn1837), the top section of the profile at Winterswijk, to which layer 43 belongs, has been tentatively dated to the youngest substage of the Anisian (Illyrian) (Hagdorn & Simon, Reference Hagdorn and Simon2010). Herngreen et al. (Reference Herngreen, Van Konijnenburg - Van Cittert and Oosterink2005) suggested that part of the top section of the profile (up until approximately layer 37) belonged to the older Bithynian substage of the Anisian based on palynological data. A more recent work (During et al., Reference During, Pietersen, Voeten, Abadi, Verdegaal-Warmerdam, Schulp, Smit, Reijmer and Van der Lubbe2019) suggested that the exposed Muschelkalk layers span the Bithynian, Pelsonian and the earliest Illyrian, with layer 43 being ascribed to the earliest Illyrian.
Methods and materials
The specimen described here was found in August 2020 in the top section of the aforementioned layer 43 in the Winterswijk quarry (Fig. 1). The fossil specimen consists of a part and its counterpart. The specimen is housed in the collection of the Naturalis Biodiversity Center, Leiden, The Netherlands (RGM.1333509). Photographs were made using a Cannon Eos 77D camera, a Cannon EF 18–55 mm IS STM-lens and a Sigma 105 mm F2.8 EX DG OS HSM Macro lens. Scale bars were inserted, images were cropped and compound images were stitched together using Adobe Photoshop 2021. We used limulid xiphosuran terminology after Bicknell et al. (Reference Bicknell, Błażejowski, Wings, Hitij and Botton2021, fig. 1).

Fig. 1. Layer 43 of the Muschelkalk exposed in the Winterswijk quarry complex (∼30 m away from the location where specimen RGM.1333509 was found), the eastern Netherlands. (A). Blue line indicates entire layer 43. Hammer is ∼30 cm long. (B). Red line indicates the upper part of layer 43 in which the horseshoe crab was found. Pictures by Gerard Goris taken in April–May 2022.
Systematic palaeontology
Subphylum Chelicerata Heymons, Reference Heymons1901.
Class Xiphosura Latreille, Reference Latreille1802.
Order Xiphosurida Latreille, Reference Latreille1802.
Suborder Limulina Richter & Richter, Reference Richter and Richter1929.
Superfamily Limuloidea Zittel, Reference Zittel1885.
Family Limulidae Leach, Reference Leach and Levrault1819.
Genus Limulitella Størmer, Reference Størmer1952.
Type species
Limulites bronnii (Schimper, Reference Schimper1853).
Other species
Limulitella (?) liasokeuperensis (Braun, Reference Braun1860); L. tejraensis Błażejowski et al., Reference Błażejowski, Niedźwiedzki, Boukhalfa and Soussi2017; L. (?) volgensis (Ponomarenko, Reference Ponomarenko1985).
Limulitella bronnii (Schimper, Reference Schimper1853)

Fig. 2. Photo plate of specimen RGM.1333509 (part), Limulitella bronnii (Schimper, Reference Schimper1853) from the Anisian of Winterswijk. (A) shows a top view of the specimen. (B) A detail photograph of the compound eye and the ophthalmic ridge. (C) Detail of the opisthosoma. (D) (composite image) A photo taken from an angled perspective. (E) (composite image) Display of top view of the specimen but under oblique lighting. Scale bars: 10.0 mm.

Fig. 3. Photo plate of specimen RGM.1333509 (counterpart), Limulitella bronnii from the Anisian of Winterswijk. (A) A top view of the complete specimen. (B) A more detailed photograph. Scale bar: 10.0 mm.

Fig. 4. Holotype of Limulitella bronnii (Schimper, Reference Schimper1853) from the Middle Triassic (lower Anisian) Grès à Voltzia Formation of the Upper Buntsandstein in NE France. (A). Drawing in Schimper (Reference Schimper1853, pl. 3) interpreted to be inverted. (B). Photo of UNISTRA.2015.0.50968, courtesy of Lea Grauvogel-Stamm and used with permission. Scale bar: 10.0 mm.
Limulites bronnii Schimper, pp. 6–7, pl. 3.
Limulites bronni Schimper; Bill, Reference Bill1914, pp. 327–330, pl. 15.8.
Limulus sandbergeri Kirchner, Reference Kirchner1923, pp. 635–636, fig. on p. 635.
? Limulus bronni (Schimper); Pfannenstiel, Reference Pfannenstiel1928, pp. 536–541, figs. 1 and 2.
Limulitella bronni (Schimper); Størmer, Reference Størmer1952, fig. 1K.
Limulitella bronni (Schimper); Gall, Reference Gall1971, p. 35, pl. 7.1.
Limulitella bronni (Schimper); Mader, Reference Mader1984, fig. 5.
Limulitella bronni (Schimper); Gall & Grauvogel-Stamm, Reference Gall, Grauvogel-Stamm, Hauschke and Wilde1999, fig. 8.
Limulitella bronni (Schimper); Weber & Lebrun, Reference Weber and Lebrun2014, p. 11, figs. 3 and 4.
Limulitella bronni (Schimper); Röhling & Heunisch, Reference Röhling and Heunisch2010, fig. 5.
Limulitella bronnii (Schimper); Bicknell & Pates, Reference Bicknell and Pates2020, fig. 28A.
Limulitella bronnii (Schimper); Bicknell et al., Reference Bicknell, Błażejowski, Wings, Hitij and Botton2021, fig. 4C.
Diagnosis
Prosoma including genal spine wider than long (l/w ratio ∼0.75). Prosomal length ∼30% longer than thoracetron, as measured along axis. Genal spines not clearly separated from occipital bands, making this part relatively wide. Posterior margins of genal spines straight or nearly so, at ∼140° from posterior margin of central part of prosoma. Ophthalmic ridges somewhat sinuous. Cardiac margins straight, somewhat converging to anterior prosoma. Cardiac ridge present. Thoracetron subtriangular, with 6–7 movable spines laterally and two fixed spines posteriorly adjacent to longitudinal axis. Apodemal grooves converging to posterior thoracetron. Long medial ridge present. Flange with distinct groove, somewhat diverging from lateral margin of thoracetron toward posterior thoracetron. Telson nearly as long as combined length of prosoma and thoracetron [based on specimens in Schimper (Reference Schimper1853), Gall (Reference Gall1971), Bicknell & Pates (Reference Bicknell and Pates2020), and herein].
Studied material
RGM.1333509.
Type material
Holotype: UNISTRA.2015.0.50968 (collections de paléontologie, Jardin des sciences de l’Université de Strasbourg), figured in Weber & Lebrun (Reference Weber and Lebrun2014, p. 11, fig. 4) and herein (Fig. 4b).
Description
Prosoma including genal spine wider than long (l/w ratio ∼0.75), with faint ridge along outer margin. Prosomal length ∼30% longer than thoracetron, as measured along axis. Anterior portion prosoma not preserved. Genal spines not clearly separated from occipital bands, making this part relatively wide. Posterior margin of genal spine straight or nearly so, at ∼140° from posterior margin of central part of prosoma. Ophthalmic ridges sinuous due to presence of eye, anterior portion not exposed. Eyes directed laterally, convex toward lateral side, ∼4 times as long as wide and height comparable to width. Cardiac margins straight, converging to anterior prosoma. Cardiac ridge most prominent at base of axis, fainter more anteriorly. Ocelli not exposed. Thoracetron subtriangular, with 6 partially preserved, non-decorated, flattened, posterolaterally directed spines on left lateral margin. Lateral margins of thoracetron with small triangular extensions in between aforementioned spines and with faint, straight ridge slightly more inward. Apodemal grooves straight overall, may contain small beads, converging to posterior thoracetron. Long medial ridge extending towards posteriormost preserved part, more prominent than cardiac ridge. Flange with distinct groove, initially diverging from lateral margin of thoracetron toward posterior thoracetron, then appears to split and reconnect more posteriorly, perhaps due to preservation. Telson and posteriormost part thoracetron not preserved. (Description based on new specimen, RGM.1333509.)
Remarks
We assign the new specimen to Limulitella bronnii because the specimen herein resembles the figured specimens of L. bronnii in detail (e.g., Gall, Reference Gall1971, pl. 7.1; Weber & Lebrun, Reference Weber and Lebrun2014, p. 11, figs. 3 and 4; Bicknell & Pates, Reference Bicknell and Pates2020, fig. 28A), including the holotype (Fig. 4) that is less well preserved than other specimens. The Winterswijk specimen also originates from the Anisian and is also from Western Europe. A diagnosis of L. bronnii is provided for the first time herein.
The new specimen cannot be ascribed to Limulitella (?) liasokeuperensis from the Early Jurassic of Germany because representatives of this species have narrower genal spines and proportionally smaller eyes (Braun, Reference Braun1860, fig. 1; Hauschke & Wilde, Reference Hauschke and Wilde1984, figs. 1 and 3; Bicknell & Pates, Reference Bicknell and Pates2020, fig. 30D). Limulitella tejraensis from the Middle Triassic (Anisian–early Ladinian) of Tunisia, morphologically close to L. bronnii, appears to have an ophthalmic ridge that is more S-shaped overall and the interophthalmic ridge of L. tejraensis is more sinuous than the specimen herein (Błażejowski et al., Reference Błażejowski, Niedźwiedzki, Boukhalfa and Soussi2017, fig. 3). Limulitella (?) volgensis from the Early Triassic (Olenekian) of Russia exhibits a lower (inner) margin of the genal spine at a substantially lower angle relative to the axial posterior margin of the prosoma than in the specimen herein (compare Bicknell et al., Reference Bicknell, Błażejowski, Wings, Hitij and Botton2021, fig. 4F).
While Lamsdell (Reference Lamsdell2020) put both Limulitella henkeli (von Fritsch, Reference von Fritsch1906) and L. vicensis (Bleicher, Reference Bleicher1897) in different genera, Bicknell et al. (Reference Bicknell, Błażejowski, Wings, Hitij and Botton2021) did not. Therefore, we also compare L. bronnii to these species. The specimen herein does not compare favourably with L. henkeli from the Middle Triassic (Anisian) of Germany because representatives of this German species have a clear angle on the posterior side of the genal spine (or the occipital band is absent) (Bicknell et al., Reference Bicknell, Błażejowski, Wings, Hitij and Botton2021, fig. 4E), which is not seen in the specimen herein. Limulitella vicensis from the Late Triassic (Carnian-Norian) of France (Lamsdell, Reference Lamsdell2020, fig. 6D; Bicknell et al., Reference Bicknell, Błażejowski, Wings, Hitij and Botton2021, fig. 4A) appears to have smaller eyes and the ophthalmic ridge is straighter.
Limulitella bronnii was first reported from a quarry with Buntsandstein deposits near Wasselonne in NE France (Schimper, Reference Schimper1853), ∼500 km south of Winterswijk, from the Middle Triassic (lower Anisian) Grès à Voltzia Formation of the Upper Buntsandstein (Gall & Grauvogel-Stamm, Reference Gall and Grauvogel-Stamm2005), and was considered to be part of the freshwater to brackish fauna of this formation. Schimper (Reference Schimper1853, p. 7) reported that the specimen was found in 1851. Weber & Lebrun (Reference Weber and Lebrun2014, p. 11, fig. 4) figured a specimen reminiscent of the holotype, shown also herein (Fig. 4b). The locality information and date when the specimen was found provided by Weber & Lebrun (Reference Weber and Lebrun2014) (Soultz-les-Bains [∼10 km south of Wasselonne], in the early part of the 19th century) do not fully match the information provided by Schimper (Reference Schimper1853), but the features of the fossil and the size match the drawing and description by Schimper (Reference Schimper1853, pl. 3), so we accept this specimen to represent the holotype. The drawing in Schimper (Reference Schimper1853) may have been inverted due to the use of an engraving technique (e.g., Gould, Reference Gould and Gould1995; Allmon, Reference Allmon2007; Hyžný, Reference Hyžný2014) because the rock surrounding the fossil differs markedly in outline when comparing the drawing to the photographed specimen (Fig. 4), but the horseshoe crab is identical in the placement of many features (hole in between the cardiac margin and the ophthalmic ridge on the right side, details near telson, etc.). In that case, one would represent the part and the other the counterpart. Alternatively, Schimper (Reference Schimper1853) did not depict the rock surrounding the specimen accurately.
Bill (Reference Bill1914) figured a different specimen from the same locality near Wasselonne. Gall (Reference Gall1971, p. 35) and Weber & Lebrun (Reference Weber and Lebrun2014) reported this species from the Grès à Voltzia Formation exposed at several localities in the same area in NE France the type specimen originated from. The species has also been reported from Germany. A small specimen ascribed to this species was reported from Villingen in Baden-Württemberg, SW Germany, from the fluvial Plattensandstein Formation (Pfannenstiel, Reference Pfannenstiel1928) of the Upper Buntsandstein, which is Middle Triassic (early Anisian (Aegean)) in age (Hagdorn & Nitsch, Reference Hagdorn and Nitsch2009; Hochuli et al., Reference Hochuli, Schneebeli-Hermann, Brack, Ramseyer and Rebetez2020). Earlier, Kirchner (Reference Kirchner1923) described Limulus sandbergeri from the Plattensandstein of Bad Brückenau, Unterfranken, in southern Germany, a species subsequently synonymised with L. bronnii (e.g., Lamsdell, Reference Lamsdell2020).
A specimen provisionally (cf.) ascribed to this species from lowermost Jurassic marine deposits of Germany by Wincierz (Reference Wincierz1960) should not be considered conspecific because only the ventral side is preserved, implying that too few diagnostic characters of this species are available to assess this hypothesis. Moreover, the temporal gap of ∼50 million years renders conspecificity highly unlikely.
The new specimen represents the third country from which this species is recorded. It is also the youngest representative of the species because the specimen originates from the late part of the Anisian (earliest Illyrian). The temporal range of this species within the Anisian is now a few million years.
Stratigraphic and geographic distribution
Winterswijk quarry complex (sector M18, at approximately 51.96750°N, 6.78172°E), The Netherlands. Vossenveld Formation, upper part Layer 43, uppermost substage of the Anisian (earliest Illyrian).
Discussion and conclusions
Horseshoe crabs from the Netherlands are rare. Other than the specimen herein and the one from the same locality reported previously (Hauschke et al., Reference Hauschke, Oosterink and Wilde2009; Zuber et al., Reference Zuber, Laaß, Hamann, Kretschmer, Hauschke, Van de Kamp, Baumbach and Koenig2017), we are only aware of various Late Carboniferous (Westphalian) horseshoe crabs from the mine area in the southern part of the Limburg province in the southeastern Netherlands (Van der Heide, Reference Van der Heide1951). Van der Heide (Reference Van der Heide1951) reported on Belinurus reginae Baily, Reference Baily1863, Belinurus sp., Prestwichianella rotundata (Prestwich, Reference Prestwich1840), Euproops cambrensis Dix & Pringle, Reference Dix and Pringle1929, E. danae (Meek & Worthen, Reference Meek and Worthen1865), E. communis Van der Heide, Reference Van der Heide1951, E. orbicularis Van der Heide, Reference Van der Heide1951, and Euproops sp.
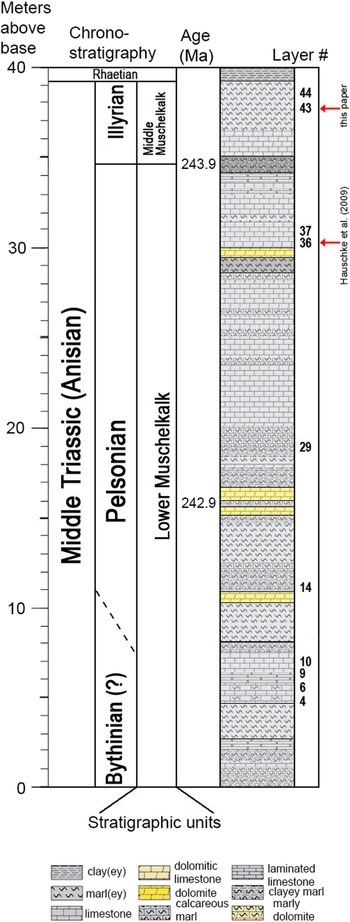
The first-reported horseshoe crab from Winterswijk identified as Limulitella sp. (Hauschke et al., Reference Hauschke, Oosterink and Wilde2009; Zuber et al., Reference Zuber, Laaß, Hamann, Kretschmer, Hauschke, Van de Kamp, Baumbach and Koenig2017) originated from layer 36, interpreted to be deposited during the Pelsonian substage (During et al., Reference During, Pietersen, Voeten, Abadi, Verdegaal-Warmerdam, Schulp, Smit, Reijmer and Van der Lubbe2019). Thus, the specimen herein from the upper part of layer 43 is younger, perhaps a few hundreds of thousands of years (During et al., Reference During, Pietersen, Voeten, Abadi, Verdegaal-Warmerdam, Schulp, Smit, Reijmer and Van der Lubbe2019, fig. 6) (Fig. 5). Diedrich (Reference Diedrich2011, fig. 5D) figured a horseshoe crab trackway (Kouphichnium Nopcsa, Reference Nopcsa1923) from Winterswijk, but the layer in which this specimen was found was not provided.

Fig. 5. Stratigraphy of the exposed Muschelkalk layers at the Winterswijk quarry complex in the eastern Netherlands with the horseshoe crab finds indicated. Modified from During et al. (Reference During, Pietersen, Voeten, Abadi, Verdegaal-Warmerdam, Schulp, Smit, Reijmer and Van der Lubbe2019, figs. 1 and 6), used with permission.
Little terrigenous influence was reported within layer 43 in During et al. (Reference During, Pietersen, Voeten, Abadi, Verdegaal-Warmerdam, Schulp, Smit, Reijmer and Van der Lubbe2019, fig. 1) based on a terrigenous weight percentage analysis (lithogenic wt% TGA). The observations of one of the authors (HW) indicate that the amount of marine versus terrestrial influence may vary both within the profile of 1.20-m-thick layer 43 as well as across the different places at which layer 43 has been sampled. Some parts appear fully marine given the presence of marine fish fossils (Wijngaarden et al., Reference Wijngaarden, Diependaal and Reumer2019), whereas other parts appear to have a terrestrial signal because of the presence of plant and insect fossils (e.g., Van Eldijk et al., Reference Van Eldijk, Goris, Haarhuis, Lankamp, Winkelhorst, Reumer, Nel and Wappler2017). Lobsters, fishes and insects found by one of us (HW) within layer 43 originate from horizons lower than the layer where the horseshoe crab originated from. Lobsters described previously, for which the exact stratigraphic layer is unknown, were interpreted to have inhabited a low energy, stressed environment with fluctuating salinity levels (Klompmaker & Fraaije, Reference Klompmaker and Fraaije2011). The specific horizon in which the horseshoe crab was found does not contain many other body fossils (pers. obs. HW). Although more detailed analyses within layer 43 would be welcome, many of the other faunal elements present in layer 43 suggest the horseshoe crab is unlikely to be marine. This suggestion is further corroborated by the fact that previous records of L. bronnii in Germany and France are considered non-marine (see above).
In addition to the horseshoe crab species herein, another part of the Winterswijk fauna resembles that of the interpreted non-marine faunal elements of the Grès à Voltzia Formation (Middle Triassic, lower Anisian) in the Vosges mountains of NE France (see Gall & Grauvogel-Stamm, Reference Gall and Grauvogel-Stamm2005, table 1). In terms of the brackish/freshwater elements, a lobster, a cyclidan crustacean and a bivalve overlap. The lobster Clytiopsis argentoratensis Bill, Reference Bill1914, is found in both deposits, and this species is the most common lobster at Winterswijk (Klompmaker & Fraaije, Reference Klompmaker and Fraaije2011). Congenerics of the cyclidan Halicyne are also known from both faunas (Gall & Grauvogel-Stamm, Reference Gall and Grauvogel-Stamm2005; Schweitzer et al., Reference Schweitzer, Feldmann and Schinker2019). For bivalves, Myophoria vulgaris occurs in both faunas and is even dominant at Winterswijk (Van Eldijk et al., Reference Van Eldijk, Heerschop, Haarhuis, Winkelhorst and Klompmaker2019). However, M. vulgaris occurs throughout the stratigraphic column at Winterswijk (Oosterink, Reference Oosterink1981, Reference Oosterink1986), and others have considered M. vulgaris to be also marine (Szulc et al., Reference Szulc, Matysik, Hagdorn and Haczewski2015). More terrestrial elements have been reported from both faunas such as plants and insects (e.g., Gall & Grauvogel-Stamm, Reference Gall and Grauvogel-Stamm2005; Van Eldijk et al., Reference Van Eldijk, Goris, Haarhuis, Lankamp, Winkelhorst, Reumer, Nel and Wappler2017).
In summary, the specimen of L. bronnii presented here expands the temporal range of this species. Furthermore, as this species is most probably non-marine, it could indicate some freshwater/terrestrial influx in the top section of the Winterswijk quarry complex. Finally, this specimen highlights the overlap between the fauna from Winterswijk and the fauna of the Grès à Voltzia Formation in NE France.
Acknowledgements
We thank Kévin Janneau (Université de Strasbourg), Lea Grauvogel-Stamm and Jean-Claude Gall (Université de Strasbourg) for providing information on the holotype of L. bronnii, Lea Grauvogel-Stamm for kindly sharing and permission to use a picture of the holotype, Gerard Goris for providing pictures and stratigraphic information about layer 43, René Fraaije (Oertijdmuseum) for a literature item, Melanie During (Uppsala University) for kindly providing high resolution figures that served as the basis for Fig. 5 and Gerard ten Dolle (Sibelco) for his continued support and access to the Winterswijk quarry complex. Our gratitude also goes to Carolin Haug (LMU Munich) and an anonymous reviewer for helpful comments that strengthened this paper.





 Open access
Open access