



Introduction
Scyllarids have been poorly studied, particularly in comparison to spiny and clawed lobsters (MacDiarmid & Sainte-Marie, Reference MacDiarmid, Sainte-Marie and Phillips2006), and the existing information is limited to a few commercially relevant species in the genera Ibacus (Leach, 1815), Scyllarides (Gill, 1898) and Thenus (Leach, 1816) (Sekiguchi et al., Reference Sekiguchi, Booth, Webber, Lavalli and Spanier2007). This is due to scyllarids traditionally having been regarded as secondary targets for fisheries. Nevertheless, since the 1990s, these lobsters have gained in commercial interest due to a decrease in the abundance of the main target species and, as a consequence, have suffered a sharp decrease in stock abundance at a local scale (Lavalli & Spanier, Reference Lavalli, Spanier, Lavalli and Spanier2007). The main reason for the alleged overexploitation of many scyllarids is the lack of information about their life history parameters, which are essential in order to precisely assess population size and resilience to fishing pressure or to environmental changes (O'Malley, Reference O'Malley2011).
In the particular case of Scyllarus arctus (Linnaeus, 1758), its increased commercial relevance can be confirmed by its high market values (€60 kg−1, approximate average price at first sale, and maximum of €188 kg−1, between 2001 and 2019 in Galicia, NW Iberian Peninsula), although the abundance of catches has not allowed for the development of a monospecific fishery. Captures of this scyllarid in Galicia are accessory and associated with coastal artisanal fisheries using octopus traps or trammel nets. In recent decades, despite some local regulation measures, the size of populations of this species on the Galician coast has been reduced (personal communication from fishermen), as has also been observed in the archipelago of the Azores (Santos et al., Reference Santos, Hawkins, Monteiro, Alves and Isidro1995) as well as in some coastal areas in the Mediterranean (Lloret & Riera, Reference Lloret and Riera2008).
A better knowledge of the reproductive biology of S. arctus is essential for the sustainable management of populations (Alborés et al., Reference Alborés, García-Soler and Fernández2019). Moreover, the scarce information about the biology of this species is focused on studies of Mediterranean populations (Pessani & Mura, Reference Pessani, Mura, Lavalli and Spanier2007) and is insufficient for developing appropriate measures for an effective management of populations across its broad geographic distribution. Within Crustacea, intraspecific differences in brood investment, number and size of produced eggs have been reported (Brante et al., Reference Brante, Fernández, Eckerle, Mark, Pörtner and Arntz2003; Verísimo et al., Reference Verísimo, Bernárdez, González-Gurriarán, Freire, Muiño and Fernández2011). Larger eggs contain more vitellum, which indicates substantial investment by the female in each embryo (Clarke, Reference Clarke1993), so that, after hatching, the larvae will have a greater chance of survival (Verísimo et al., Reference Verísimo, Bernárdez, González-Gurriarán, Freire, Muiño and Fernández2011; Alborés et al., Reference Alborés, García-Soler and Fernández2019).
Differences found in reproductive traits are attributed to environmental factors associated with habitat type along a latitudinal or bathymetric gradient (Brante et al., Reference Brante, Fernández, Eckerle, Mark, Pörtner and Arntz2003; Laptikhovsky, Reference Laptikhovsky2006; Fischer et al., Reference Fischer, Thatje and Brey2009; Alborés et al., Reference Alborés, García-Soler and Fernández2019), or to seasonal variability (Bas et al., Reference Bas, Spivak and Anger2007; Verísimo et al., Reference Verísimo, Bernárdez, González-Gurriarán, Freire, Muiño and Fernández2011; Urzúa et al., Reference Urzúa, Paschke, Gebauer and Anger2012). Furthermore, biological factors such as female size (Hines, Reference Hines1982; Verísimo et al., Reference Verísimo, Bernárdez, González-Gurriarán, Freire, Muiño and Fernández2011), energetic condition (Bryant & Hartnoll, Reference Bryant and Hartnoll1995; Verísimo et al., Reference Verísimo, Bernárdez, González-Gurriarán, Freire, Muiño and Fernández2011), age of reproductive individuals (Groeneveld, Reference Groeneveld2005) or interactions with other species (Griffen et al., Reference Griffen, Altman, Hurley and Mosblack2011), have been identified as other important sources of variation.
The aim of the present study is the characterization of the reproductive strategy of S. arctus in the NE Atlantic. With this purpose, brood size (fecundity and weight) and individual egg traits (size and weight) in early and late egg stages, were analysed in relation to female size and temporal variations (both seasonal and interannual) in two breeding cycles (2008 and 2009). In addition, we propose a tool for the regulation of this resource: a predictive model for the fecundity of S. arctus which takes lobster size and temporal variability during the reproductive period into account.
Materials and methods
Sampling
This study was carried out in Malpica (43°20′N 8°50′W), one of the main exploitation sites for the small European locust lobster Scyllarus arctus in Galicia (NW Iberian Peninsula), during the years of 2008 and 2009 (Figure 1). Individuals of S. arctus were obtained monthly from catches by artisanal fishing vessels using octopus traps. Captured lobsters were kept in insulated tanks containing cooled water, in order to reduce stress and avoid egg loss during transport. Once in the laboratory, wet weight (precision ± 1 mg), and cephalothorax length (CL) of each female were determined, as well as the distance between the rostral sinus and the posterior margin (to the nearest 0.01 mm). Egg masses were carefully removed from pleopods and submerged for 3 min in an ammonium formate solution (31.5 g l−1) in order to remove salts, being subsequently drained and weighed to obtain brood wet weight (BWW). Once brood colour was recorded, a sample of 30–40 eggs was taken from each brood and preserved in a 4% neutralized formaldehyde solution in order to determine embryonic developmental stage and measure egg size. Samples were then freeze-dried to constant weight for 48 h and brood dry weight (BDW) was determined with a precision of 0.1 mg. For this study, according both to macroscopic (colour) and microscopic criteria (see González-Gurriarán et al., Reference González-Gurriarán, Fernández, Freire and Muiño1996), two types of broods were used: broods in an early egg stage, constituted by yellow-coloured eggs with yolk occupying most of their volume (>80%) (from the blastula phase to an early differentiation of the naupliar mesoderm into an early gastrula), and broods in a late egg stage, characterized by brown-coloured eggs with well-developed larvae, where yolk occupied less than 10% or was absent.

Fig. 1. Location of the regular fishing ground for Scyllarus arctus in Malpica in the Costa da Morte (Galicia, NW Iberian Peninsula).
The gravimetric method was used to estimate fecundity (FE) from three replicates of egg subsamples. Previously, a study was performed to determine if there was an optimal egg sample size that allowed for a better estimation of FE (Verísimo et al., Reference Verísimo, Bernárdez, González-Gurriarán, Freire, Muiño and Fernández2011). Subsamples of freeze-dried eggs of known weight were separated in a Petri dish containing a 70% alcohol solution for subsequent counting. This was carried out through image analysis with Nikon's Nis Elements software, using a Nikon DXM 1200 digital camera coupled to a Nikon SMZ 1500 stereomicroscope. Likewise, image analysis was used to determine average egg volume (V) by measuring the major and minor axis of 30 previously preserved eggs per brood. Egg V was obtained using the oblate ellipsoid volume equation,
 $$\[V = \pi d_1^2{d_2}/6,\]$$
$$\[V = \pi d_1^2{d_2}/6,\]$$where d 1 is the average minor axis and d 2 is the major axis.
Data analysis
For the study of FE, broods from 212 ovigerous females were analysed as a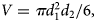 $V = \pi d_1^2 d_2/6\comma \;$ function of size, brood period and egg developmental stage. Three periods were established due to the extended spawning period during a reproductive cycle, the presence of berried females with eggs in various developmental stages during most of the year, including multiparous females (Alborés et al., Reference Alborés, García-Soler and Fernández2019), and the different environmental conditions (temperature, available food, etc.) in each period or season. An early period of the reproductive cycle was established between December and February, characterized as ‘winter brood’, as well as a second period between April and May, called ‘spring brood’, and a third period, the ‘summer brood’, comprised of broods of berried females obtained from July to September. Females captured in March and June were excluded from the analysis with the purpose of avoiding a possible overlap between periods. According to the stage of egg development in broods, FE estimates were divided as: realized FE, estimated as the number of eggs in broods with early stage eggs, and effective FE, estimated from broods with eggs close to hatching.
$V = \pi d_1^2 d_2/6\comma \;$ function of size, brood period and egg developmental stage. Three periods were established due to the extended spawning period during a reproductive cycle, the presence of berried females with eggs in various developmental stages during most of the year, including multiparous females (Alborés et al., Reference Alborés, García-Soler and Fernández2019), and the different environmental conditions (temperature, available food, etc.) in each period or season. An early period of the reproductive cycle was established between December and February, characterized as ‘winter brood’, as well as a second period between April and May, called ‘spring brood’, and a third period, the ‘summer brood’, comprised of broods of berried females obtained from July to September. Females captured in March and June were excluded from the analysis with the purpose of avoiding a possible overlap between periods. According to the stage of egg development in broods, FE estimates were divided as: realized FE, estimated as the number of eggs in broods with early stage eggs, and effective FE, estimated from broods with eggs close to hatching.
For the purpose of data analysis, it was necessary to transform FE, BDW, CL, egg dry weight (DWE) and egg V data using natural logarithms. Temporal differences within the two reproductive cycles studied, both for broods in early and late egg stages, were analysed through analysis of covariance (ANCOVA) using CL as covariable. Two-way ANCOVAs were performed to assess differences in reproductive traits among seasons and years (2008 and 2009) for early-stage broods. For late-stage broods, one-way ANCOVAs were performed to assess differences in reproductive traits among seasons during 2008. Furthermore, a two-way ANCOVA was performed between realized and effective FE with CL as covariable, to assess seasonal variability in egg loss during the spawning period. Interactions between categorical factors and the covariable were tested (CL × Season; CL × Stage). Since interactions were not significant, the interaction term was removed and models were refitted (homogeneity of slopes). Tukey's multiple comparison post-hoc test was performed when significant differences were found. For all the analyses performed, the obtained results were normal (Shapiro–Wilk test, P > 0.05) and homoscedasticity was met (Bartlett's test, P > 0.05). In order to determine temporal differences in the studied variables, data correction by size was performed on those for which it showed a significant effect. The effect of size was removed by standardizing data through linear regression between each dependent variable and size of S. arctus, calculated for the overall data set. Differences in average values of reproductive traits among broods for different developmental stages were assessed using Generalized Least Squares (GLS) to suitably model variance structure.
In a subsequent phase, generalized linear models (GLM) were used to analyse the variations of realized FE and the relative importance of the different variables, in order to develop a predictive model for FE during the reproductive cycle. The following predictive variables were considered: month of capture, CL and egg V. The study of residual distribution regarding the studied variables, previous to their insertion in the model, made it possible to determine a pattern for the variable Month, which was included in the model as a quadratic term. A first model was built where all the variables were included. In the process of variable selection (by backward elimination approach), Akaike's Information Criterion (AIC) and percentage of explained variation (deviance-D2) were used to evaluate the relative importance of each variable and so determine which variables were to be included in the final model and in which order. Likewise, the analysis of deviance to assess the significance (test F) of the variables included in the final model was carried out as a step by step process. The selection of the model, the most suitable link function and error distribution were based on residual distribution. A gamma distribution with a log link function was found to be the most suitable (Green et al., Reference Green, Gardner and Kennedy2009), which implies that the explanatory variables have had a multiplicative effect on the response parameter.
 $$\log \;\lpar {{\rm {\opf E}}\lpar {{\widehat{{{\rm FE}}}}_i} \rpar } \rpar = \eta _i = \beta _0 + \beta _1\;\log \,\;{\rm CL} + \beta _2\,{\rm Mont}{\rm h}^2 + \beta _3\,{\rm Month}\;$$
$$\log \;\lpar {{\rm {\opf E}}\lpar {{\widehat{{{\rm FE}}}}_i} \rpar } \rpar = \eta _i = \beta _0 + \beta _1\;\log \,\;{\rm CL} + \beta _2\,{\rm Mont}{\rm h}^2 + \beta _3\,{\rm Month}\;$$where the average FE 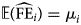 ${\rm {\opf E}}\lpar {{\widehat{{{\rm FE}}}}_i} \rpar = \mu _i$ for an observation i is estimated as
${\rm {\opf E}}\lpar {{\widehat{{{\rm FE}}}}_i} \rpar = \mu _i$ for an observation i is estimated as 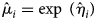 $\;\hat{\mu }_i = \exp \;\lpar \hat{\eta }_i\rpar $. All the analyses and model fitting have been performed using R.
$\;\hat{\mu }_i = \exp \;\lpar \hat{\eta }_i\rpar $. All the analyses and model fitting have been performed using R.
Results
Optimal egg sample size
The previous study of optimal sample size (250, 500, 750 and 1000 eggs) did not yield significant differences in average DWE among the different sample sizes used (ANOVA, F (3,333) = 0.372, P > 0.05). Nevertheless, for the purpose of this study, an optimal sample size of 750 eggs was selected, since samples of this size had a lower variability in the DWE obtained and, consequently, in the estimations of total number of eggs for each brood. The estimations carried out for the selected sample size showed coefficients of variation <3% in DWE determination (mean = 0.89%, 95% CI = 0.81–0.96%).
Reproductive effort
To estimate the realized FE of females of Scyllarus arctus, 149 individuals with a size range between 24.4 and 44.6 mm CL (34.5 ± 4.0 mm, mean ± SD) and a body wet weight between 11.8 and 71.9 g (35.2 ± 12.1 g) were used. The analysed broods showed a great variability both in weight, with a BWW between 0.76 and 7.03 g (2.74 ± 1.11 g) and a BDW between 0.32 and 2.50 g (1.05 ± 0.40 g), and in number of eggs, with 14,370–108,029 eggs (45,292 ± 16,265 eggs). The investment in brood weight relative to body weight of this lobster during the reproductive cycle ranged between 4.9% and 12.1% (7.7 ± 1.1%). Egg production per gram of body weight also showed great variation, ranging from 924.6–1742.2 eggs g−1 (1293.5 ± 175.5 eggs g−1).
Effective FE was determined for a size range similar to the one used for the determination of realized FE, between 28.5–44.6 mm (35.3 ± 4.4 mm, N = 65), and a body wet weight between 18.2–73.4 g (37.7 ± 13.4 g). The estimated number of eggs in broods in late egg stage ranged between 11,425 and 77,417 eggs (37,141 ± 16,587 eggs) for a BWW between 1.04–9.58 g (3.89 ± 1.82 g) and a BDW from 0.23–1.80 g (0.85 ± 0.39 g). The average number of eggs per gram of body weight determined for late broods was 987.7 ± 212.3 eggs g−1 (range: 551.9–1418.7 eggs g−1).
During embryogenesis, egg V increased by 22.2% on average, from 499 ± 62 mm3 10−4 (early eggs) to 610 ± 67 mm3 10−4 (late eggs). In regard to DWE, early eggs showed a great variation, ranging from 17.6–29.2 μg (23.2 ± 2.2 μg), while for eggs close to hatching, mean weight varied between 15.6–31.6 μg (22.4 ± 2.4 μg).
Influence of maternal size
Early-stage broods
Two-way ANCOVAs performed on brood size (FE and BDW) showed that female size, measured as CL, had a strong significant effect during the two years of study, accounting for >86% of the total variation (FE: F (1,147) = 1120.11, P < 0.001; BDW: F (1,147) = 1477.89, P < 0.001). The influence of CL on investment in brood size remained constant during the whole reproductive cycle (slope homogeneity). In general, the slopes of the adjusted allometric equations for brood size in each period ranged from 2.39–3.15, indicating the presence of a positive allometry (Figure 2). On the other hand, relative brood size (number of eggs per gram of body weight) did not show any relationship with lobster size (Pearson's r = 0.05, P > 0.05).

Fig. 2. Relationship between log realized fecundity (FE, number of eggs) and log brood dry weight (BDW, g) with body size (CL, mm) of a female Scyllarus arctus for winter samples (white dots, solid line), spring (black dots, solid line) and summer (grey dots, dashed line) for 2008 (top) and 2009 (bottom). Regression equation, coefficient of determination, significance and number of samples are indicated for each relationship corresponding to the season analysed.
Regarding individual egg traits, maternal size had a significant effect on DWE (7.2% of variance explained; F (1,147) = 29.07, P < 0.001). The relationship between DWE and CL exhibited positive allometry, determined by a mean estimated slope of 0.18 (95% CI = 0.08–0.29). On the other hand, egg V did not show any significant relationship with female size (F (1,147) = 2.06, P > 0.05), although it showed a positive correlation with DWE (Pearson's r = 0.55, P < 0.001).
Late-stage broods
Brood size (FE and BDW) in late-egg broods also showed a significant positive relationship with lobster size (FE: F (1,61) = 204.70, P < 0.001; BDW: F (1,61) = 279.74, P < 0.001). Variation in brood size and CL between seasons showed that variation in the number of eggs is mainly explained by the size of females (>75.17%). Two-way ANCOVA performed to compare realized and effective FE revealed a significant brood loss in S. arctus during incubation in the different periods analysed in 2008 (F (4,144) = 98.75, P < 0.001). Nevertheless, no differences were found between the slopes of the fitted lines (F (4,144) = 3.00, P > 0.05), indicating that egg loss during incubation remains constant throughout the analysed size range (Figure 3).

Fig. 3. Log-log relationship between realized fecundity (black dots, solid line) and effective fecundity (white dots, dashed line), and size of female Scyllarus arctus (CL, mm) for winter, spring and summer periods during 2008. Regression equation, coefficient of determination, significance and number of samples are indicated.
On the other hand, the influence of maternal size on DWE remained for eggs with developed larvae (17.21%; F (1,61) = 20.87, P < 0.001), with an average slope of 0.22 (95% CI = 0.07–0.37). For egg V, no significant relationship with size of individuals (P > 0.05) and DWE (Pearson's r = 0.10, P > 0.05) was found.
Temporal variations
Early-stage broods
The two-way ANCOVA analyses performed on FE (F (4,144) = 27.94, P < 0.01), BDW (F (4,144) = 10.66, P < 0.001), and DWE (F(4,144) = 60.89, P < 0.001), with CL as the covariate, showed a temporal pattern in reproductive traits. Main differences were found between the functions' intercepts for the different brood periods analysed (P < 0.001).
The average values of standardized realized FE in spring (48,116 ± 6314 eggs) were 18.0 ± 6.5% higher than FE of broods spawned in winter (40,815 ± 4428 eggs) in both years of the study, and 15.5 ± 0.8% higher than summer broods, with 41,660 ± 6163 eggs (Tukey's HSD, P < 0.001). The maximum differences in realized FE among broods were observed in 2009, when number of eggs in spring broods was 22.6% higher than in winter broods (Figure 4). On the other hand, standardized BDW decreased significantly at the end of the reproductive period by 10 ± 0.4% (mean summer BDW: 0.91 ± 0.14 g) with respect to broods spawned in winter (1.01 ± 0.11 g) and by 15.3 ± 0.1% with respect to spring (1.07 ± 0.12 g) in both years (Tukey's HSD, P < 0.01).

Fig. 4. Seasonal variations in brood and egg traits of Scyllarus arctus for both developmental stages in 2008 and 2009 (mean ± 95% CI). Mean values of fecundity (FE, number of eggs), brood dry weight (BDW, g), dry weight per egg (DWE, μg) standardized by size, and mean volume of eggs (mm3 10−4) are represented. Different letters indicate significant differences in brood traits (GLS, P < 0.01).
Investment in DWE was significantly higher in the first broods of the reproductive cycle, spawned during winter (24.7 ± 2.2 μg), than in broods spawned during spring and summer (Tukey's HSD, P < 0.001; Figure 4). However, this decrease in DWE was not significant between spring (22.4 ± 1.3 μg) and summer periods (21.8 ± 1.5 μg, Tukey's HSD, P > 0.05). DWE also showed significant differences between years of study with respect to season, DWE being higher in winter 2009 (25.5 ± 2.2 μg) than in 2008 (23.9 ± 1.9 μg; Tukey's HSD, P < 0.01). In contrast, average egg V was slightly higher in winter (514 ± 61 mm3 10−4) than in spring broods (503 ± 56 mm3 10−4) for the two years of study; however, this observed egg V was significantly reduced by 11.8 ± 0.7% in summer broods (451 ± 58 mm3 10−4, Tukey's HSD, P < 0.01).
Late-stage broods
Broods with eggs close to hatching also showed significant seasonal differences in FE (F (2,58) = 4.30, P < 0.05), BDW (F (2,58) = 10.74, P < 0.001) and DWE (F (2,58) = 15.15, P < 0.001), but not in egg V (P > 0.05). These reproductive traits showed a similar pattern to that observed in recent broods, although data showed a higher dispersion (Figure 4). Standardized effective FE in spring (36,889 ± 8901 eggs) was 21% higher than in summer broods (30,396 ± 8165 eggs; Tukey's HSD, P < 0.05); nevertheless, neither showed any difference with winter broods (33,454 ± 7662 eggs). Egg loss was evident in each season studied during 2008 (GLS, P < 0.001; Figure 4), showing an increase throughout the reproductive period, from 20.8% in winter broods to 23.0% in spring broods and 27.0% in summer broods (mean egg loss: 23.6 ± 3.2%). Effective investment in BDW showed a significant decrease in summer broods (0.63 ± 0.18 g) regarding winter and spring broods, with a mean BDW of 0.79 ± 0.14 g and 0.81 ± 0.18 g respectively (Tukey's HSD, P < 0.01; Figure 4).
DWE was also lower in summer broods (20.9 ± 1.3 μg) compared with winter (23.4 ± 1.7 μg; Tukey's HSD, P < 0.001) and spring broods (22.3 ± 1.2 μg; Tukey's HSD, P < 0.05). During the incubation period, eggs showed a significant increase in V, with an average of 25.4 ± 9.5% in the three seasons (GLS, P < 0.001; Figure 4), reaching an average egg V of 605 ± 90 mm3 10−4 for eggs in late stage.
Model for the estimation of realized FE
The variation in FE with female size and season demonstrates the importance of developing a model to predict FE during the reproductive period of S. arctus. For this analysis, data from years 2008 and 2009 were grouped since no significant differences in FE were found between years.
The model which only took female size into account explained 85.12% of variation in FE (Table 1), while the inclusion of month as a polynomial term accounted for an additional 5.87% of total deviance (90.98%). The model including all the variables showed the lowest AIC value, and all were found to be significant (P < 0.01). However, in spite of its significance (F = 10.37, P < 0.01, AIC = 2937), average egg V only accounted for an additional 0.60% of deviance (Table 1). In addition, even though AIC for this model was 7 units lower, Burnham & Anderson (Reference Burnham and Anderson2004) argue that there is no evidence of model improvement in cases like this. Therefore, based on these two criteria, a simpler model was selected, which did not include this variable. No evidence was either found of an interaction between female size and month of the reproductive cycle on the estimations of FE (F = 3.10, P > 0.05), thus the model proposed as the best one to assess realized FE was the one that included variables CL and Month (AIC = 2945), where all terms were significant (P < 0.001). Thus, the variable Month allowed an explanation of the relationship between FE and body size in this lobster, with a common slope of 2.91 (95% CI = 2.76–3.06) and a different intercept for each month during the annual reproductive cycle (Figure 5). Taking all this into account, the following expression is proposed for the prediction of FE in S. arctus during the reproductive period:
 $$\log \big( {{\rm {\opf E}}\lpar {{\widehat{{{\rm FE}}}}_i} \rpar } \big) = \eta _i = 0.064 + 2.907\,\log \,{\rm CL}-0.018\,{\rm Mont}{\rm h}^2 + 0.174\,{\rm Month}$$
$$\log \big( {{\rm {\opf E}}\lpar {{\widehat{{{\rm FE}}}}_i} \rpar } \big) = \eta _i = 0.064 + 2.907\,\log \,{\rm CL}-0.018\,{\rm Mont}{\rm h}^2 + 0.174\,{\rm Month}$$Table 1. Results of sequential adjustment of model for realized fecundity of Scyllarus arctus

Note: Columns represent degrees of freedom corresponding to the added term (Df), residual degrees of freedom for each model (Res. Df), deviance (D2), residual deviance (Res. D2), explained residual deviance (Exp. D2, %), P-value when a F test is used as significance test and AIC value.

Fig. 5. Monthly realized fecundity (FE, number of eggs) during the reproductive cycle of Scyllarus arctus (January–September). Dots represent estimated FE for each individual, while the greyscale shows the size class they belong to. Solid lines show predicted FE for the corresponding size (30, 35 and 40 mm; CL) during the spawning period, while dashed lines indicate 95% CI.
Discussion
The presence of females with broods showing eggs in early and late stages in all the seasons analysed in this study is in concordance with the extended spawning period shown by Scyllarus arctus in the Mediterranean, which practically encompasses the whole year, except for autumn, a season during which a peak moult could take place prior to the start of a new reproductive period (Vilotte, Reference Vilotte1982; Mura et al., Reference Mura, Cau and Deiana1984; Cau et al., Reference Cau, Davini, Deiana and Salvadori1988).
Estimations of realized FE on the Galician coast (14,370–108,029 eggs, 24–45 mm CL, N = 149) were higher than previous estimations in the Mediterranean (30,000–70,000 eggs, 28–34 mm CL, N = 4; Vilotte, Reference Vilotte1982). These geographic differences are partly due to methodological factors such as the narrow size range used and the number of females studied (Currie & Schneider, Reference Currie and Schneider2011). In this study, the smallest females used were close to the size of sexual maturity (25.7 mm CL, Alborés et al., 2019), while the largest ones were over 45 mm CL (180 mm in total length), which is larger than the maximum length cited for this lobster (160 mm in total length; Relini et al., Reference Relini, Bertrand and Zamboni1999).
In general, the reproductive potential of female S. arctus has shown a strong positive correlation with the size of individuals (CL), which has also been described in other slipper lobsters (Oliveira et al., Reference Oliveira, Freire and Bertuol2008), and other crustaceans (Hines, Reference Hines1982; Attard & Hudon, Reference Attard and Hudon1987; Fischer et al., Reference Fischer, Thatje and Brey2009). Egg production in scyllarids is a power function of body size, although the type of function described can vary among populations and species of lobsters (DeMartini & Williams, Reference DeMartini and Williams2001). In the specific relationships found between size-FE and size-brood weight in S. arctus, the observed slope for all relationships was not significantly different from a theoretical slope of 3 (t-test, P > 0.05), indicating a volumetric constraint to egg storage related to female size (Somers, Reference Somers, Wenner and Kuris1991). Although female body size (CL) is the most important determinant of FE, other factors also contribute to the total variance. The influence of reproductive senescence (or reduced FE with age) was noted in large-sized females of some spiny lobsters (Chubb, Reference Chubb, Phillips and Kittaka2000). MacDiarmid & Kittaka (Reference MacDiarmid, Kittaka, Phillips and Kittaka2000) suggest another reason: selective fishing of larger males. A male's capacity to fertilize the whole brood of a female mainly depends on its body size and, relatedly, on the amount of sperm it can produce. This limitation reaches a higher relevance in species with external fertilization and which do not accumulate spermatophores (MacDiarmid & Butler, Reference MacDiarmid and Butler1999), as is the case of this slipper lobster. However, as relative brood size showed no relation to body size in S. arctus, there was no evidence that would support such an hypothesis. This is consistent with the fact that it is a highly exploited population, and larger (and older) individuals are quickly eliminated from the population.
The positive influence of maternal size on egg weight (DWE) has also been observed both in clawed lobsters such as Homarus gammarus (Linnaeus, 1758) (Tully et al., Reference Tully, Roantree and Robinson2001; Agnalt, Reference Agnalt2008) and H. americanus (H. Milne Edwards, 1837) (Ouellet & Plante, Reference Ouellet and Plante2004), and in spiny lobsters such as Palinurus elephas (Fabricius, 1787) (Goñi et al., Reference Goñi, Quetglas and Reñones2003) and Jasus edwardsii (Hutton, 1875) (Smith & Ritar, Reference Smith and Ritar2007), although this effect has not been previously described in scyllarid lobsters. There are several plausible explanations for this positive relationship in the life history theory: (i) density-dependent survival or competition due to an increase in brood size with female size (Parker & Begon, Reference Parker and Begon1986); (ii) maternal phenotype-specific optimal egg sizes related to size-dependence in habitat selection (Hendry et al., Reference Hendry, Day and Cooper2001); (iii) physiological or morphological constraints on egg size imposed by the size of the mother (Fox & Czesak, Reference Fox and Czesak2000); (iv) size- or age-dependence in female provision to their offspring (Sakai & Harada, Reference Sakai and Harada2001; Kindsvater et al., Reference Kindsvater, Bonsall and Alonzo2011). However, there is no evidence to support these theories for marine decapods, and further research is needed to determine the source of this maternal effect.
In addition to the effect of size of individuals, seasonal variations observed in brood traits during the long spawning period in both years of study could reflect the existence of a reproductive strategy. Our data revealed that seasonal changes in reproductive traits follow a cyclical pattern, reflecting the inverse relationship between values of FE and DWE in the two first seasons of the cycle, with a maximum egg weight in winter and a maximum number of produced eggs in spring. The seasonality in brood traits observed in this scyllarid is in accordance with observations in other species of crustaceans with a long reproductive period, such as Maja brachydactyla (Balss, 1922) (Verísimo et al., Reference Verísimo, Bernárdez, González-Gurriarán, Freire, Muiño and Fernández2011), Panulirus inflatus (Bouvier, 1895) (Garcia, Reference Garcia1985), P. argus (Latreille, 1804) (Fonseca-Larios & Briones-Fourzan, Reference Fonseca-Larios and Briones-Fourzan1998), Crangon crangon (Linnaeus, 1758) (Oh & Hartnoll, Reference Oh and Hartnoll2004; Urzúa et al., Reference Urzúa, Paschke, Gebauer and Anger2012), and Cancer setosus (Molina, 1782) (Fischer et al., Reference Fischer, Thatje and Brey2009). Nevertheless, in S. arctus this pattern does not continue in summer, when a lower reproductive investment in broods is observed, both in terms of total energetic investment and in terms of individual characteristics of eggs within them (DWE and V). This decrease in investment in the final broods of the reproductive cycle, also observed in other lobsters, is related to the capacity to carry out successive spawnings (Chubb, Reference Chubb, Phillips and Kittaka2000; Vijayakumaran et al., Reference Vijayakumaran, Murugan, Remany, Leema, Kumar, Santhanakumar, Venkatesan and Ravindran2005; Chang et al., Reference Chang, Sun, Chen, Yeh and Chiang2007). In this way, the capacity of S. arctus females to spawn several times during a reproductive period (up to three subsequent broods in captivity, Alborés et al., Reference Alborés, García-Soler and Fernández2019), in terms of energetic investment in broods, might be conditioned by their nutritional status (Wen et al., Reference Wen, Chen, Zhou, Ai and Deng2002; Verísimo et al., Reference Verísimo, Bernárdez, González-Gurriarán, Freire, Muiño and Fernández2011). Furthermore, the balance of energy available for growth and reproduction (see Hartnoll, Reference Hartnoll and Wenner1985), which bears more importance in species with indeterminate growth (which continue to grow after sexual maturity), as in S. arctus, might contribute to the decrease in reproductive investment in the last broods produced during summer.
In marine crustaceans, seasonal differences found regarding brood traits have also been related to environmental factors such as temperature, food availability and day length (Pond et al., Reference Pond, Harris, Head and Harbour1996; Fischer et al., Reference Fischer, Thatje and Brey2009; Urzúa et al., Reference Urzúa, Paschke, Gebauer and Anger2012). On the Galician coast, seasonal variation in egg traits in S. arctus might be related to the variation in oceanographic conditions caused by the occurrence of upwelling of nutrient-rich deep waters (Alvarez et al., Reference Alvarez, Gomez-Gesteira, DeCastro, Lorenzo, Crespo and Dias2011). Thus, broods showing a higher investment per egg are produced during winter (December–February), when water temperature is lower and plankton is less abundant; while the production of smaller eggs takes place during spring (April–May) and summer (July–September), when water temperature and plankton densities are either rising or show higher values (Álvarez et al., Reference Álvarez, Lorenzo and DeCastro2012). Nevertheless, this explanation about an adaptive response to environmental changes is difficult to validate, since available data are limited to a two-year-long series.
The seasonal pattern and maternal size effect observed in DWE were similar to that of broods with eggs close to hatching, thus confirming the persistence of traits from early eggs to those in the final stages of their incubation period. Egg V and DWE are widely used to determine investment in offspring in marine invertebrates; nevertheless, variation in egg V within a species may not be related to variation in parental investment (see Clarke, Reference Clarke1993). In marine decapods, DWE is a good approximation of offspring size (Giménez & Anger, Reference Giménez and Anger2003; Moland et al., Reference Moland, Moland Olsen and Stenseth2010; Diez & Spivak, Reference Diez and Spivak2012; Urzúa & Anger, Reference Urzúa and Anger2013) and predictor of the physiological condition of larvae, which affects the capacity of newly hatched larvae to survive in highly variable environmental conditions (Schuh & Diesel, Reference Schuh and Diesel1995; Hancock et al., Reference Hancock, Hughes and Bunn1998; Paschke et al., Reference Paschke, Gebauer, Buchholz and Anger2004). Therefore, observed differences in average DWE in S. arctus could represent actual differences in larval size or energetic condition. However, further research is needed in order to determine how this variation in DWE translates to offspring in this slipper lobster.
The presence of this intraspecific variability in FE during the reproductive period, is evidence of the importance of the development of an optimal predictive relationship between number of eggs produced and body size. The advantage of using GLM with respect to classical linear models lies in the fact that it provides estimations in the original measuring scale, thus removing interpretive difficulties associated with subsequent transformations and generating a useful tool for stock assessment (Green et al., Reference Green, Gardner and Kennedy2009). In addition, the insertion of a variable with a higher temporal resolution (month) allowed the final model to account for more variation. The great percentage of variation explained by this model (>90%) indicates its potential use for the estimation of FE during the annual reproductive cycle (January to September).
The model obtained for the determination of FE in S. arctus suggests the need to protect larger females, with the highest FE, and which are not protected by current legislation. However, it is worth mentioning that the relative contribution to population egg production by any size class is a function both of frequency of spawning and of the proportion of the population it represents (Aiken & Waddy, Reference Aiken, Waddy, Cobb and Phillips1980). The determination of these parameters and the constant egg loss rate (24%) shown by S. arctus, will allow for the calculation of more accurate annual egg production indices and an improvement in estimations of relative reproductive potential.
The results of this research have direct implications for fisheries management of S. arctus and other commercial crustacean species with similar reproductive strategies. The development of a fisheries management plan should take into account both seasonal and maternal size effects on investment per egg. Since variations in egg weight affect the initial larval stage, as well as later phases of life history of marine decapods (Jørstad et al., Reference Jørstad, Prodöhl, Kristiansen, Hughes, Farestveit, Taggart, Agnalt and Ferguson2005; Giménez, Reference Giménez2006; Bas et al., Reference Bas, Spivak and Anger2008; Guay et al., Reference Guay, Sainte-Marie and Brêthes2011), this trait could have implications both in future recruitment success and in population recovery of S. arctus.
Acknowledgements
The authors would like to thank the skipper of the Malpica fishing boat ‘Venanciño’ for his invaluable help with fieldwork.
Financial support
This study is part of the project ‘Study of the reproductive biology and larval development of the small European locust lobster Scyllarus arctus: basic knowledge for a Sustainable Fishery Management’ (07MMA009103PR) co-funded by the Xunta de Galicia and the European Regional Development Fund (ERDF).







 Open access
Open access