



Introduction
Reliable documentation of the characters and knowledge of the diversity within a fossil group is needed to determine the relationships and phylogeny of species in that group. This is not available for late Paleozoic pinnids because of limited study, incomplete taxonomic descriptions, and unjustified reconstructions of pinnid species of that age. From the beginning of study of late Paleozoic pinnid species, descriptions were influenced by assumptions of character states when such features were not clearly observed. Subsequently, these inaccuracies became incorporated into the genus concept of first-described genera and were perpetuated in later works. Current examination of fossil collections and literature illustrations reveals a diversity of species present in late Paleozoic strata, but the ability to define stable taxonomic groups is hampered by inaccurate descriptions and loss of types.
The troubles build from the start of descriptive work on late Paleozoic pinnids (Münster, Reference Münster1839), with the report on an incomplete specimen questionably identified as a pinnid and described with few characters and poor illustration. A series of missteps in subsequent redescription led to reports on the presence of shell characters that are not present in other specimens from the source strata. A redescription (Geinitz, Reference Geinitz1861) introduced the concept of a raised, subterminal beak on the shell, which is a departure from the basic design of younger pinnids.
Understanding early pinnid bivalve relationships became more complicated with the synonymy (Hind, Reference Hind1901; Cox and Hertlein, Reference Cox, Hertlein and Moore1969) of Aviculopinna with Pteronites—a genus with subterminal beak position and anterior and posterior wings. These characters are typical of species in the family Pterineidae, blurring the distinction between the Pterineidae and the Pinnidae. The synonymy of Aviculopinna with Pteronites by Cox and Hertlein (Reference Cox, Hertlein and Moore1969) was accompanied by a definition of the synonymized genus as having a subterminal beak. This led to the adoption of a hypothesis that beak position in pinnid species is a significant factor in determining pinnid relationships. All living pinnids have entirely terminal beaks, so the occurrence of subterminal beaks like that of Pteronites meant the existence of a group separated from the terminal beak pinnids and potentially members of a different family.
Uncertainty and confusion about relationships that extend from species to family have to be resolved by critical examination of all work on the subject, starting with the first publications. This centers on the status of genus Aviculopinna and species descriptions presented in the 1800s. This report presents the results of study of that genus and the two species assigned to it that occur in strata of the Zechstein Group (late Permian) in north-central Europe. Some specimens of both species have been located that were examined by their describers, supplemental material has been located, and specimens have been recollected from type localities. The results are clear enough to clarify the status of species and genus and to provide enough data to resolve questions of family relationship and status within the Pteriomorphia.
Previous work
The source of confusion about Aviculopinna arises from selecting a type species that has a confusing description, resulting in a vague genus concept and indiscriminate application of the genus name. The genus name Aviculopinna was established by Meek (Reference Meek1864) as a genus name for the late Paleozoic bivalve species pinnaeformis, which Meek considered to be a member of the family Pinnidae, a determination indicated by written statement in the text and by inclusion of “pinna” in the name of the new genus. The species pinnaeformis (Geinitz, Reference Geinitz1848) is the type species of Aviculopinna by monotypy. The name pinnaeformis was introduced by (Geinitz, Reference Geinitz1848) when renaming a specimen identified as Pinna? prisca by Münster (Reference Münster1839) (Fig. 1), a species name given (Münster in Goldfuss, Reference Goldfuss1837) to a different bivalve of the Keuper (Triassic). Geinitz (Reference Geinitz1861) later re-assigned the species to the genus Avicula and provided a longer description of the species, accompanied by drawings of four specimens (Fig. 2). Meek (Reference Meek1864) placed the species and new genus Aviculopinna in the family Pinnidae—a genus assignment the same as that made by Münster (Reference Münster1839).

Figure 1. Münster (Reference Münster1839) illustration of Pinna? prisca (=A. pinnaeformis) from the lower shale cycle (z1, Kupferschiefer, T1), Zechstein Group, Wuchiapingian Stage, Lopingian, Permian, Gera, Germany. Width of posterior margin is 28 mm (Geinitz, Reference Geinitz1848).
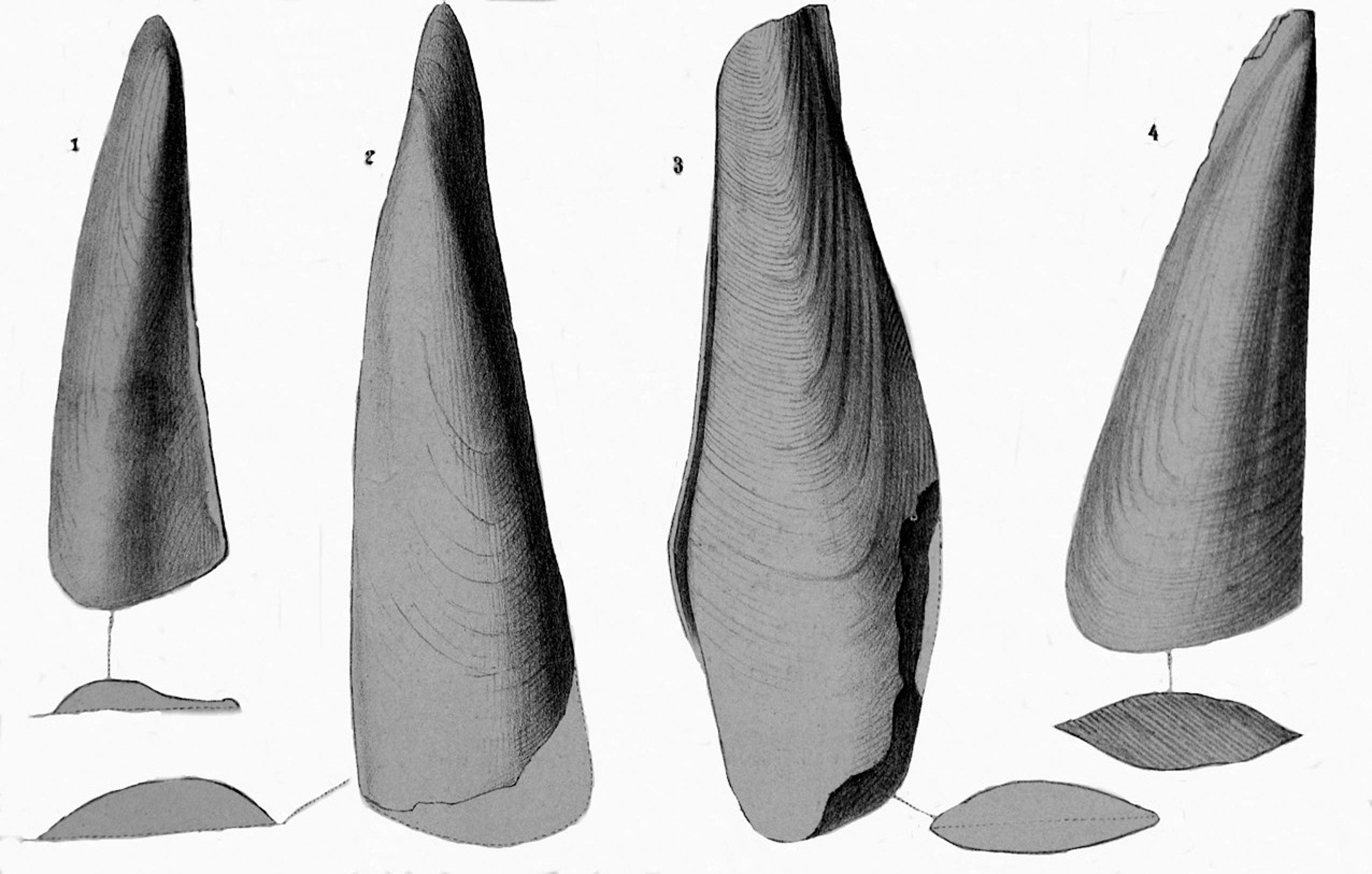
Figure 2. Geinitz (Reference Geinitz1861) illustration of Avicula pinnaeformis specimens from the Werra Formation (z1), Zechstein Group, Wuchiapingian Stage, Lopingian, Permian, Gera and Rückingen, Germany. Length of specimen 4 is ~6 cm.
The written description of Münster (Reference Münster1839) is too limited for accurate species identification and the Geinitz (Reference Geinitz1861) written description is problematic for mentioning characters not recognized by other workers. By stating that the relationship of Pinna? prisca to Avicula anomala Sowerby is “unmistakable,” Geinitz (Reference Geinitz1861) revealed that he believed the Zechstein fossil to be similar to a species of Pteria (=Avicula) of a type now placed in the family Pterineidae, not in the Pteriidae or Pinnidae. In addition to unclear description, the Geinitz (Reference Geinitz1861, pl. 14, figs. 1–4) illustration of a group of specimens shows too much variation among specimens to determine a reliable circumscription for the genus or to determine a solid family assignment for the genus. The Geinitz illustrations were drawn to show a substantial anterior wing and a flattened anterior byssal flange separate from the inflated portion of the valve, characters not recognizable in well-preserved specimens of Zechstein Aviculopinna.
Translations of the original German species
Original German species descriptions are translated here, using the style of 1800s German language, with some words in brackets inserted to provide clarity.
Münster (Reference Münster1839, p. 45, P.? prisca, pl. 4, fig. 4).—“from the Kupferschiefer with spotted galena of the Merzenberg [Merzen Hill] near Milbitz, not far from Gera. This fossil coming nearest to a Pinna from its outline and striation was presented together with the former [a brachiopod] by its owner, Mr. Laspe, to the Geognostic Section of the meeting of natural scientists in Jena. Specimens presented of this dubious fossil are upper and lower impressions [external molds of valves]. On the margins longitudinal striae are present, cut by concentric lines. The shell is extremely thin.”
Geinitz (Reference Geinitz1848, p. 8, Solen? pinnaeformis).—Replacement name proposed by Geinitz without illustration: “According to Münster's figure this very slightly convex mussel [bivalve] is very elongated and widens gradually posteriorly, where it is truncated with rounded angles. The small umbo is situated at the anterior end, which is obliquely truncated [it is broken], in such a way that the lower edge of the shell forms a projection. The length estimates 100 mm, the height [dorsal-ventral width] at the umbo/beak is 11 mm, near the posterior margin is 28 mm.” “The specimen comes from the Kupferschiefer of the Merzenberg [Merzen Hill] near Gera between Milbitz and Thieschitz.”
Geinitz (Reference Geinitz1861, p. 77, Avicula pinnaeformis, pl. 14, figs. 1–4).—A later redescription: “If the figures of this contribution are compared with Avicula anomala Sowerby, their relationships are unmistakable. Missing teeth along the hinge margin do not allow placement in Gervillia. The up to 100 mm long shell has a long wedge-shaped outline, as it prolongs unusually from anterior to posterior and broadens very slowly (up to 28 mm). At the posterior, mostly gaping end it is obliquely truncated, meeting the hinge margin at an obtuse angle and with a rounded edge with the lower margin. The small, not raised umbo/beak, is situated near the anterior end of the shell. Anterior to the umbo/beak a wing is developed, more or less pronounced with a rounded margin. This [wing] was broken off in the specimen from the Laspe collection figured by Münster and visible only from its external mold. The posterior wing, which separates the linear hinge line from the vaulted [convex] part of the shell, is very long and narrow. From the umbo/beak in posterior direction lines [striae] are developed, crossed by concentric growth lines. Frequently, the former appears very faint and indistinct, the latter become pronounced near the lower margin and sometimes merge to wrinkles. A slight bay [sinus] behind the anterior wing in the right valve is intended for the exit of the byssus.” “Occurrence: In the Kupferschiefer at the Merzenberg [Merzen Hill] between Milbitz and Thieschitz as well as near Trebnitz near Gera. In the lower Zechstein from Thieschitz, Roschitz, Lutzschethal, Trebnitz, Lasen in the vicinity of Gera, Seissla near Rahnis in Thuringia, and in the upper Zechstein from Rückingen in the Wetterau [Hesse]; as a great rarity also in the Shell limestone of Tunstall Hill near Sunderland [England].”
Discussion of previous works
When reading the original German descriptions it should be noted that the source of these specimens, listed as Kupferschiefer, refers to their occurrence in the basal unit of the Zechstein Group in central Germany, not to an occurrence in sediment deposited in an anoxic environment. The presence of benthic pinnid bivalves indicates presence of oxygenated ocean waters in a shallow-water marine facies of the Kupferschiefer (Peryt et al., Reference Peryt, Hałas and Hryniv2010b).
The progression from Münster (Reference Münster1839) to Geinitz (Reference Geinitz1861) resulted in major differences in illustrations of the species. The Münster (Reference Münster1839) illustration shows a triangular form with straight margins and an indication of radial features, whereas the Geinitz (Reference Geinitz1861) illustrations show an elongate form with rounded growth lines and rounded ventral margin (on three of four specimens) and distinct subterminal beak position with prominent anterior wing projection and raised umbones. The Geinitz (Reference Geinitz1861) report is the first time that mention of an anterior wing is used in describing the species.
The presence of a substantial subterminal beak, prominent umbo, and anterior wing are features incompatible with known pinnids. When describing the American species americana (now placed in Meekopinna), Meek (Reference Meek1872) regarded the subterminal beak characterization of type species pinnaeformis as an exaggeration, but the concept of a subterminal beak with an anterior wing was adopted by de Koninck (Reference de Koninck1885) in his monograph of European Carboniferous bivalves. Subsequent discussion by Hyatt (Reference Hyatt1892), Hind (Reference Hind1901), and Girty (Reference Girty1915) commented on the inability to verify the presence of a distinct anterior wing in late Paleozoic pinnids. The controversy remained unresolved because the apex of late Paleozoic pinnids is seldom preserved and when present is very rarely preserved with quality adequate for showing the position of the prodissoconch.
Subsequent study of Aviculopinna by Langenhan (Reference Langenhan1899 [1900], Reference Langenhan1908) and Riedel (Reference Riedel1917) of Permian pinnids from localities in Poland resulted in recognition of the species neukirchensis (Fig. 3), a species closely related to the type species pinnaeformis, and those authors mentioned the similarity of the new species to specimen 4 on plate 14 of Geinitz (Reference Geinitz1861). The Polish specimens described by Langenhan provided additional documentation of the adult shell morphology of Aviculopinna but did not add data about the apex or location of the beak on the shell. Later studies (Trechmann, Reference Trechmann1944; Logan, Reference Logan1967; Paul, Reference Paul, Harwood and Smith1986; Raczyński, Reference Raczyński1997; Biernacka et al., Reference Biernacka, Borysiuk and Racyński2005; Brandt, Reference Brandt2017) reported on the occurrence of Aviculopinna in strata of the Zechstein basin, but provided little further documentation on characters of the species or genus. Current study of German and Polish collections of Aviculopinna species from basal Zechstein strata now provides enough data to produce a circumscription of the genus that establishes a solid foundation for study of late Paleozoic pinnids. Aviculopinna can best be defined as the group of characters common to the two closely related species.

Figure 3. Langenhan (Reference Langenhan1908) illustration of Aviculopinna neukirchensis from the Werra Formation (z1), lower carbonate cycle (Ca1), Zechstein Group, Wuchiapingian Stage, Lopingian, Permian, Nowy Kościół (Silesia), Poland. Length of largest specimen is ~7 cm.
Materials and methods
This study redescribes genera and species, based on 10 specimens from the Gera Museum für Naturkunde, Gera, Germany, including two (MNG-8529 and MNG-8530) illustrated by Geinitz (Reference Geinitz1861) as pinnaeformis; two specimens (YPM-507884 and YPM-507885) from the Yale Peabody Museum, Connecticut, USA, identified by Hanns B. Geinitz as pinnaeformis and sent to the Yale Peabody Museum, Connecticut, USA by Geinitz before 1868 (written communication, Susan Butts and Jessica Utrup, 9 June 2020); eight specimens of from the paleontological collections of the Museum Uniwersytet Wrocławski, including two specimens collected by Alwin Langenhan and illustrated by Langenhan (Reference Langenhan1908); ~30 specimens collected for this study by PR, now located in the collections of the Non-Vertebrate Paleontology Laboratory, University of Texas, Austin, Texas, USA; ~10 specimens from the collection of SB; three specimens from NW Hesse (coll. H. Kaufmann), and two specimens from Hasbergen (Lower Saxony) in the collections of the German Geological Survey (Bundesanstalt für Geowissenschaften und Rohstoffe). Specimens of Pteronites angustatus M'Coy, Reference M'Coy1844, type species for Pteronites and part of the M'Coy collection of the National Museum of Ireland (NMING), are redescribed to evaluate the basis of synonymizing Aviculopinna with Pteronites.
The location of specimens of Aviculopinna pinnaeformis collected at Gera, Germany, is 50.8971°N, 12.0730°E, and specimens of Aviculopinna neukirchensis collected from Nowy Kościół, Poland is 51.0743°N, 15.8660°E.
Repositories and institutional abbreviations
Muzeum Geologiczne Uniwersytetu Wrocławskiego/Geological Museum of the University of Wrocław, Wrocław, Poland (MGUWr); Museum für Naturkunde Gera/ Gera Natural History Museum, Gera, Germany (MNG); Non-Vertebrate Paleontology Laboratory, University of Texas, Austin, Texas, USA (NPL); Yale Peabody Museum, Yale University, New Haven, Connecticut, USA (YPM); National Museum of Ireland, Dublin, Ireland (NMING).
Systematic paleontology
Family Pinnidae Leach, Reference Leach1819
Genus Aviculopinna Meek, Reference Meek1864
Type species
Solen? pinnaeformis Geinitz, Reference Geinitz1848 (=Pinna? prisca Münster Reference Münster1839, a replacement name), by original designation (Meek, Reference Meek1864, p. 212), from the late Permian of Germany.
Diagnosis
Elongate narrow shell with blunt, rounded apex; edentulous; linear dorsal and ventral margins; low apical angle on mature shell; well-developed dorsomarginal fold for ligament; ridged growth lines that are not raised into lamellae; blunt, semi-linear posterior margin; ventral margin not thickened (no byssal flange); growth lines meet the hinge line at vertical or obtuse angles on the mature shell; shallow subdorsal radial groove.
Occurrence
Europe, North America: known from Zechstein Basin of northern and central Europe and western North America; Lopingian, Permian.
Remarks
The growth line configuration of Aviculopinna and shallow subdorsal radial groove provide the most distinctive features for identification. Young juvenile shells have a blunt, rounded posterior margin that changes within a half centimeter to linear posterior growth lines that meet the hinge line at a nearly 90° angle. On later growth of the mature shell, the posterior margin is straight and linear and growth lines meet the hinge line with an obtuse angle. This results in the greatest shell length occurring on the posteroventral corner of the shell.
Aviculopinna pinnaeformis (Geinitz, Reference Geinitz1848)
Figure 4
- non Reference Goldfuss1837
Pinna prisca Münster in Goldfuss, p. 164, pl. 127, fig. 2.
- Reference Münster1839
Pinna? prisca; Münster, p. 45, pl. 4, fig. 4.
- Reference Geinitz1848
Solen? pinnaeformis Geinitz, p. 8.
- Reference Geinitz1857
Avicula pinnaeformis Geinitz; Geinitz, p. 210.
- part Reference Geinitz1861
Avicula pinnaeformis Geinitz; Geinitz, p. 77; non pl. 14, figs. 1–4.
- Reference Eisel1862
Avicula pinnaeformis Geinitz; Eisel, p. 29.
- non Reference Geinitz1866
Avicula pinnaeformis Geinitz; Geinitz, p. 31, pl. 2, fig. 13 [=Aviculopinna americana Meek, 1867].
- Reference Zittel1885
Aviculopinna prisca Münster; Zittel, p. 45.
Neotype
MNG-8528 and MNG-8527, part and counterpart, designated herein (Fig. 4.1). Consultation with the Urwelt-Museum Oberfranken, Bayreuth, reports that the name-bearing type specimen of Pinna? prisca of Münster (Reference Münster1839) is not present in their collections and is lost. When this species name was replaced by pinnaeformis, the first revising author, H.B. Geinitz (Reference Geinitz1848, Reference Geinitz1861), did not select a type from the collections of specimens available at that time nor did any subsequent author. Although the neotype selected here was not figured by Geinitz (Reference Geinitz1861), it represents the most common form of the species present in Kupferschiefer strata at Gera. It is closest to the specimen illustrated by Geinitz (Reference Geinitz1861, pl. 14, fig. 4), which is a specimen from Rückingen, Hesse, that was selected by later workers (Langenhan, Reference Langenhan1899 [1900], 1908; Riedel, Reference Riedel1917) as representative of pinnaeformis. Selecting this specimen as neotype preserves the traditional concept of the species, and it is from the type locality. The neotype is kept in the Museum für Naturkunde Gera, Germany. The type stratum and type locality is Werra cycle (z1), Kupferschiefer (T1) (zWTu), Werra Formation, Zechstein Group, from Trebnitz near Gera, Thuringia, Germany; Wuchiapingian Stage, Lopingian, Permian.
Diagnosis
Elongate equivalved shell with straight dorsal and ventral margins on mature shell; edentulous; valves moderately inflated; projecting posteroventral margin on mature shell; posterior shell margin linear; ventral margin growth lines turn upward in small curve toward hinge line, meeting the hinge line at a vertical to obtuse angle on young and mature shell; subdorsal shallow, radial groove present close to hinge line; well-developed dorsomarginal fold for ligament; ventral portion of shell has ridges of thickened growth lines enclosing finer growth lines that may continue up to hinge line; apical angle on mature shell 20°; beak not preserved.
Occurrence
Gera and Rückingen, Germany, Werra Formation (z1), Zechstein Group, Wuchiapingian Stage, Lopingian, Permian. Collected from sediments deposited in a shallow marine, oxygenated ocean-water environment.
Remarks
This description is based on specimens MNG-8527, -8528, and -8552. The neotype (MNG-8528/-8527) is a 6 cm long specimen consisting of both valves compressed together as part and counterpart. Growth lines turn upward on the posteroventral corner, meet the hinge line at a vertical to obtuse angle on mature shell, but have a high acute angle on the first 2 cm of shell. The anterior portion of shell has pronounced curvature on the first 1 cm of growth. These specimens are similar to the specimen illustrated by Münster (Reference Münster1839, pl. 4, fig. 4) and are preserved in clay-rich matrix characteristic of the basal Zechstein of the area around Gera. They are dissimilar to specimens of the genus present in the overlying carbonates. This species can be distinguished from A. neukirchensis by its more prominent subdorsal radial groove, larger size, greater inflation, and greater length before growth lines change from acute angle intersection with the dorsal margin (hingeline) to an obtuse angle.
The drawings of Geinitz (Reference Geinitz1861) neither accurately show the characteristic shape of the growth lines nor the well-developed dorsomarginal fold. The mention of anterior and posterior wings on the shell is inappropriate. The blunt anterior end is evenly rounded. There is no posterior wing on a shell where the posteroventral corner of the shell projects farther than the hinge line (dorsoventral corner). The written mention of striae crossed with growth lines is confusing because the only radial line crossing growth lines is the single shallow groove near the hinge line. The shallow radial groove present below the hinge line, recognizable on most Gera specimens, is not mentioned in the previous study, although it is shown in Geinitz (Reference Geinitz1861, pl. 14, fig. 4). A slight gap for the byssal opening is normal for pinnids, but it is seldom developed enough to be seen as a distinct character and is not observed on the shells examined.
There is no confirmed occurrence of Aviculopinna in British Zechstein deposits. Logan (Reference Logan1967) reported that, despite previous reports of the genus in England, no specimens can be located to confirm an occurrence. Hollingworth and Pettigrew (Reference Hollingworth and Pettigrew1988) did not mention any pinnids from the reef deposits of the British Zechstein. No specimens have been found in recent examinations of Zechstein strata in England (SB, personal observation).
Aviculopinna neukirchensis (Langenhan, (Reference Langenhan1899 [1900])
Figures 3, 5–9
- Reference Langenhan1899 (1900)
Pinna neukirchensis Langenhan, p. 48 [pl. 2, figs. 13–15 published in offprints only].
- Reference Langenhan1908
P.[inna] neukirchensis Langenhan, p. 123, pl. 3, figs. 1–8.
- Reference Riedel1917
Aviculopinna prisca Münster; Riedel, p. 72.
- Reference Eieentraut1939
Pinna neukirchensis Langenhan; Eisentraut, p. 92.

Figure 4. Aviculopinna pinnaeformis in the Museum für Naturkunde Gera from basal clay-rich strata of the lower Werra Formation (z1), Zechstein Group, Wuchiapingian Stage, Lopingian, Permian, Gera, Germany. (1) Neotype (MNG-8528, right valve, mostly flattened); (2) MNG-8552, right valve view with steinkern, mostly flattened. Scale bars are 2 cm.

Figure 5. Aviculopinna neukirchensis from clay-rich mudstone and carbonate strata of the lower carbonate cycle (Ca1), Zechstein Group, Wuchiapingian Stage, Lopingian, Permian, Nowy Kościół (Silesia), Poland. (1) Lectotype (MGUWr-1780s/1); (2) prodissoconch and nepioconch of juvenile (NPL-0003); (3) juvenile specimen with blunt apex (NPL-0001). Scale bars are (1, 3) 2 cm; (2) 1 mm.

Figure 6. Aviculopinna neukirchensis in life position in calcareous cemented layer within mudstone strata, lower carbonate cycle (Ca1), Zechstein Group, Wuchiapingian Stage, Lopingian, Permian, Nowy Kościół (Silesia), Poland. Scale bar is 2 cm. This photo appears in Peryt et al. (Reference Peryt, Geluk, Mathiesen, Paul, Smith, Doornenbal and Stevenson2010a).

Figure 7. Aviculopinna neukirchensis shells, bedding plane group, lower carbonate cycle (Ca1), Zechstein Group, Wuchiapingian Stage, Lopingian, Permian, Nowy Kościół (Silesia), Poland.

Figure 8. Aviculopinna neukirchensis, paralectotype, MGUWr-1780s/2, lower carbonate cycle (Ca1), Zechstein Group, Wuchiapingian Stage, Lopingian, Permian, Nowy Kościół (Silesia), Poland, Langenhan collection. An example of a radial array of growth line ridges shown in illustrations of Langenhan (Reference Langenhan1908). Scale bar is 2 cm.

Figure 9. Shell microstructure of A. neukirchensis, showing thick outer columnar prismatic calcite shell layer (left) marked with growth increment bands, and a thin, inner shell layer (right) of aragonite recrystallized to calcite, lower carbonate cycle (Ca1), Zechstein Group, Wuchiapingian Stage, Lopingian, Permian, Nowy Kościół (Silesia), Poland. Scale bar is 1 mm.
Types
MGUWr-1780s/1 (lectotype designated here) and MGUWr-1780s/2 (paralectotype), University of Wrocław, Poland
Diagnosis
Elongate shell with slender trapezoid shape; blunt, rounded apex; edentulous; linear dorsal and ventral shell margins; apical angle of 10–15° on mature shells; moderate shell inflation on young shells; well-developed dorsomarginal fold for ligament; narrow, shallow radial groove on shell near hinge line; distinct, close-spaced growth lines irregularly spaced with some grouping into bundles, suggesting episodic growth variation; posterior growth lines curve gradually up to hinge line on small juvenile shells; at distance of ~2 cm from apex the growth lines turn upward from ventral in a tight bend on the posteroventral margin and change to linear posterior margin that meets hinge line at a vertical to obtuse angle of as much as 120°; shell inflation of juvenile shell with shallow curvature of ~1:3 to 1:4, and inflation index of ~0.33; minor angulation may be present near midline of valve of mature shell; prodissoconch situated on hinge line and not raised; anteroventral shell growth (anterior of prodissoconch) of 2 mm, forming blunt apex; ventral margin characterized by raised ridges but no byssal flange.
Occurrence
Nowy Kościół (formerly Neukirch) and Leszczyna (formerly Haasel; 6 km NE of Nowy Kościół), Poland, Lower Carbonate cycle (Ca1) of Zechstein Group, Wuchiapingian stage, Lopingian, Permian.
Remarks
There are two specimens of Langenhan-collected fossils present in the University of Wrocław collections (MGUWr-1780s/1, 1780s/2) and 30 newly collected specimens from the type locality (NPL-0001–0018). The lectotype, which has an anterior-posterior length of 6.4 cm and dorsal-ventral width of 2.1 cm, has good preservation of growth lines and shows a blunt anterior apex. The newly collected specimens confirm the shell shape and pattern of growth lines illustrated by Langenhan (Reference Langenhan1908). The new specimens supplement the Langenhan (Reference Langenhan1908, p. 123) description by revealing the presence of a subdorsal shallow groove below the hinge line and showing that the supposed radial lines of juvenile shells is merely a group of raised bundles of growth lines present on the anteroventral margin of the shell. The new specimens include one with the prodissoconch preserved and a few specimens with a well-defined blunt anterior margin. Several specimens show a change in angle of intersection of the posterior growth line with the hinge progressing from an acute angle on the juvenile shell to an obtuse angle on the mature shell. The prodissoconch and early nepioconch have a rounded posterior margin that changes to an acute, projecting margin within a few millimeters.
Aviculopinna neukirchensis can be distinguished from A. pinnaeformis by its smaller, more shallow subdorsal radial groove, by its smaller size, lesser inflation, and change from acute angle to obtuse angle intersection of growth lines with the dorsal margin (hingeline) within 2 cm of the anterior end of the shell.
Family Pterineidae Miller, Reference Miller1877
Genus Pteronites M'Coy, Reference M'Coy1844
Figure 10
Type species
Pteronites angustatus MʽCoy, Reference M'Coy1844, by subsequent designation of Stoliczka (Reference Stoliczka1871, p. 388; printing error Pterinites [sic] angustus [sic]).

Figure 10. Pteronites angustatus, holotype (NMING-F7369). Scale bar is 1 cm.
Diagnosis
Small shells (<5 cm long) with small subterminal beak projecting slightly above (dorsal to) hinge line; straight opisthodetic hinge line; angular posterior wing without sulcus on posterior margin, meeting the posterior part of the hinge line with acute angle.
Remarks
The holotype (NMING F7369) of Pteronites angustatus M'Coy, Reference M'Coy1844, type species for Pteronites, is preserved as a mold in a muddy sand matrix. The type is 4 cm long and 1.6 cm wide with shell inflation of 2–3 mm. There is a small beak projecting 0.5 mm above (dorsal to) the hinge line close to the anterior end. Syntype NMING-F5193 has a small, rounded anterior wing. Growth lines meet the hinge line margin at a high angle at a midpoint then become inclined to an acute angle of ~30° on the posterior portion of the shell. More complete description requires having better preserved specimens available for study.
The presence of a subterminal beak raised above the hinge line as a raised umbo, angular posterior, protruding wing, and small size are characters indicative of placement in the Pterineidae. The type species of Pteronites, P. angustatus M'Coy, Reference M'Coy1844, lacks most characters common to pinnids and the genus Pteronites is excluded from the Pinnidae. The presence of a dorsomarginal fold on the hinge line is a character shared with some pterineid genera (e.g., Leptodesma and Leiopteria). Pteronites has a size and shell form suggesting an epifaunal life style.
Results
The Gera and Nowy Kościół specimens provide a foundation for redescription of the genus and stabilization of the type species of Aviculopinna. Specimens preserving the anterior (apex), the juvenile, and the mature portions of the shell are documented from Zechstein species, providing data needed to establish a good genus description. Newly collected material includes specimens preserving anterior shell apex with growth lines present on prodissoconch and nepioconch portions of the shell (Fig. 5.2) and specimens (Fig. 5.3) showing the position of the prodissoconch relative to the rounded anterior apex of the shell. These specimens display pinnid bivalve characters, including a dorsomarginal fold that holds the ligament, a thick outer columnar prismatic shell layer, and an equivalved shell. They provide evidence for confident assignment of the genus to family Pinnidae, resolving uncertainty introduced by the proposed synonymy of Aviculopinna with Pteronites.
Specimens of Aviculopinna have a spatulate shape with a rounded anterior apex, a blunt posterior margin, and are thin-shelled (Fig. 5). Maximum observed length (anterior-posterior dimension) is 12 cm. The shell has a single shallow radial groove located ~20%–25% of the dorsal-ventral distance below the dorsal margin—a character not previously indicated in species descriptions, but present in Zechstein specimens and distinct to the genus. This feature needs low-angle light illumination to be seen properly, but is confirmed to be present on nearly all of the specimens examined. These characters are distinctive and distinguish Zechstein pinnids from other late Paleozoic pinnids.
Important genus characters of Aviculopinna include: (1) elongate trapezoid shape with apical angle of 10–15° on young shells and decreasing on mature shells; (2) growth lines that are linear on the ventral margin, then curve up on the posteroventral corner to become straight along the posterior margin; (3) growth lines meet the hinge line at a vertical to obtuse angle on adult shell; (4) presence of a well-developed dorsomarginal fold (holds ligament); (5) presence of a shallow subdorsal groove; (6) presence of a blunt apex; (7) absence or minimal development of a byssal flange; and (8) prodissoconch located on the hinge line ~2 mm from the anterior end of the shell. In contrast, drawings presented by Geinitz (Reference Geinitz1861) show a raised umbo like that of Bakevellia. Specimens of young Aviculopinna do not have a raised, vaulted umbo. Shell containing the prodissoconch has the same smooth inflated surface as the surrounding shell.
Shell dimensions are available for several specimens (Table 1). The shell widens with rapid increase in width within 0.5 cm of the apex, then has a 10–15° apical angle for most of the mature shell. Some longer specimens have nearly parallel dorsal and ventral margins. Shell inflation is low (I/W = 0.20–0.33 for A. neukirchensis) and increases along the shell length between apex and mid length. On specimens with the posterior end preserved, posterior shell inflation decreases, a feature expected for an animal that flexes the shell for closure, which is a standard action of pinnids (Yonge, Reference Yonge1953; Turner and Rosewater, Reference Turner and Rosewater1958; Waller, Reference Waller and Morton1990). Specimens of A. pinnaeformis are not suitable for such measurements but appear to have a similar pattern of shell inflation.
Table 1. Dimensions and calculated inflation index for three specimens of A. neukirchensis, lower carbonate cycle (Ca1), Zechstein Group, late Wuchiapingian Stage, Lopingian, Permian, Nowy Kościół (Silesia), Poland. Measurements of the three specimens are arranged in a series corresponding to distance from apex of the shell. L = shell length, W = dorsal-ventral width, I = inflation of combined valves.

There are many specimens of Aviculopinna present at Nowy Kościół, some of which are preserved in growth position (Fig. 6) in calcareous sandy mudstones and carbonates (Raczyński, Reference Raczyński1997; Biernacka et al., Reference Biernacka, Borysiuk and Racyński2005) in a position nearly vertical to bedding, which is a standard pinnid orientation of anterior-posterior axis vertical relative to the sediment surface. Shells preserved in life position in calcareous mudstone often show anterior-posterior wrinkling and folding of the shell resulting from sediment compaction. Despite the compaction, they preserve original dimensions of shell width and shell inflation. Preservation of the carbonate shell occurs in some shell-concentration layers produced by waves and storm-generated bottom currents. The presence of common pinnids in strata subject to storm wave action (Raczyński, Reference Raczyński1997; Biernacka et al., Reference Biernacka, Borysiuk and Racyński2005) indicates life within a shallow marine, inner shelf depositional setting. The concentration of shells shown on a bedding plane (Fig. 7) indicates many specimens living close together, a common feature of living pinnids and fossil pinnid occurrences.
Although some specimens in the Museum für Naturkunde Gera and Langenhan specimens in the University of Wrocław collections are labeled as being from middle Zechstein strata, the Gera specimens are considered to be from the carbonate member of the Werra Formation (z1), the lowest Zechstein cycle in Germany, and the Nowy Kościół specimens are from the co-eval lower carbonate-bearing depositional cycle (Ca1) of the Zechstein in Poland (Piątkowski, Reference Piątkowski1966; Paul, Reference Paul, Harwood and Smith1986; Biernacka et al., Reference Biernacka, Borysiuk and Racyński2005; Peryt et al., Reference Peryt, Geluk, Mathiesen, Paul, Smith, Doornenbal and Stevenson2010a).
Discussion
Examination and description of specimens of the two described Zechstein species of Aviculopinna, from Gera, Germany and Nowy Kościół, Poland, provide much more data on the characters of the genus and its type species A. pinnaeformis. Photographs of specimens previously illustrated by drawings (Geinitz, Reference Geinitz1861; Langenhan, Reference Langenhan1899 [1900], Reference Langenhan1908) reveal distinctive characters for the genus. Major features of Aviculopinna include having an elongate, spatulate shape with a nearly straight posterior margin, growth lines that meet the hinge line at an obtuse angle, and an anterior end that is blunt and rounded and extends up to 2 mm anterior of the prodissoconch. Shell width (dorsal-ventral dimension) increases rapidly during juvenile growth and increases slowly after reaching a 0.5 cm shell length (anterior-posterior dimension). Dorsal and ventral margins on young and mature shells are linear, and those portions of the shell have an apical angle of ~10–15°. The basal Zechstein species A. pinnaeformis and A. neukirchensis have a shallow, radial, subdorsal groove near the hinge line. This combination of characters distinguishes the genus Aviculopinna from other late Paleozoic pinnid genera. The redefined description of Aviculopinna shows that most pinnid species currently assigned to the genus do not belong in the genus. A neotype is selected for the Aviculopinna type species A. pinnaeformis (MNG–8528 and counterpart MNG–8527) to stabilize the species (Fig. 4.1) and genus circumscriptions, and a lectotype (MGUWr-1780s/1) is selected for A. neukirchensis (Fig. 5.1).
Aviculopinna is most similar to genus Meekopinna Yancey, Reference Yancey1978, another genus with a straight, linear posterior shell margin. The two genera differ in that Meekopinna has uplifted shell lamellae on growth lines whereas shell lamellae are lacking in Aviculopinna. The growth lines with uplifted shell lamellae of Meekopinna are more regularly spaced than the growth line bundles of Aviculopinna. Both Aviculopinna and Meekopinna have some shell growth anterior of the prodissoconch (2 mm for Aviculopinna and 1–2 mm for Meekopinna) and a blunt apex. Other late Paleozoic pinnid fossils have a much more angular anterior apex, but available specimens lack preservation of the prodissoconch to make reliable comparisons possible.
The mention of anterior and posterior wings on the shell of Aviculopinna (Geinitz, Reference Geinitz1861) is incorrect. The blunt anterior end is evenly rounded and extends no more than 2 mm anterior of the prodissoconch. There is no posterior wing on a shell where the posteroventral corner of the shell projects farther than the hinge line (dorsoventral corner). The written mention of striae crossed with growth lines is confusing because the only radial line crossing growth lines is the single shallow groove near the hinge line. A slight gap for the byssal opening is normal for pinnids but it is seldom developed enough to be seen as a distinct character and is not observed on the shells examined.
The interpretation that Aviculopinna has radial lines on the shell originates with Münster (Reference Münster1839) and was exaggerated by Langenhan (Reference Langenhan1908, pl. 1, figs. 6–8) when presenting illustrations interpreted to show shells with strong radial ridges. On a Langenhan specimen (Fig. 8) present in the Wrocław University collections (MGUWr-1780s/2), the apparent radial lines are identifiable as ridges of grouped, raised growth lines present on the anteroventral margin of shells. They are growth margins, not radial lines crossing growth lines. The raised ridges appear on the anteroventral part of the shell but shrink and smooth out toward the posterior. Raised, thickened growth ridges have semi-regular spacing and are separated by intervals of finer growth lines. Growth lines and ridges (Fig. 8) can be traced as they curve upward to meet the dorsal hinge line and a similar semi-regular grouping of growth lines may be visible along portions of the hinge line. They appear to mark periodic growth interruptions corresponding with regular (perhaps monthly) tidal variations. The raised growth ridges on shell fragments were illustrated by Langenhan (Reference Langenhan1908, pl. 1, figs. 6–8) without discussion of how they fit with the rest of the shell. Specimen MGUWr-1780s/2 cannot be identified as one of the three drawings in Langenhan (Reference Langenhan1908), but it is certainly the basis for one or more of the drawings. This specimen shows that the raised growth line ridges are part of the normal growth lines that occur on the anteroventral region of mature shells. Langenhan's (Reference Langenhan1908, pl. 1, figs. 6–8) figures are incorrect representations of growth line ridges and should be dismissed as improper reconstructions.
Better descriptions of Zechstein species of Aviculopinna make it possible to test the validity of the proposed synonymy of Aviculopinna with Pteronites M'Coy, Reference M'Coy1844, by Cox and Hertlein (Reference Cox, Hertlein and Moore1969, p. N283). That action resulted in the placement of all smooth-shelled Paleozoic pinnids into a single poorly defined genus and raised the possibility that fossils recognized as late Paleozoic pinnids have shell characters unlike those of living pinnids. If accepted, the synonymy suggests a placement of those late Paleozoic species in the Pterineidae, not the Pinnidae. This was not a useful action because pinnids proliferated in the late Carboniferous into large and small species of varied morphology. At the same time, Sulcatopinna Hyatt, Reference Hyatt1892, was synonymized with Pinna (Cox and Hertlein, Reference Cox, Hertlein and Moore1969, p. N283), but this is unjustified and the Sulcatopinna name continues to be used for Paleozoic species while the name Pinna is abandoned for pinnids of that age. Similarly, the synonymy of Aviculopinna (based on a late Permian species) with Pteronites (an early Carboniferous genus) is not accepted by most workers despite the previous lack of documentation of the dissimilarity. A redescription of genus Pteronites is given here for comparison with genus Aviculopinna, which shows the two genera differ in many ways to an extent that they are assigned to different families.
Paleozoic pinnid relationships based on terminal versus subterminal position of the prodissoconch
The proposed synonymy of Aviculopinna with Pteronites by Hind (Reference Hind1901) and Cox and Hertlein (Reference Cox, Hertlein and Moore1969) created the hypothesis that differences in prodissoconch position on pinnid species was an important factor in determining pinnid relationships and phylogeny. This is problematic because all living pinnids have entirely terminal beaks (Turner and Rosewater, Reference Turner and Rosewater1958; Schultz and Huber, Reference Schultz and Huber2013), so the occurrence of subterminal beaks like that of Pteronites (the proposed senior synonym of Aviculopinna) implied the existence of a separate group of pinnids or a group separated from pinnids. Living and fossil pinnids have a very distinctive life habit characterized with having a vertical anterior-posterior shell orientation, living in a fixed location, and held in place by byssal attachment. This normally results in shell damage to the apex by abrasion and corrosion (Turner and Rosewater, Reference Turner and Rosewater1958), so continued shell growth anterior of the prodissoconch serves no useful function.
Although there is a difference in position of the prodissoconch on species of Meekopinna and Aviculopinna from pinnid species with a completely terminal beak, the difference is minor. There is no development of a raised umbo or slanting umbonal ridge. Another reason to consider the terminal versus subterminal beak position concept as unimportant in determining pinnid relationships is the lack of documentation of prodissoconch position on late Paleozoic pinnids other than species of Aviculopinna and Meekopinna. Other late Paleozoic pinnid species, including species of Sulcatopinna and smooth-shelled genera, are simply assumed to have a terminal beak; it is not a documented character. Without data to test it, this leaves the concept of determining relationship based on terminal versus subterminal beak position without a basis for evaluating its validity. At this time, it remains a hypothesis and without utility in the study of fossils pinnids.
Some studies of early Mesozoic pinnids (Waller and Stanley, Reference Waller and Stanley2005; McRoberts, Reference McRoberts2017) have considered prodissoconch (beak) position as a factor in describing post-Paleozoic pinnids, along with the grouping of pinnid into clusters having divided or non-divided nacreous inner shell layer. Studies of middle and late Mesozoic pinnids (Schultz and Huber, Reference Schultz and Huber2013; Koppka, Reference Koppka2018) are concerned with grouping based on divided versus non-divided nacreous shell layer and make no reference to beak position as an important character. The apex of other late Paleozoic and Triassic pinnid genera remains unknown, so the importance of terminal versus subterminal beak position cannot be tested or evaluated. It stands as a concept with a weak foundation, based primarily on assumptions instead of data.
Conclusions
The synonymy of Aviculopinna with Pteronites proposed by Cox and Hertlein (Reference Cox, Hertlein and Moore1969) is inappropriate because species of Pteronites are much smaller, have a wing-like posterior termination, and have a more rounded shape than Aviculopinna species; they are limited to early Carboniferous strata. Aviculopinna is established for a late Permian species of larger size with a slender trapezoid shape. Aviculopinna has a rounded, blunt apex with 2 mm of anterior growth but no inflated umbo or beak protruding dorsally above the hinge line. The age of occurrence of Aviculopinna is limited to the late Permian, excluding many species presently identified as being members of the genus. A neotype is designated for the genotype species Aviculopinna pinnaeformis Geinitz from Gera, Germany, and the genus characters are supplemented by comparison to well-preserved specimens of the closely related Aviculopinna neukirchensis Langenhan from Nowy Kościół, Poland. An expanded description of A. neukirchensis is presented based on new collections of specimens and a lectotype is selected for A. neukirchensis from specimens collected by A. Langenhan.
A redescription of the type species Pteronites angustatus M'Coy is presented to provide comparison to Aviculopinna. An evaluation of the subterminal beak versus terminal beak concept in pinnids is presented with the conclusion that there are few data available to support the concept or test it. Extremely few fossil pinnids are preserved with the apex present or in a condition to identify the prodissoconch position.
Acknowledgments
We are grateful for the help of S. Butts and J. Utrup, Yale Peabody Museum, USA, for providing data and loaning specimens of fossils identified and sent to the museum by H.B. Geinitz; to museum curators in Germany and Poland who responded to our queries when searching for specimens studied by H.B. Geinitz and G. Graf zu Münster; and to B. Seuss, Erlangen and W. Werner, Munich, Germany, for help in contacting museum staff. Special thanks to F. Hrouda, Museum für Naturkunde Gera, for assistance during a visit of the Geinitz collection and H. Kaufmann, Burgwald, Germany, for additional information on unstudied Zechstein bivalves. Comments by reviewer M.G. Simoes and an anonymous reviewer helped improve the manuscript for publication.
Author contributions
This study originated from work on taxonomic revision of North American late Paleozoic pinnids by TY. MA visited and examined fossils kept in the Museum für Naturkunde Gera, Germany, evaluated data on stratigraphic occurrences, and provided critical editing of the text. PR provided critical new documentation of well-preserved fossils, collected new material, and provided photographs of the Langenhan specimens retained in the Uniwersytet Wrocławski collections, Poland. SB compiled data on the stratigraphic occurrence of the genus from literature and field experience.
Competing interests
The authors declare no competing interests.












 Open access
Open access