Introduction
Zealandia is a center of extant penguin diversity, and accumulating fossil evidence supports the hypotheses that both total-group and crown penguins originated in this region (Thomas et al., Reference Thomas, Tennyson, Scofield, Heath, Pett and Ksepka2020b). The oldest and most stemward-known fossil penguin, Waimanu manneringi Ando, Jones, and Fordyce in Slack et al., Reference Slack, Jones, Ando, Harrison, Fordyce, Arnason and Penny2006, was uncovered in the early Paleocene (ca. 61.6–60.5 Ma) Waipara Greensand of the South Island of New Zealand (Slack et al., Reference Slack, Jones, Ando, Harrison, Fordyce, Arnason and Penny2006). Since that discovery, a number of additional stem group penguins have been reported from the Waipara Greensand, capturing the morphology of penguins during the early stages of the evolution of flightless wing-propelled diving (Mayr et al., Reference Mayr, Scofield, De Pietri and Tennyson2017a, Reference Mayr, De Pietri, Love, Mannering and Scofieldb, Reference Mayr, De Pietri, Love, Mannering and Scofield2019). Contemporary penguin fossils from the Paleocene Takatika Grit (62.5–60 Ma) have been reported from the Chatham Islands, an archipelago located ~750 km from mainland New Zealand (Blokland et al., Reference Blokland, Reid, Worthy, Tennyson, Clarke and Scofield2019). Slightly younger penguins from the late Paleocene Moeraki Formation (59.5–55.9 Ma) occur on mainland New Zealand. Two Moeraki Formation penguin specimens have been published to date: a partial coracoid and scapula of a modest-sized penguin mentioned briefly in a review (Fordyce and Jones, Reference Fordyce, Jones, Davis and Darby1990) and a partial skeleton of the giant stem penguin Kumimanu biceae Mayr et al., Reference Mayr, Scofield, De Pietri and Tennyson2017a. A crushed distal end of a tibiotarsus (OU8743) from the Moeraki Formation, which was identified as belonging to a large bird by Fordyce (Reference Fordyce, Vickers-Rich, Monaghan, Baird and Rich1991), also appears to belong to a penguin (A.J.D.T., personal observation, 2015).
Along with the aforementioned Paleocene fossils, younger specimens ranging in age from the early Eocene Kaiika maxwelli Fordyce and Thomas, Reference Fordyce and Thomas2011 (Kauru Formation), to the recently extinct Eudyptes warhami Cole et al., Reference Cole, Ksepka, Mitchell, Tennyson and Thomas2019, document a remarkable diversity of 23 named penguin species from the fossil record of New Zealand, with additional taxa either awaiting formal description or inferred to be present based on fragmentary material (Huxley, Reference Huxley1859; Oliver, Reference Oliver1930; Marples, Reference Marples1952, Reference Marples1960; Simpson, Reference Simpson1971, Reference Simpson1972; Fordyce and Thomas, Reference Fordyce and Thomas2011; Ksepka et al., Reference Ksepka, Fordyce, Ando and Jones2012; Thomas and Ksepka, Reference Thomas and Ksepka2016; Mayr et al., Reference Mayr, Scofield, De Pietri and Tennyson2017a, Reference Mayr, De Pietri, Love, Mannering and Scofieldb; Cole et al., Reference Cole, Ksepka, Mitchell, Tennyson and Thomas2019; Thomas et al., Reference Thomas, Ksepka, Holvast, Tennyson and Scofield2020a, Reference Thomas, Tennyson, Scofield, Heath, Pett and Ksepkab; Giovanardi et al., Reference Giovanardi, Ksepka. and Thomas2021).
One surprising insight into the evolutionary history of penguins revealed by recent discoveries is the apparently rapid attainment of enormous body size in the Paleocene, as evinced by specimens with proportions greatly outstripping those of the largest extant penguins (Tambussi et al., Reference Tambussi, Reguero, Marenssi and Santillana2005; Mayr et al., Reference Mayr, De Pietri, Love, Mannering and Scofield2017b). Given that the earliest giant New Zealand penguin fossils pre-date the occurrence of penguins on other continents, it is tempting to speculate that penguins approached the peak of their known body size distribution while still restricted to their Zealandian center of origin. However, this hypothesis remains largely untested due to gaps in the early Paleocene marine record elsewhere in the Southern Hemisphere, and uncertainties regarding the phylogenetic relationships among early stem penguins. Therefore, untangling the phylogenetic relationships of Paleocene stem penguins has potential to clarify key aspects of penguin macroevolutionary change. These include revealing the tempo and pattern of sphenisciform body size evolution and providing new insight into the evolution of the characteristic penguin bauplan, including their distinctive flipper apparatus.
In this study, we report nine new stem penguin specimens from the late Paleocene Moeraki Formation. All were collected from beach-washed boulders at Hampden Beach (Otago, New Zealand) (Fig. 1). At this locality, concretions occur in situ in mudstones but are more often found completely exhumed from the softer surrounding matrix by coastal erosion (A.J.D.T., personal observation, 2021). The new penguin specimens were all found within the intertidal zone in remnants of concretions that had been shattered by the elements to reveal their interiors. Given the state of wear and rounding of many of these rocks, most evidently had been subject to wave action for many years. Although this leaves their precise stratigraphic horizon uncertain, the contact between the Moeraki Formation and the overlying Kurinui Formation is considered to be equivalent to the Teurian/Waipawan boundary (and hence the Paleocene/Eocene boundary) (Morgans, Reference Morgans2009), providing a minimum age of ca. 56.0 Ma for the fossils. The upper part of the Moeraki Formation exposed at Hampden Beach is specifically considered to represent the upper Teurian local stage based on evidence from microfossils (Crouch and Brinkhuis, Reference Crouch and Brinkhuis2005; Morgans, Reference Morgans2009). Evidence from agglutinated foraminiferal taxa suggests that the fossiliferous concretions originally formed in a relatively shallow shelf to upper bathyal environment (Morgans, Reference Morgans2009).
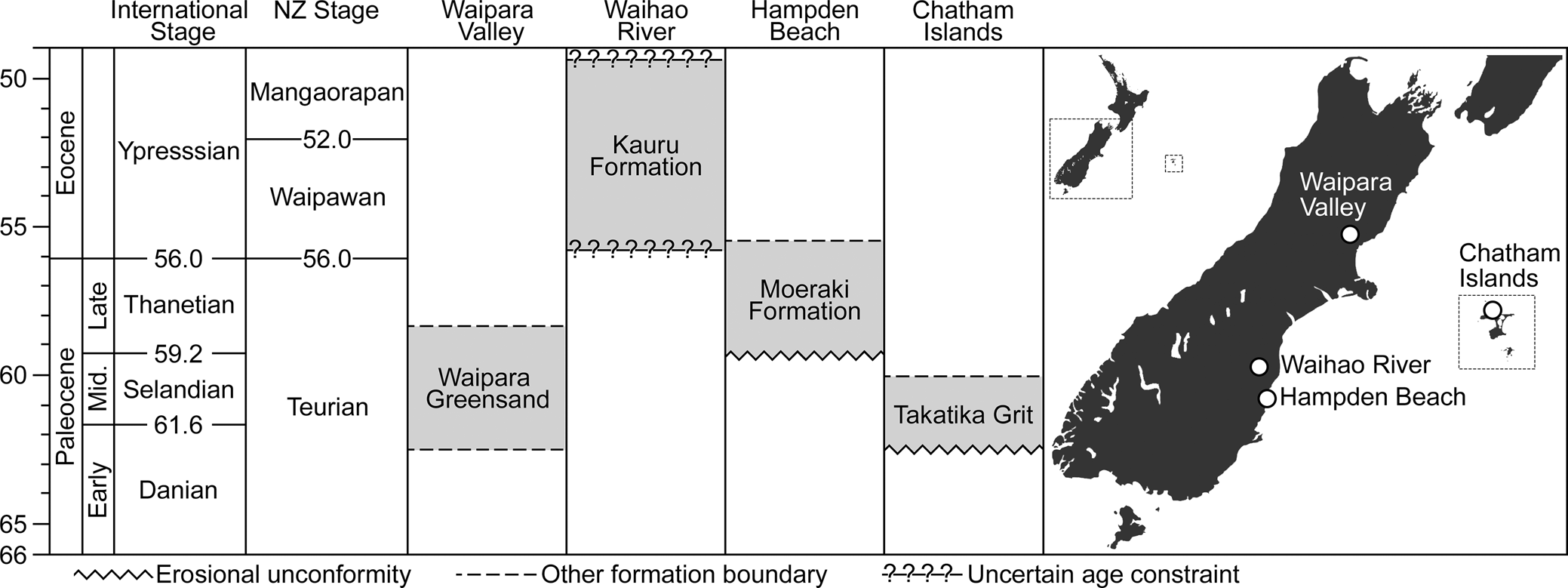
Figure 1. Simplified comparison of stratigraphic horizons for key Paleogene penguin localities in New Zealand. Fossil sites discussed in this paper are indicated on map at right.
The largest of the new fossils belongs to a new species of giant penguin represented by a partial associated skeleton, including the largest penguin humerus yet reported. An additional large fossil can be referred to cf. Kumimanu biceae, and five smaller specimens belong to a penguin slightly larger than the extant emperor penguin, Aptenodytes forsteri Gray, Reference Gray1844, that we describe as a second new species. The two smallest specimens belong to an unnamed penguin intermediate in size between the king penguin Aptenodytes patagonicus Miller, Reference Miller1778, and the emperor penguin. We provide allometric regressions based on the dimensions of humeri from crown penguins to estimate body mass in the new fossil taxa and present an expanded phylogeny of extinct and extant total clade penguins.
Materials and methods
Anatomical nomenclature
In the descriptions below, we use the anatomical terminology of Baumel and Witmer (Reference Baumel, Witmer, Baumel, King, Breazile, Evans and Vanden Berge1993). Following the recent suggestion of Richards (Reference Richards2019), who considered that the vertical posture of penguins results in the terms “cranial” and “caudal” being applied to different faces of the flipper than when applied to the wings of a typical bird, we orient the humerus in an “extended during swimming” position for purposes of the description (e.g., the oblong fossa marking the insertion of m. pectoralis is placed on the ventral face of the bone in this context).
3D digital replicas
Digital replicas of fossils (i.e., 3D meshes) were generated using a HandySCAN BLACK handheld laser scanner (Creaform Inc., Quebec, Canada). 3D scan data were processed using VXelements 9.1.0 (Creaform Inc., Quebec, Canada). To facilitate systematic comparisons, humeri were digitally extracted from their respective fossil blocks and made into watertight meshes using VXelements 9.1.0. 3D meshes were rendered using Blender 2.93 (Blender Online Community, 2022) and were digitally mirrored when necessary to facilitate comparisons between left versus right elements. The minimum resolvable surface feature of the 3D models was ~0.2 mm diameter.
The humerus of NMNZ S.47426 has been twisted out of life position by deformation. In order to correct this distortion, we used Blender 2.93 to digitally separate the head from the diaphysis, and then to separate the head into two sections, with the separation following the broken edges between each part. We then manually reoriented these three parts using digital models of other fossil penguin humeri as visual guides.
Phylogenetic analyses
We scored the new species into a data set based on the extant penguin matrix of Bertelli and Giannini (Reference Bertelli and Giannini2005), which was expanded to include fossil taxa by Ksepka et al. (Reference Ksepka, Bertelli and Giannini2006) and has been modified and expanded in numerous subsequent studies, most recently by Giovanardi et al. (Reference Giovanardi, Ksepka. and Thomas2021). We added the previously described fossil penguins Crossvallia unienwillia Tambussi et al., Reference Tambussi, Reguero, Marenssi and Santillana2005, Crossvallia waiparensis Mayr et al., Reference Mayr, De Pietri, Love, Mannering and Scofield2019, Kaiika maxwelli, and Kupoupou stilwelli Blokland et al., Reference Blokland, Reid, Worthy, Tennyson, Clarke and Scofield2019, and added or modified seven characters to represent new features observed in our review of recently collected Paleocene penguin material. The expanded matrix comprises 279 morphological characters and 2 outgroups, 20 extant penguin taxa, and 44 fossil penguin taxa.
Phylogenetic analyses were conducted using PAUP*4.0a168 (Swofford, Reference Swofford2003) with a heuristic search strategy specifying 10,000 replicates of random taxon addition holding 10 trees per replicate, with TBR branch swapping limited to 10,000,000 rearrangements per replicate. All characters were equally weighted, multistate codings were considered to represent polymorphism, and branches with a minimum length of zero were collapsed.
Bayesian phylogenetic analysis was conducted using RevBayes 1.1.1 (Höhna et al., Reference Höhna, Landis, Heath, Boussau, Lartillot, Moore, Huelsenbeck and Ronquist2016) using a specimen-level fossilized birth-death process as the prior on the tree topology and divergence times (Stadler, Reference Stadler2010; Heath et al., Reference Heath, Huelsenbeck and Stadler2014), with exponential hyperpriors on the diversification parameters. These analyses follow the protocol found in the RevBayes tutorial (https://revbayes.github.io/tutorials/fbd/fbd_specimen.html). Two chains were run with 1 move per generation until the maximum difference of clade frequencies (max_diff, Lartillot and Philippe, Reference Lartillot and Philippe2004) was <0.1 (350,000,000 generations). All data and scripts associated with these analyses are provided in the supplementary information.
Repositories and institutional abbreviations
AMNH, Department of Ornithology, American Museum of Natural History, New York, U.S.A.; CM, Canterbury Museum, Christchurch, New Zealand; IB/P/B, Institute of Biology, University of Białystok, Białystok, Poland; MEF, Museo Egidio Feruglio, Trelew, Argentina; MLP, Museo de La Plata, La Plata, Argentina; NMNZ, Museum of New Zealand Te Papa Tongarewa, Wellington, New Zealand; MUSM, Museum of San Marcos University, Lima, Peru; MNHN, Muséum National d'Histoire Naturelle, Paris, France; OM, Otago Museum, Dunedin, New Zealand; OU, Geology Museum, University of Otago, Dunedin, New Zealand; SAM, Iziko South African Museum, Cape Town, South Africa; SGO-PV, Sección de Paleontología del Museo Nacional de Historia Natural, Santiago, Chile; UCMP, University of California Museum of Paleontology, Berkeley, California, U.S.A.; UCN, Museo Geológico Prof. Humerto Fuenzalida, Universidad Católica del Norte; WM, Waikato Museum, Hamilton, New Zealand.
Systematic paleontology
Aves Linnaeus, Reference Moore, Douglas, Mills, McKinley, Nelson and Murphy1758
Sphenisciformes Sharpe, Reference Sharpe1891
Kumimanu Mayr, Scofield, De Pietri, and Tennyson Reference Mayr, De Pietri, Love, Mannering and Scofield2017
Type species
Kumimanu biceae Mayr, Scofield, De Pietri, and Tennyson Reference Mayr, De Pietri, Love, Mannering and Scofield2017.
Revised diagnosis
Differs from all other Paleocene penguins in larger size and in robust femur shaft (observable only in Kumimanu biceae). Differs from Waimanu, Muriwaimanu Mayr et al., Reference Mayr, De Pietri, Love, Mannering and Scofield2017b, Sequiwaimanu Mayr et al., Reference Mayr, De Pietri, Love, Mannering and Scofield2017b, and Petradyptes n. gen. in lacking ovoid depressions on the lateral faces of the posterior thoracic vertebrae. Differs from Muriwaimanu, Sequiwaimanu, and Kupoupou in impressio m. pectoralis forming an extremely wide fossa. Differs from Waimanu, Muriwaimanu, Sequiwaimanu, and Crossvallia in greater distal extension of insertion scar for m. supracoracoideus (condition unknown in Waimanu) and placement of sulcus extensorius of tibiotarsus close to midline. Differs from Kaiika and Petradyptes in less strongly flattened humeral shaft. Further differs from all post-Paleocene penguins other than Kaiika in having the border of the sulcus scapulotricipitalis formed by a short posterior trochlear ridge, positioned on the ventral face of the humerus (versus a longer posterior trochlear ridge extending to the caudal border of the humeral shaft in Eocene and later penguins).
Kumimanu fordycei new species
Figure 2

Figure 2. Kumimanu fordycei n. sp. holotype (NMNZ S.47426). (1) Holotype block oriented to show humerus in dorsal view with humerus of an extant emperor penguin (Aptenodytes forsteri; NMNZ OR.23039) oriented in the same view to indicate scale, (2) block oriented with humerus in ventral view (partially overlain by scapula), (3) comparison of fifth cervical vertebra in caudal view with fifth cervical vertebra of emperor penguin for scale, (4) oblique view of humerus showing wide fossa formed by impressio m. pectoralis (which is partially hidden by overlying elements) in ventral view. (5) Distal portion of ulna shaft in dorsal view. (6) Close-up of tentatively identified patella. (7) Block oriented with distal end of humerus exposed and (8) close up of distal trochlea. Abbreviations: acr, processus acrocoracoideus; ch, caput humeri; cor, coracoid; cv, cervical vertebral; ft, fossa tricipitalis; hu, humerus; ip, impressio m. pectoralis; mtr, middle trochlear ridge; pat, patella; sb; scapular blade; sup, insertion scar for m. supracoracoideus; tc, tuberculum coracoideum, vtr, ventral trochlear ridge. Scale bars equal 10 mm.
Holotype
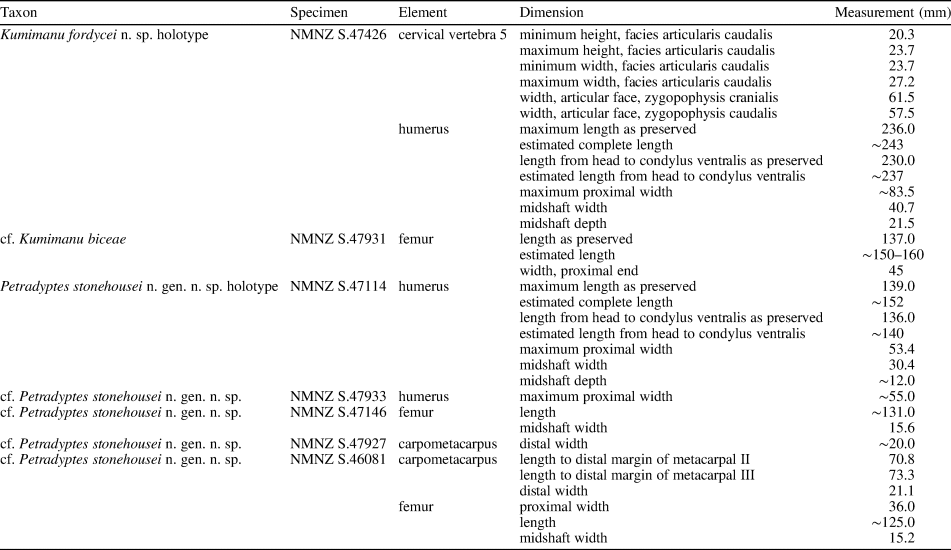
NMNZ S.47426: complete cervical vertebra, partial right coracoid, partial right scapula, nearly complete right humerus, shaft of right ulna, tentatively identified left and right patellae, fragment of left tibiotarsus, and several unidentified fragments of bone. Measurements in Table 1.
Table 1. Measurements from Moeraki Formation penguin fossils. All measurements in mm.
Diagnosis
Differs from Kumimanu biceae by substantially larger size (humerus midshaft width 40.7 mm versus 33.0 mm), more curved humerus shaft, and proportionally less expanded proximal end (proximal width ≈ 2.0 times midshaft width versus 2.27 times midshaft width in Kumimanu biceae).
Occurrence
The holotype was collected in a beach-washed concretion from the Moeraki Formation at Hampden Beach, North Otago, New Zealand (Fig. 1). This site is cataloged under NZ Fossil Record Number J42/f0956 (precise locality information is recorded at NMNZ).
Description
The holotype is preserved in a concretion (Fig. 2). The humerus, coracoid, and scapula are nearly in contact at the shoulder joint, whereas the remaining elements are disarticulated and scattered throughout the block. A single vertebra appears to correspond to cervical vertebra 5. It is heterocoelous and bears a short processus spinosus, a completely enclosed foramen transversarium, and a well-developed processus transversus. The articular facets of the zygapophysis cranialis and zygapophysis caudalis are extremely enlarged compared to those of extant penguins (Fig. 2.3). Enlarged facets on the vertebral articular processes are observed in other giant stem penguins such as Icadyptes salasi Clarke et al., Reference Clarke, Ksepka, Stucchi, Urbina, Giannini, Bertelli, Narváez and Boyd2007, whereas the proportions of these facets in smaller-bodied Paleocene penguins like Sequiwaimanu rosieae Mayr et al., Reference Mayr, De Pietri, Love, Mannering and Scofield2017b, resemble those of crown penguins (Ksepka et al., Reference Ksepka, Clarke, DeVries and Urbina2008).
The right scapula is preserved in two segments: a portion of the omal end and a larger portion of the blade. The tuberculum coracoideum is strongly rounded and omally projected. The ball-like morphology of the tuberculum coracoideum is very similar to that in Kumimanu biceae, although it is clearly larger in absolute size. In the smaller Paleocene stem penguins Muriwaimanu tuatahi (Ando, Jones, and Fordyce in Slack et al., Reference Slack, Jones, Ando, Harrison, Fordyce, Arnason and Penny2006) and Sequiwaimanu rosieae, the tuberculum coracoideum is also rounded, but is proportionally smaller and more weakly projected. A more oblong shape characterizes the tuberculum coracoideum in later stem penguins such as Icadyptes salasi, as well as extant penguins. Unfortunately, the acromion and facies articularis humeralis are not preserved. The intact portion of the scapular blade indicates it was flattened and widened but lacked the paddle-like distal expansion seen in Neogene penguins. Compared to Kumimanu biceae, the scapular blade of the new species appears less expanded, but this is possibly an artifact of preservation because the borders of the blade are slightly eroded.
The coracoid is nearly complete except for the omal end, but is generally poorly preserved. The length from the facies articularis sternalis to the omal rim of the cotyla scapularis is ~176 mm. If the proportions of the missing omal tip were similar to those in Sequiwaimanu rosieae, the coracoid would have been slightly shorter than the humerus, as in other Paleogene penguins. Such proportions would represent an intermediate condition between volant birds and later penguins: the coracoid is much shorter than the humerus in most extant birds, whereas the coracoid exceeds the humerus in length in all known Eocene and later penguins. The facies articularis sternalis is greatly widened and the presence of a processus lateralis cannot be determined.
The humerus is nearly complete, lacking only the distal condyles, although some morphological features are obscured by erosion or overlying bones and the proximal part of the caput humeri has been damaged. The shaft is strongly flattened and tapers distally. The impressio m. pectoralis forms an extremely wide fossa (Fig. 2.4), closely resembling the condition in Kaiika maxwelli and, to a lesser extent, some Alcidae. This oblong impression extends far distal to the level of the crista bicipitalis. The distal margins of the impressio m. pectoralis and crista bicipitalis appear to be at the same level in Kumimanu biceae (Fig. 3), however this may be at least in part an artifact of preparation given that the matrix has not been completely removed from the impression. The m. supracoracoideus insertion scar is elongated, raised, and shifted towards the caudal border of the shaft. The fossa tricipitalis is very deep and appears to have been undivided, although matrix obscures its exact dimensions. The ventral and middle trochlear ridges are intact, whereas the dorsal trochlear ridge has been lost to erosion (Fig. 2.8). The ventral and middle trochlear ridges are much more deeply separated than in crown penguins. The absence of the dorsal trochlear ridge indicates that it must have been anteriorly placed and failed to extend to the level of the other two trochleae, as in other Paleocene penguins.
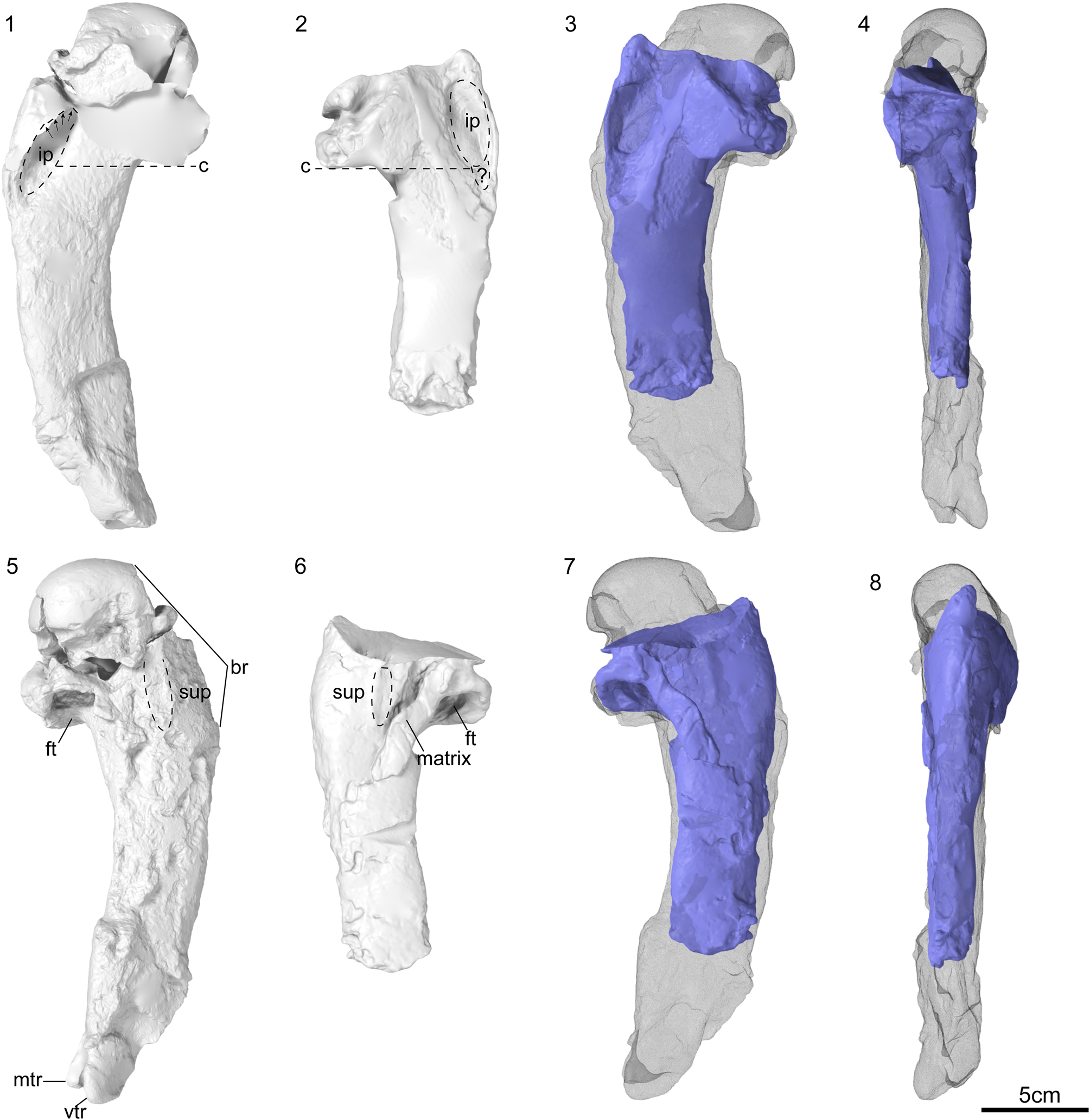
Figure 3. Comparison of the digitally reconstructed holotype humerus of Kumimanu biceae (NMNZ S.45877) and the holotype humerus of Kumimanu fordycei n. sp. (NMNZ S.47426). Images show watertight models after digital extraction from fossil blocks, rendered using Blender 2.93 (Blender Online Community, 2022). The two broken sections of the head were digitally re-aligned to one another and to the diaphysis for Kumimanu fordycei. Ventral view of (1) right humerus from K. fordycei n. sp. and (2) left humerus of K. biceae. (3, 4) Digitally mirrored K. biceae humerus (blue) overlain on K. fordycei humerus (wireframe) in (3) ventral and (4) caudal view. Dorsal view of (5) right humerus from K. fordycei and (6) left humerus of K. biceae. (7, 8) Digitally mirrored K. biceae humerus (blue) overlain on K. fordycei humerus (wireframe) in (7) dorsal and (8) cranial view. Abbreviations: br, broken margin; c, line indicating level of distal margin of crista bicipitalis; ft, fossa tricipitalis; ip, impressio m. pectoralis; matrix = matrix/bone fragment adhering to surface of humerus; mtr, middle trochlear ridge; sup, insertion scar for m. supracoracoideus; vtr, ventral trochlear ridge; ?, possible extension of impressio m. pectoralis obscured by matrix. Arrows indicate extension of impressio m. pectoralis, which is obscured by overlying matrix in NMNZ S.47426.
A portion of the right ulna including the distal third of the shaft is preserved, although the distal articular region is lost. The cross-section is ovoid, as in Kumimanu biceae, being less dorso-ventrally flattened than in post-Paleocene penguins.
Two small irregular elements are tentatively identified as patellae. Each is block-like and has one strongly rimmed concave face resembling the articular surface for the femur; the more complete of the two also preserves a second concave face corresponding to the articular surface for the tibiotarsus (Fig. 2.6). Provided our identification of these ossifications as patellae is correct, they are smaller relative to body size than those of extant penguins (the long axis is within the size range of the much smaller emperor penguin). Unlike extant penguins, these elements also lack a sulcus or canal for the tendon of m. ambiens.
A small fragment of the left tibiotarsus, comprising the rim of the condylus medialis, is preserved near the head of the humerus at the edge of the block and was separated during preparation. As is typical for penguins, the epicondylus medialis is strongly developed. Its distal rim appears to lack the pronounced notch that is characteristic of Procellariiformes (the extant sister group to penguins), but which is extremely weakly developed or absent in most penguins. The mediolateral width of the condylus medialis is proportionally narrower than in extant penguins.
Etymology
In honor of R. Ewan Fordyce, in recognition of his contributions to the field of paleontology including his discovery, collection, and study of penguin fossils in New Zealand and Antarctica, and his generous mentorship of many paleontologists, including authors DTK and DBT, throughout their careers.
Remarks
Kumimanu fordycei is a candidate for the largest known penguin, as discussed below.
cf. Kumimanu biceae
Figure 4
Holotype
NMNZ S.45877: associated partial skeleton including three vertebrae and partial synsacrum, partial left scapula and right coracoid, cranial section of sternum, partial left humerus and ulna, right femur, and partial right tibiotarsus.

Figure 4. cf. Kumimanu biceae (NMNZ S.47931) proximal end of right femur and shaft of left femur, proximal ends of left tibiotarsus and fibula. Abbreviations: cc, crista cnemialis cranialis; cl, crista cnemialis lateralis; fe, femur; fib, fibula, il, tentatively identified iliac blade fragment; rib, rib. Scale bar equals 10 mm.
Occurrence
Late Paleocene Moeraki Formation at Hampden Beach, North Otago, New Zealand.
Description
The neck of the right femur is very thick, such that there is little constriction between the head and the shaft. The shaft of the left femur is unfortunately partially lost, thus a midshaft diameter cannot be reliably obtained. Based on the intact portion, we estimate this bone was slightly shorter than the femur of the Kumimanu biceae holotype. The proximal ends of the tibiotarsus and fibula, which are not preserved in any other specimens of Kumimanu, are intact. The crista cnemialis lateralis is relatively thin and comes to a sharp point. In caudal view, the ridge running along the long axis of the shaft is more prominent than in extant penguins.
Materials
NMNZ S.47931: fragments of rib and ilium, proximal end of right femur and shaft of left femur, proximal ends of left tibiotarsus and fibula. Measurements in Table 1.
Remarks
The referred specimen also belongs to a giant penguin, but is too small to belong to Kumimanu fordycei. The proximal end of the right femur is 45 mm wide versus 52 mm in the holotype of Kumimanu biceae, suggesting it may belong to a smaller individual of that species. Intraspecific size variation in stem penguins remains poorly understood, but the referred leg bones provide additional support for recognition of two species of Kumimanu because the holotype specimen of K. fordycei represents an individual ~15% larger than that represented by the holotype specimen of K. biceae, which in turn represents an individual ~15% larger than NMNZ S.47931.
Petradyptes new genus
Type species
Petradyptes stonehousei n. gen. n. sp.
Diagnosis
As for the type species by monotypy.
Etymology
From the Greek petra for rock and dyptes for diver. In Ancient Greek, petra carries a connotation of rock cliffs by the sea, eliciting the fossil locality.
Remarks
Although not the most complete of the specimens, we selected NMNZ S.47114 as the holotype to ease comparisons with existing Paleocene penguin species, the majority of which are based on holotypes that include a humerus. Several additional specimens are also assigned to Petradyptes. NMNZ S.47933 also preserves the proximal end of the humerus, which agrees well in size and morphology with the holotype and preserves the proximal end of the tibiotarsus, which in turn agrees well with the proximal tibiotarsus of NMNZ S.46081. NMNZ S.46081 preserves the femur and carpometacarpus, allowing comparisons to NMNZ S.47146 and NMNZ S.47927. This chain of evidence and the consistent overall size of the specimens support the conclusion that all belong to Petradyptes, although we consider these referrals provisional pending discovery of more complete specimens preserving the humerus.
Petradyptes stonehousei new species
Figures 5, 6
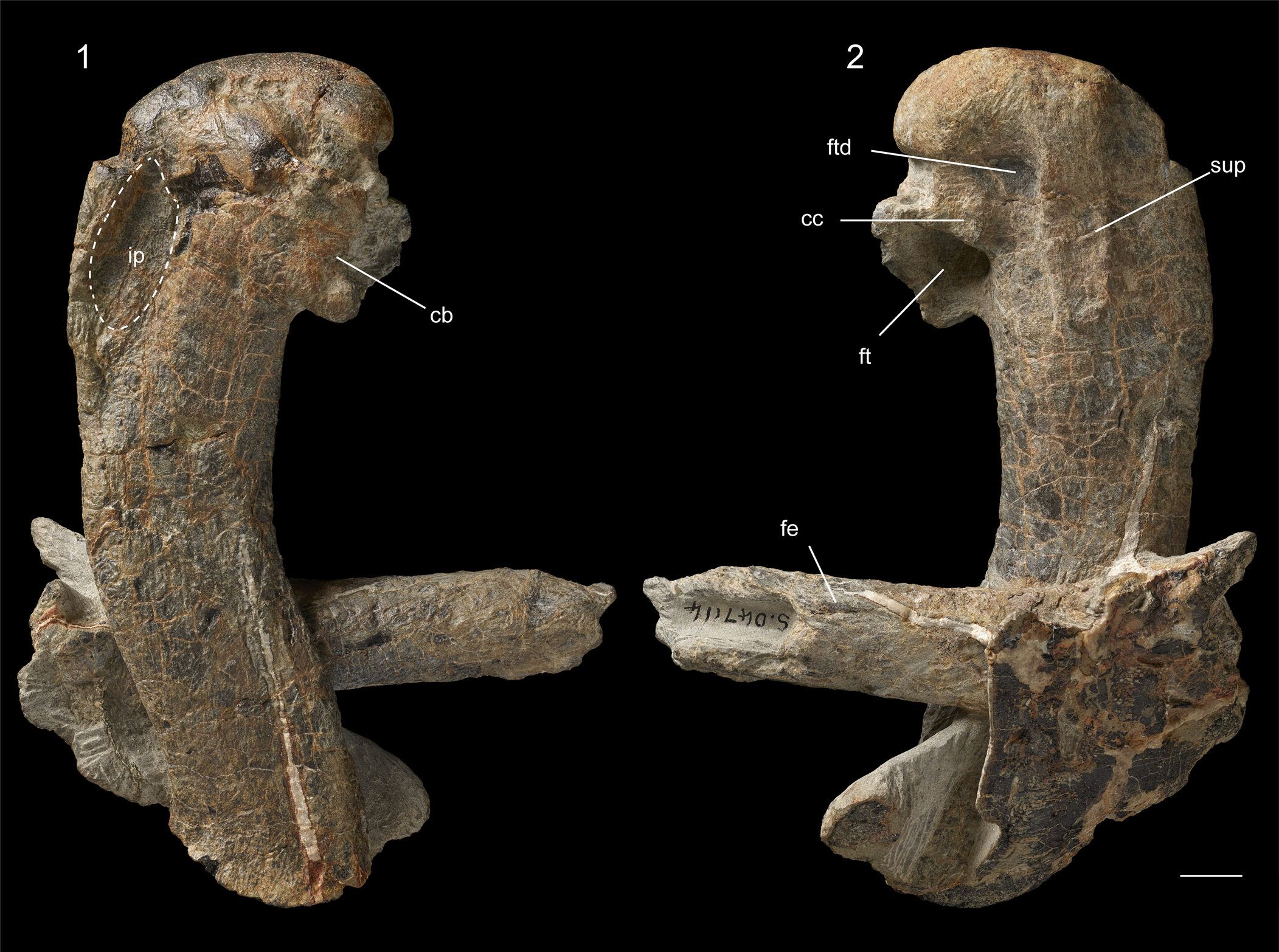
Figure 5. Petradyptes stonehousei n. gen. n. sp. holotype (NMNZ S.47114) complete right humerus and distal portion of left femur in (1) ventral view and (2) dorsal view. Abbreviations: cb, insertion scar for m. coracobrachialis caudalis; cc, insertion scar for m. coracobrachialis caudalis; fe, femur shaft; ft, fossa tricipitalis; ip, impressio m. pectoralis; ftd, fossa tricipitalis dorsalis; sup, insertion scar for m. supracoracoideus. Scale bar equals 10 mm.

Figure 6. Referred specimens of Petradyptes stonehousei. (1-5) NMNZ S.46081, oriented (1) exposing carpometacarpus in dorsal view with (2) close up of proximal portion of carpometacarpus, (3) exposing proximal end of tibiotarsus in cranial view with (4) close up of distal end of carpometacarpus, and (5) exposing femur in caudal view. (6-7) NMNZ S.47146, oriented (6) exposing femur in cranial view and pelvis in dorsal view and (7) exposing femur in caudal view and synsacrum in ventral view. (8-10) NMNZ S. 47933, oriented (8) with humerus exposed in oblique view showing dorsal face of proximal end and cross-section of broken shaft with (9) close-up showing wide fossa pectoralis, and (10) exposing proximal end of humerus in dorsal view. (11-13) NMNZ S. 47927 oriented (11) exposing omal end of scapula and distal end of tibiotarsus and (12) exposing broken carpometacarpus with (13) close up of distal end of carpometacarpus with exposed cross-section near midshaft. Abbreviations: acr, acromion; ccc, crista cnemialis cranialis; ccl, crista cnemialis lateralis; cmc, carpometacarpus; cv, cervical vertebra; ex, expansion at distal end of metacarpal II; fe: femur; fib, fibula; fo, fossa; fp, fossa pectoralis; hu, humerus; il, iliac blade; ip, impressio m. pectoralis; mc, medullary cavity; mcI, alular metacarpal; mcII, metacarpal II; mcIII, = metacarpal III; ra, radius; rad, radiale; rib, rib; sb, scapular blade; sc, scapula; st, sternum; sup, insertion scar for m. supracoracoideus, syn, synsacrum; tbt, tibiotarsus; tc tuberculum coracoideum; tv, thoracic vertebra; uln, ulnare, vt, vertebra (position uncertain). Scale bars equal 10 mm.
Holotype
NMNZ S.47114: nearly complete right humerus and distal portion of left femur. Measurements in Table 1.
Diagnosis
Differs from all other Paleocene penguins in strongly flattened humerus shaft (midshaft width more than twice midshaft depth). Differs from Muriwaimanu, Sequiwaimanu, and Kupoupou in wider fossa formed by impressio m. pectoralis. Differs from Muriwaimanu, Sequiwaimanu, and Crossvallia in more distally extended insertion scar for m. supracoracoideus. Differs from Kaiika in proportionally wider humerus shaft, greater proximodistal length of crista bicipitalis, and more proximodistally elongate scar for m. supracoracoideus. We note that the deep groove on the tuberculum ventrale of Kaiika maxwelli, which was originally considered autapomorphic for that species, is more likely to be an artifact of incomplete preservation of the natural mold used to cast the holotype humerus.
Occurrence
Late Paleocene Moeraki Formation, Hampden Beach, North Otago, New Zealand.
Description
The holotype humerus is strongly sigmoid in shape (Fig. 5). It is only slightly longer (estimated complete length 150 mm) than the humerus of Kaiika maxwelli (146.8 mm), but the shaft is ~50% wider and is more strongly dorsoventrally flattened. As in Kumimanu and Kaiika, the fossa formed by impressio m. pectoralis is extremely wide. It is deep near its proximal end, becoming shallower distally. The m. supracoracoideus insertion scar is strongly raised, although less so than in earlier Paleocene taxa such as Muriwaimanu and Sequiwaimanu. The scar extends distally to the level of the rim of the fossa tricipitalis and is positioned close to the midline of the shaft as in Muriwaimanu and Sequiwaimanu, whereas it is shifted towards the caudal border of the shaft in Kumimanu and most other penguins. The distal margin of the crista bicipitalis forms a convex projection, which resembles that of Muriwaimanu tuatahi and Kaiika maxwelli. The insertion scar for m. coracobrachialis caudalis is short and ovoid, oriented almost craniolaterally, and clearly separated from the head. In turn, a platform of bone proximal to the scar separates the incisura capitis from the fossa tricipitalis dorsalis, which is deep, but occupies a small area due to being impinged upon by the m. supracoracoideus insertion scar. The distal end, including the condyles and trochlear processes, is missing.
NMNZ S.46081 (Fig. 6.1–6.5) is the most informative referred specimen. The intact portion of the sternum measures 177 mm, but it is unfortunately lacking both its cranial and caudal margins. Because no indication of the incisura caudalis is visible along the lateral margin, the sternum must have been considerably longer when complete. A prominent intramuscular line is developed. This feature varies in prominence in extant penguins. Based on the intact left side near mid-keel, the sternum width would have been within the range of the extant emperor penguin despite its greater length. These proportions together suggest an elongate and slender sternum, resembling that of Kairuku in general proportions (see Ksepka et al., Reference Ksepka, Fordyce, Ando and Jones2012, fig. 3).
The radiale is more robust and mediolaterally wider than in extant penguins. The ulnare is greatly expanded, but has a disc-like shape, unlike the sharp triangular shape present in crown penguins. The carpometacarpus is slightly shorter than the species average for emperor penguins. Complete skeletons of Paleocene penguins are rare, but the wing proportions in associated skeletons of Muriwaimanu and Sequiwamanu indicate that the carpometacarpus was proportionally shorter relative to the humerus, as in other early stem penguins. Based on the proportions of Muriwaimanu and Sequiwamanu, the carpometacarpus length of NMNZ S.46081 (proximal margin to distal margin of metacarpal II: 70.8 mm) falls within the size expected for Petradyptes stonehousei based on humerus length in the holotype (predicted range 68.5–74.4 mm). The proximal end of the carpometacarpus is partially embedded in matrix and is exposed in dorsal view (Fig 6.2), whereas the distal end of the carpometacarpus projects free from the matrix (Fig. 6.4). As in other Paleocene penguins, the proximal margin of metacarpal I is distinctly separated from the proximal rim of the trochlea carpalis, whereas in Eocene and later penguins it is shifted to nearly the same level. Metacarpal I bears a facet for a free alular digit and extends distally so as to overlap the spatium intermetacarpale, as in Sequiwaimanu and later penguins, but unlike in Muriwaimanu in which metacarpal I is more proximally restricted. Metacarpal II is straight, lacking the cranial bowing present in Eocene and later penguins. There is no indication of a sulcus tendineus. A wedge-shaped cranial expansion is present at the distal end of metacarpal II. Metacarpal III bears a small, ventrally projecting flange near its proximal end, and projects far distally to metacarpal II, unlike in other Paleocene penguins. As in other Paleocene penguins and the Eocene Perudyptes, the articular face of metacarpal II is rounded, whereas it is compressed in more crownward penguins.
A portion of the cranial section of the pelvis is preserved, including part of the synsacrum and the pre-acetabular portion of the right ilium. The ala preacetabularis ilii is quite wide compared to that in modern penguins. The femur lacks the distal condyles, but the preserved section indicates it was slightly longer yet slightly more slender than the femur of the emperor penguin. At the proximal end, the neck is nearly as wide as the head and the fovea ligamentum capitis is oriented dorsomedially as in most penguins (oriented medially in Crossvallia unienwillia). The shaft is straight and the crista trochanteris is moderately projected. The proximal portions of the tibiotarsus and fibula are preserved in articulation. The crista cnemialis cranialis has a strong square shape, as in Crossvallia waiparensis Mayr et al., Reference Mayr, De Pietri, Love, Mannering and Scofield2019, and is more laterally deflected than in extant penguins (Fig. 6.5). The crista cnemialis has a slight proximal hook, which is absent in Kumimanu. In both features, the tibiotarsus differs from that of extant penguins and more closely resembles the tibiotarsus of some Procellariiformes.
NMNZ S.47146 (Fig. 6.6, 6.7) closely resembles NMNZ S.46081 in size, the presence of a wide iliac blade, and the proportions of the nearly complete femur. This specimen also preserves two complete thoracic vertebrae and the caudal margin of the scapular blade. The vertebra preserved closest to the synsacrum presumably represents the caudal-most thoracic element. The corpus is weakly opisthocoelous, and a large ovoid fossa is present on the exposed lateral surface (Fig. 6.7). A second weakly opisthocoelous thoracic vertebra is preserved nearby and likely represents the penultimate thoracic element. As in Kumimanu, the crista spinosa synsacri is broad and rounded, in contrast to the thinner, sharper shape in modern penguins. The scapular blade is roughly half as wide as the blade in emperor penguins, despite the larger overall size of the specimen, reflecting the lack of the extreme “paddle-like” distal expansion seen in extant penguins.
NMNZ S.47933 (Fig. 6.8–6.10) includes a large number of elements, most of which are incomplete. Three cervical vertebrae and two thoracic vertebrae are preserved. A large portion of the left scapula is preserved, but unfortunately the omal end and caudal end are both missing. The preserved portion of the blade is strap-like. The proximal end of the right humerus is preserved. The intact section is nearly identical in width to the Petradyptes stonehousei holotype humerus, and closely resembles it in exhibiting a very wide fossa formed by the impressio m. pectoralis (Fig. 6.9) and a strongly raised insertion scar for m. supracoracoideus that is positioned near the midline of the shaft (Fig. 6.10). The shaft of the humerus is exposed in cross-section slightly distal of the margin of the m. supracoracoideus insertion scar. At a coarse scale, the histological profile resembles that of Eocene penguin humeri from Seymour Island, Antarctica, in having a larger medullary cavity than in modern penguins (Ksepka et al., Reference Ksepka, Werning, Sclafani and Boles2015). The tibiotarsus resembles that of NMNZ S.47146 but is too worn to provide additional information.
NMNZ S.47927 (Fig. 6.11–6.13) includes several elements, most badly truncated. A portion of the femoral shaft is intact and closely matches the size of other specimens provisionally assigned to Petradyptes stonehousei (Table 1). Likewise, the width of the distal end of the right carpometacarpus (~20 mm) is consistent with the same measurement in referred specimen NMNZ S.46081 (21.1 mm). As in that specimen, metacarpal II bears a cranial expansion near its distal tip and metacarpal III projects distal to the level of metacarpal II (Fig. 6.13). Metacarpals II and III are both broken so as to expose their cross-section just distal to midshaft, which reveals both are nearly cylindrical as in Muriwaimanu and Sequiwaimanu, as opposed to the strongly flattened cross-section in more crownward penguins. The medullary cavity is very small in both metacarpals. The scapula is preserved in two sections: the omal portion closely resembles that of Kumimanu, and a truncated segment of the blade suggests that it was modestly expanded. The ala postacetabularis ilii is exposed in medial view and tapers to a triangular point. The distal quarter of the left tibiotarsus is preserved in cranial view, although the distal condyles are badly worn. At the level of the break, the shaft is mediolaterally widened as in Kumimanu biceae (versus less expanded in crown penguins) and is slightly concave on its cranial face and convex on its caudal face. The medullary cavity is small and surrounded by thick compacta.
Etymology
In honor of Bernard Stonehouse, in recognition of his landmark contributions to the study of penguins, as well as his work on other New Zealand birds and his generous encouragement of fossil penguin researchers.
Material
NMNZ S.46081: partial sternum, associated ribs, left radiale, complete left carpometacarpus, partial synsacrum and right ilium, right femur missing distal end, and proximal ends of right tibiotarsus and fibula. NMNZ S.47146: two thoracic vertebrae, several ribs, partial scapula, partial pelvis, nearly complete left femur. NMNZ S.47927: fragment of synsacrum, multiple ribs, partial scapula, distal end of right carpometacarpus, postacetabular portion of ilium, femoral shaft, distal end of left tibiotarsus. NMNZ S.47933: three cervical and two thoracic vertebrae, five partial ribs, partial left scapula, proximal end of right humerus, proximal end of right tibiotarsus. One fragment of bone appears to represent part of the sternum but is too incomplete to verify.
Remarks
Fordyce (Reference Fordyce, Vickers-Rich, Monaghan, Baird and Rich1991) briefly described a crushed tibiotarsus (OU 8743), which could potentially belong to Petradyptes stonehousei. The width of the distal end of the tibiotarsus is 34.0 mm in OU 8743, which compares well to the width of >30.7 mm (margins partially lost to erosion) in NMNZ S.47927.
Sphenisciformes indet.
Figure 7
Occurrence
Late Paleocene Moeraki Formation, Hampden Beach, North Otago, New Zealand.
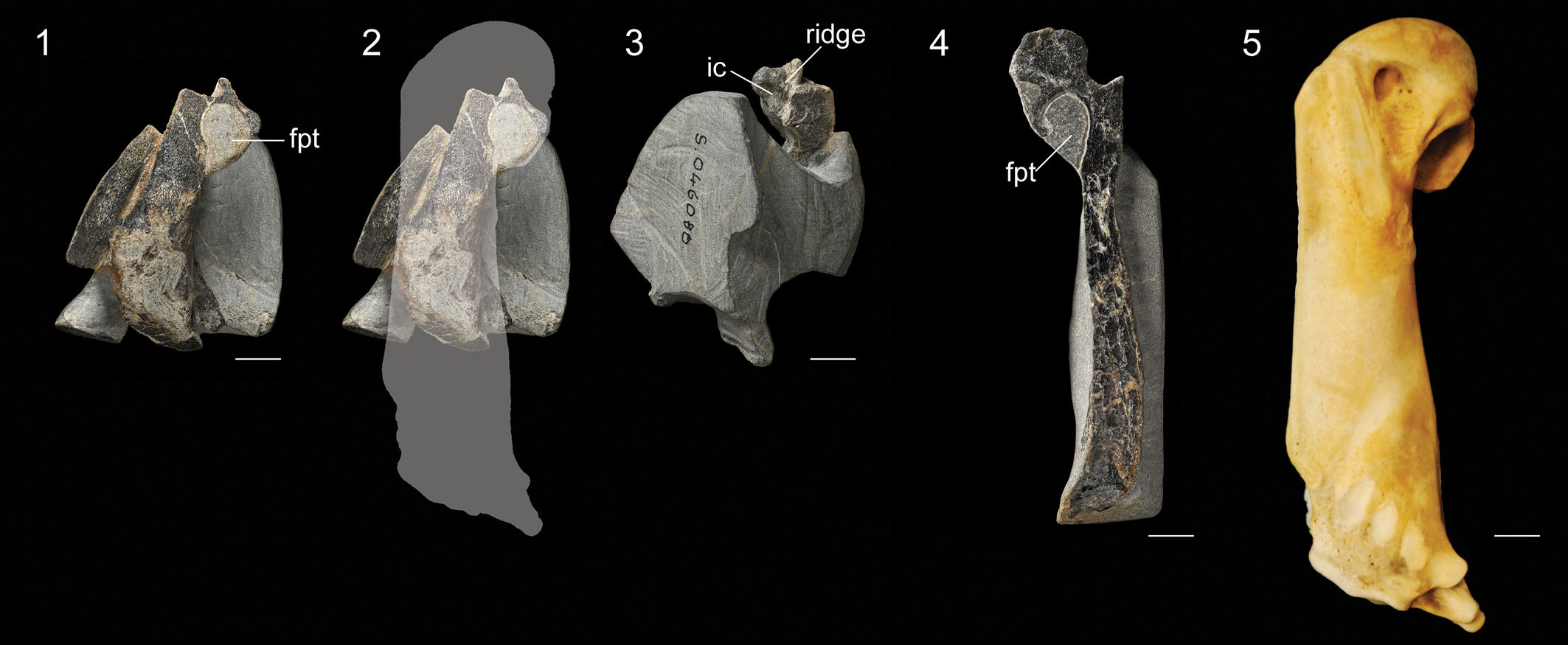
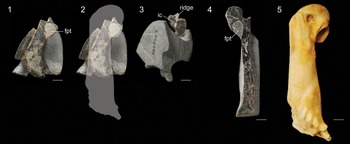
Figure 7. Specimens of Sphenisciformes ident. NMNZ S.46080: (1) humerus exposed in partial longitudinal cross-section, (2) same image with outline to indicate region preserved and estimated overall size, and (3) oblique view of proximal end. NMNZ S.47665: (4) humerus exposed in partial longitudinal cross-section. (5) Humerus of an extant emperor penguin (Aptenodytes forsteri, AMNH 8110) to indicate relative scale. Abbreviations: ic = incisura capitis; fpt = fossa pneumotricipitalis; ridge = ridge dividing the incisura capitis and sulcus ligamentum. Scale bars equal 10 mm.
Description
NMNZ S.46080 includes a fragmentary left humerus, which is missing the distal two thirds of the shaft due to erosion. This exposes the fossa tricipitalis, which is relatively large (Fig. 7.1). A portion of the ventral surface of the head is intact, which reveals that a ridge was present dividing the incisura capitis and sulcus ligamentum transversus (Fig. 7.3). This ridge is present in most Paleocene penguins (e.g., Muriwaimanu, Crossvallia, Kupoupou), although it is absent in Sequiwaimanu. Shaft width, which can be measured near the presumed midshaft, is ~90% the length of the species average for king penguins and 105% that of Sequiwaimanu rosieae. Accounting for the more slender shaft proportions of Paleocene stem penguins, this suggests an individual intermediate in size between the king penguin and emperor penguin, the two largest extant penguin species. A wide curved element preserved adjacent to the humerus may represent a portion of the furcular ramus.
NMNZ S.47665 (Fig. 7.4) is a right humerus, exposed in longitudinal cross-section. Little information can be gleaned from this specimen other than the fossa tricipitalis appears to have been undivided as in nearly all stem penguins. No meaningful measurements can be obtained, but the overall proportions suggest this humerus was very close in size to NMNZ S.46080.
Material
NMNZ S.46080: proximal end of left humerus, one rib, and two unidentified fragments of bone. NMNZ S.47665: right humerus lacking distal end.
Remarks
These specimens are too incomplete to justify erecting a new taxon. We consider it likely they belong to the same species based on size, but this cannot be confirmed due to their incompleteness. Regardless, these smaller fossils support the presence of a fourth distinct penguin species in the Moeraki Formation.
Phylogeny of Paleocene penguins
The parsimony analysis resulted in 7,128 MPTs of 601 steps (CI = 0.628, RI = 0.861, RC = 0.541). The strict consensus tree (Fig. 8.1) contains a polytomy including Kumimanu biceae, Kumimanu fordycei n. sp., and a clade uniting Kaiika maxwelli and Petradyptes stonehousei n. gen. n. sp. The polytomy is recovered one node crownward of a polytomy including Crossvallia unienwillia and Crossvallia waiparensis, and one node stemward of Kupoupou stilwelli. The relationships of the remaining fossil and extant taxa largely conform to previous results (e.g., Degrange et al., Reference Degrange, Ksepka and Tambussi2018; Thomas et al., Reference Thomas, Tennyson, Scofield, Heath, Pett and Ksepka2020b), so we focus on the relationships of Paleocene species in the discussion below.
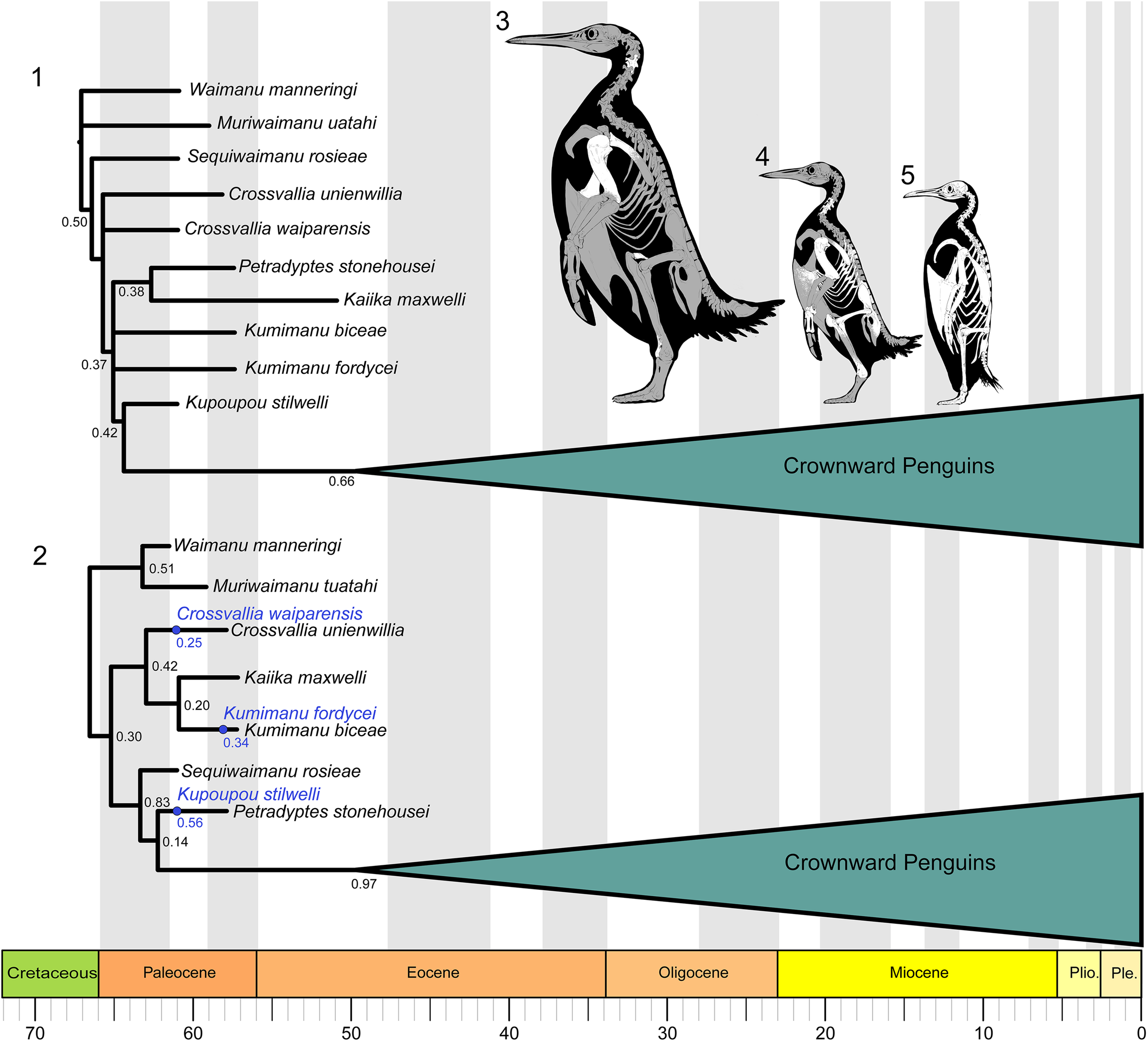
Figure 8. Phylogenetic relationships of Paleocene penguins. (1) Strict consensus 7,128 MPTs of 601 steps (CI = 0.628, RI = 0.861, RC = 0.541) from parsimony analysis, with bootstrap support indicated. (2) Maximum sampled-ancestor clade credibility tree summarized from the Bayesian combined-evidence analysis in RevBayes. Posterior probabilities are shown for internal nodes. Sampled ancestors are colored blue and posterior probabilities for the MSACC sampled ancestor positions in this tree are shown in blue. Skeletal reconstructions are provided to show the relative sizes of (3) Kumimanu fordycei n. sp., (4) Petradyptes stonehousei n. gen. n. sp., and (5) the extant emperor penguin Aptenodytes forsteri. Bones preserved in the fossil specimens described in this paper are indicated in white. Skeletal reconstructions by Simone Giovanardi.
In the parsimony results, a limited number of unambiguous synapomorphies can be identified at each node, due in part to the fact that Kaiika maxwelli is known only from the humerus. The clade uniting Petradyptes, Kaiika, Kumimanu, and crownward penguins to the exclusion of Crossvallia is united by (174:1) extension of the insertion scar for m. supracoracoideus to the level of the distal rim of the fossa tricipitalis. One additional character state, (240:0) sulcus extensorius of tibiotarsus placed close to midline, represents an ambiguous synapomorphy for this clade because it cannot be scored for Kaiika or Petradyptes. The clade uniting Kupoupou and more crownward penguins to the exclusion of Petradyptes, Kaiika, and Kumimanu is united by three unambiguous characters: (162:0) proximal end of humerus narrow, (171:1) impressio m. pectoralis forming narrow fossa, and (200:1) processus condylaris dorsalis of ulna reduced. The first two character states, however, constitute reversals. Two features of the tarsometatarsus may represent synapomorphies of Kupoupou and more crownward penguins (or a more inclusive clade): (247:2) strongly shortened tarsometatarsus (length <2.5 times width), and (261:1) metatarsal IV straight. The precise distribution of these features cannot be evaluated at present, because the tarsometatarsus remains unknown for Petradyptes, Kaiika, and Kumimanu.
A single character supports the sister group relationship between Kaiika maxwelli and Petradyptes stonehousei: (160:2) humerus midshaft width more than twice midshaft depth. Although our analysis recovered equally parsimonious topologies uniting Kumimanu biceae and Kumimanu fordycei in an exclusive clade or placing them as sequential branches along the penguin stem lineage, we consider it much more likely they represent sister taxa based on their very large size and overall similarity. We also note that one character suggests that Kumimanu may occupy a slightly more crownward position than Petradyptes. As in the stemward taxa Waimanu, Muriwaimanu, and Sequiwaimanu, the caudal thoracic vertebrae of Petradyptes bear ovoid lateral depressions. An isolated thoracic vertebra of Kumimanu biceae lacks such depressions, as do all more crownward penguins. If this character state were to be confirmed in Kumimanu fordycei, it would result in both Kumimanu species being shifted to a position one node crownward of Petradyptes + Kaiika. Likewise, we note that the cnemial crests more closely resemble those of Procellariiformes in Petradyptes than in the specimen we tentatively assigned to Kumimanu biceae. This feature is not represented by a discrete character in our character matrix, but is also consistent with Petradyptes occupying a more basal branch than Kumimanu.
Our Bayesian analysis yielded alternate placements for several taxa. In the maximum sampled-ancestor clade credibility (MSACC) tree (Fig. 8.2), the strongest point of conflict with our parsimony analysis regards the placement of Sequiwaimanu crownward of Crossvallia and Kumimanu in the MSACC tree, versus a position near the root of the tree in the parsimony strict consensus tree. Only one character is optimized as supporting the more crownward placement in the MSACC tree: (162:0) humeral proximal width less than twice midshaft depth. Crossvallia and Kumimanu are both recovered as monophyletic, and Crossvallia, Kumimanu, and Kaiika form a clade as well, unlike in the parsimony result. This clade is supported by one feature, (171:2) impressio m. pectoralis forming a wide fossa (convergently acquired in Petradyptes), and is united with all more crownward penguins to the exclusion of Waimanu and Muriwaimanu by one feature: (256:1) proximal vascular foramina of tarsometatarsus widely separated. Kupoupou and Petradyptes are potentially supported in a more crownward position than other Paleocene penguins by three features, but all are ambiguous because they cannot be scored in Sequiwaimanu or Petradyptes: (247:2) tarsometatarsus greatly shortened (length <2.5 times width), (257:0) foramen proximale vascularis medialis opens distal to crista hypotarsi medialis, and (261:1) lateral margin of metatarsal IV straight.
The parsimony tree suffers from several collapsed nodes due to non-overlap of character codings, whereas the MSACC tree is fully resolved, but may present a false sense of precision because many nodes are only weakly supported. Overall, both trees require some homoplasy to explain the distribution of characters related to the flipper morphology of Paleocene penguins. The parsimony tree implies homoplasy in humerus proximal width and the development of the incisura capitis, whereas the Bayesian tree implies homoplasy in the presence of thoracic pleurocoels, shaft flattening, and placement of the insertion for m. supracoracoideus. Both trees also imply homoplasy in the shape of the impressio m. pectoralis. We speculate that discovery of more complete fossils, in particular specimens preserving the distal portions of the flipper and the tarsometatarsus, most likely will result in additional support for placing Sequiwaimanu close to the root of the tree as in the parsimony result, but also for the monophyly of the genera Kumimanu and Crossvallia as in the Bayesian topology.
Body mass in fossil penguins
Regressions
The humerus of Kumimanu fordycei n. sp. is the largest yet reported for a penguin. We developed a set of allometric equations to enable the estimation of penguin body mass from humeral dimensions in order to obtain an estimate of body mass for the new species. An allometric equation in this context is a linear equation fitted to measurements of two independently measured traits after they have been log transformed, sensu Gould (Reference Gould1966) and Riska (Reference Riska1991). Due to a combination of historical inertia and the diagnostic nature of the humerus and tarsometatarsus, the holotypes of almost all fossil penguin species incorporate either one or both of these elements (Marples, Reference Marples1960; Fordyce and Jones, Reference Fordyce, Jones, Davis and Darby1990). Thus, we anticipate that sphenisciform body mass regressions based on humeral dimensions may be of considerable use in future studies.
Previous work exploring forelimb-based body-mass correlates for extant and fossil crown birds did not incorporate penguins or other flightless taxa (Field et al., Reference Field, Lynner, Brown and Darroch2013; Pittman et al., Reference Pittman, Heers, Serrano, Field, Habib, Dececchi, Kaye and Larsson2020). In order to estimate the original body mass of Kumimanu fordycei, three allometric equations were derived using ordinary least squares regression to predict body mass from humerus dimensions in extant penguins, based on measurements obtainable in the K. fordycei holotype: maximum humerus proximal width (HPW), maximum humerus length (HL), and humerus midshaft width (HSW).
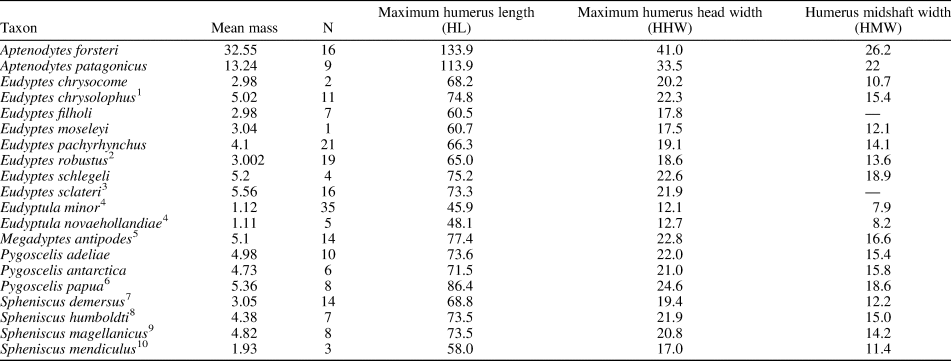
Our extant penguin body mass dataset includes humerus dimensions from 216 specimens from 20 species, measured using digital calipers sensitive to 0.1 mm. From these specimens we recorded 216 HPW measurements, 214 HL measurements, and 101 HSW measurements. The holotype humerus of Kumimanu fordycei measures 233 mm as preserved, however distortion of the head exaggerates its proximal projection (artificial addition estimated at 3.4 mm), and conversely the distal portions of the trochlear ridges are missing (estimated loss of 7 mm length). When these two factors are corrected, we infer a complete length of 236.6 mm.
We gathered body mass data primarily from the species accounts of Borboroglu and Boersma (Reference Borboroglu and Boersma2013). Penguin body mass is known to fluctuate considerably across the breeding cycle, so it is important to choose a standard point at which to compare values across species. We selected “arrival at colony” masses for this purpose both because it represents a relatively discrete time interval in the breeding cycle, and because body masses for penguins are often obtained at this stage because researchers can capture penguins easily when they are on land. Sources for species for which body masses at the arrival at colony stage were not available from Borboroglu and Boersma (Reference Borboroglu and Boersma2013) are listed in Table 2. The natural log of species-averaged body mass was used against the natural log of humerus measurements from individual birds in the regression analyses.
Table 2. Average body mass and humerus measurements of extant penguins. Body mass is based on average of male and female “arrival at colony” values from Borboroglu and Boersma (Reference Borboroglu and Boersma2013) unless otherwise indicated. Notes on body mass: 1—Average of “pre-egg laying” Heard Island population. 2—Average of parents of fledging stage chicks. 3—Average of “arrival” females (data on males not available). 4—Species average from Dunning (Reference Dunning2008). 5—Average “breeding” weight from Moore et al. (Reference Moore, Douglas, Mills, McKinley, Nelson and Murphy1991) as presented by Williams (Reference Williams1995). 6—Average of “nest building” weight from Williams (Reference Williams1995). 7—Average “breeding” weight from Williams (Reference Williams1995). 8—Weights listed as “wild penguins at colony” potentially including post-arrival individuals. 9—Average body mass at arrival from 5-year survey reported in Williams (Reference Williams1995). 10—Mid-incubation values from Williams (Reference Williams1995). Body mass reported in kilograms. Humerus measurements reported in millimeters, taken directly. Regression analyses used the natural log of species-averaged body mass against the natural log of humerus measurements from individual birds and not the species-averaged humerus measurements shown here.
The largest previously reported penguin humerus belongs to the holotype specimen of Kumimanu biceae. This specimen lacks an indeterminate portion of its distal end, which resulted in estimates of its total length ranging from 185 mm to 228 mm based on extrapolations using the humerus proportions of other stem penguins (Mayr et al., Reference Mayr, Scofield, De Pietri and Tennyson2017a). If the proportions of Kumimanu biceae matched those of Kumimanu fordycei, the humerus of the former would have approached the upper range of these estimates at 218 mm maximum length. We used this length when estimating the body mass of K. biceae. Mass estimates for these and other “giant” penguins analyzed in this study represent extrapolations from our regression models, due to the exceptionally large sizes of these fossils relative to extant penguins.
Mass estimates
The strength of correlation between skeletal measurements and body mass was assessed with a coefficient of determination (R 2) obtained from ordinary least squares regression. Maximum humerus proximal width and maximum humerus length provided accurate estimates of body mass in extant penguins (both exhibiting R 2 of 0.95). Both relationships were significant, with p-values <0.001. All emperor penguin specimens had high leverage on the HPW regression based on hat matrix values, and all emperor and several king penguin and Eudyptula specimens had high leverage on the HL regression. The equation we derived for estimating body mass (BM) from measurements of maximum humerus proximal width is:
Ln(BM) = 2.5857(Ln(HPW)) + 0.5394
The equation derived for maximum humerus length is:
Ln(BM) = 2.9613(Ln(HL)) − 4.2823
Mean body mass estimates derived for Kumimanu fordycei were 159.7 kg based on maximum humerus proximal width (142.6 kg and 178.8 kg as the lower and upper 95% confidence interval bounds, respectively), and 148.0 kg based on maximum humerus length (132.5 kg and 165.3 kg, lower and upper 95% confidence interval bounds, respectively). We therefore estimate that this species would have exhibited a colony arrival mass in the range of ~155 kg when alive—thus ranking as the heaviest penguin yet discovered.
We also calculated a body mass regression based on humerus midshaft width:
Ln(BM) = 2.5849(Ln(HSW)) + 1.5219
Our regression based on midshaft width yielded a poorer fit (R 2 = 0.8843, p-value <0.001) than those based on humerus length or proximal width, and resulted in a much lower body mass estimate of 66.3 kg for Kumimanu fordycei. Beyond the reduced fit of this equation, as indicated by its lower R 2 value, this midshaft width regression will inevitably be biased towards erroneously low estimates of body size for early stem penguins, in which the humeral shaft is less flattened than in later penguins including the crown group, resulting in a greater thickness but a lesser width for a given overall size. This results in a large disparity between estimates from midshaft width and other indices for early stem penguins (e.g., the mass estimates from humerus length and humerus proximal width are more than six times greater than the estimate from midshaft width for Muriwaimanu). We therefore consider results from the midshaft width regression unreliable for estimating the body masses of Paleocene stem penguins such as those described here. Body mass estimates from all three regressions for 40 fossil taxa are presented in Table 3.
Table 3. Comparative dimensions of fossil penguin humeri. Sources for specimens not measured directly: 1—Jadwiszczak, Reference Jadwiszczak2006; 2—Mayr et al., Reference Mayr, Scofield, De Pietri and Tennyson2017a; 3—Acosta Hospitaleche et al., Reference Acosta Hospitaleche, Reguero and Santillana2017; 4—Acosta Hospitaleche, Reference Acosta Hospitaleche2005; 5—Jadwiszczak et al., Reference Jadwiszczak, Acosta Hospitaleche and Reguero2013; 6—Mayr et al., Reference Mayr, De Pietri, Love, Mannering and Scofield2019; 7—Simpson, Reference Simpson1981; 8—Chávez Hoffmeister et al., Reference Hoffmeister, Carrillo Briceño and and Nielsen2014; 9—Acosta Hospitaleche, Reference Acosta Hospitaleche2014; 10—Chávez Hoffmeister, personal communication, 2016; 11—Emslie et al.,Reference Emslie and Guerra Correa2003; 12—Göhlich, Reference Göhlich2007. *The humerus is weathered and has lost at least part of the periosteum, so this value may be a modest underestimate of true width and hence body mass. **Note the table of Mayr et al., Reference Mayr, Scofield, De Pietri and Tennyson2017a, indicates a width of 70 mm whereas the text (Mayr et al., Reference Mayr, Scofield, De Pietri and Tennyson2017a, p. 4) indicates 75 mm. The latter value is correct.
Discussion
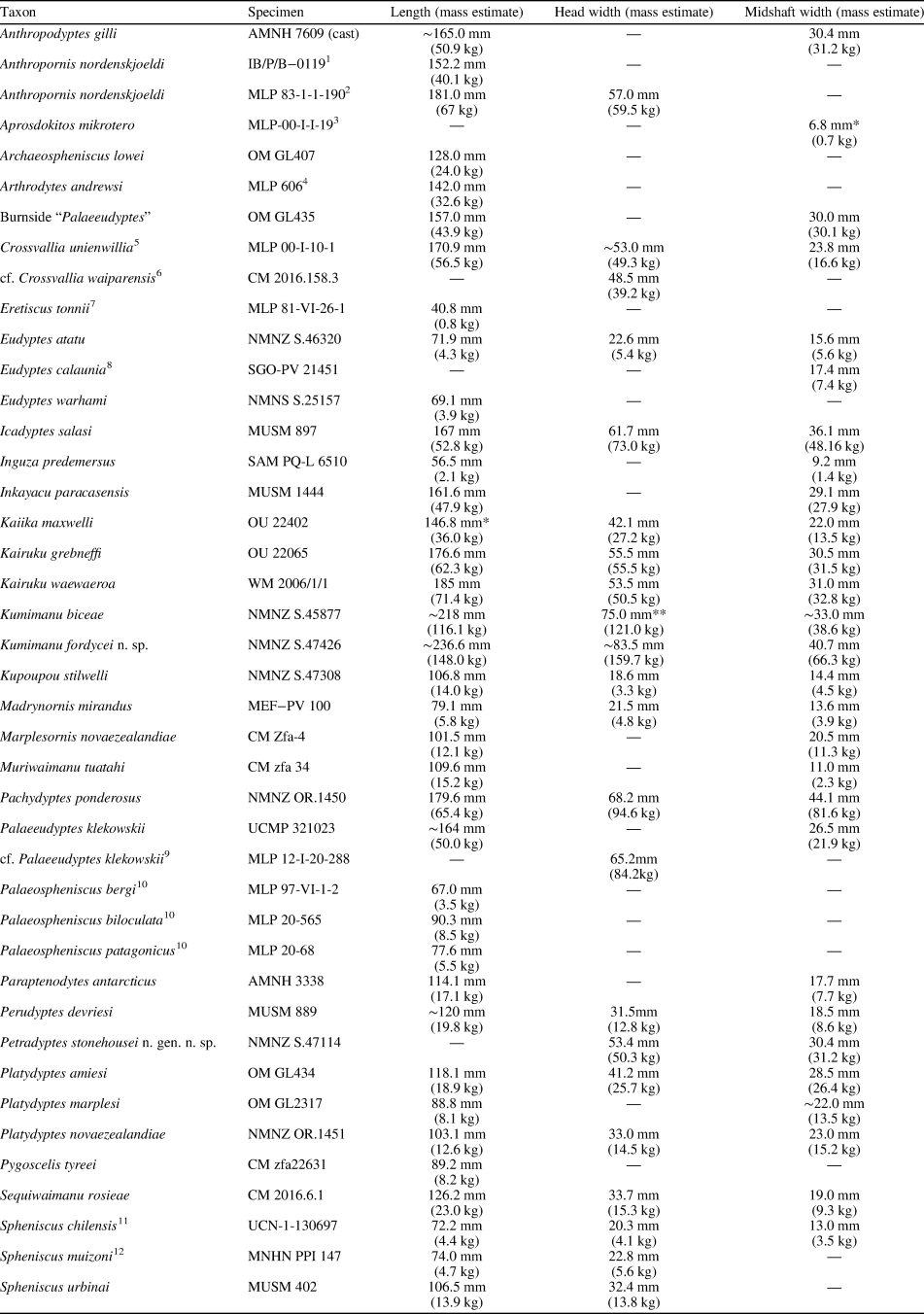
Intriguingly, some of the features of the humerus that distinguish Kumimanu and Petradyptes n. gen. from more crownward penguins resemble those in volant underwater divers such as diving petrels (Procellariiformes: Pelecanoides) and auks (Charadriiformes: Alcidae) (e.g., Smith, Reference Smith2011; Mayr et al., Reference Mayr, De Pietri, Love, Mannering, Bevitt and Scofield2020; Watanabe et al., Reference Watanabe, Field and Matsuoka2021). Similarities between the flight musculature of Paleocene penguins, diving petrels, and alcids are suggested by development of the insertion scars of key muscles involved in the upstroke and downstroke, including the wide impressio m. pectoralis, strongly raised insertion scar for m. supracoracoideus, and less distally elongated insertion scar for m. supracoracoideus scar (Figs. 9, 10). Although not well described, the m. latissimus dorsi scar in early Paleocene penguins appears to have resembled that of auks in being weak and lineate, as opposed to the strong circular insertion observed in later penguins (best observed in Sequiwaimanu). These similarities to distantly related taxa capable of both aerial flight and wing-propelled diving suggest that despite being clearly flightless, early giant penguins such as Kumimanu may have retained some ancestral features that were once necessary for aerial flight but less optimal for underwater flight. Thus, it is likely that Paleocene stem penguins employed an underwater flight stroke that was less efficient than that of later penguins. Testing this idea quantitatively will require future functional analyses.
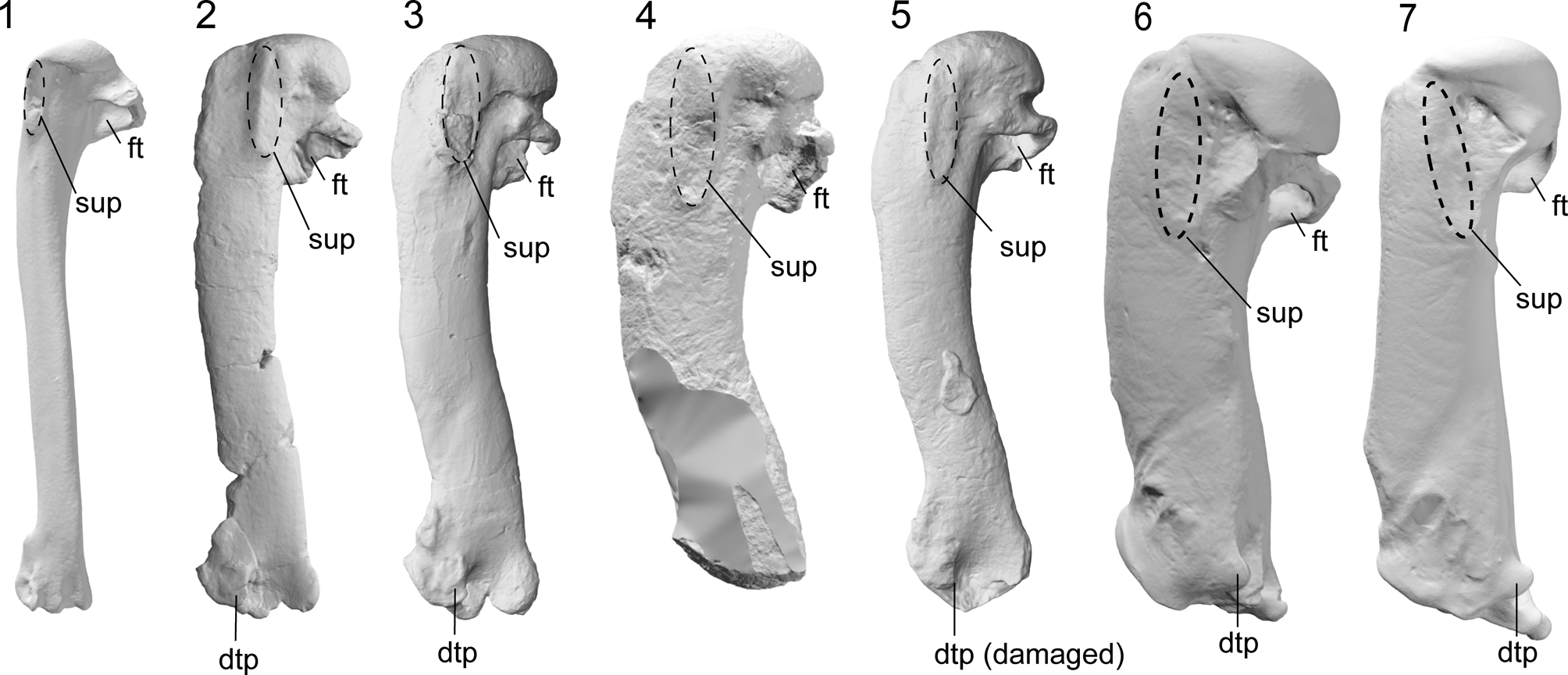
Figure 9. Comparative images of the humerus in dorsal view (swimming posture): (1) the auk (Alcidae) Alca torda Linnaeus, Reference Linnaeus1758 (NMNZ OR.12282) and the penguins (2) Muriwaimanu tuatahi (OU12651), (3) Sequiwaimanu rosieae (CM 2016.6.1), (4) Petradyptes stonehousei n. gen. n. sp. (NMNZ S. 47114), (5) Kaiika maxwelli (OU22402), (6) Pachydyptes ponderosus (NMNZ OR.1450), and (7) Aptenodytes forsteri (NMNZ OR.23039). Abbreviations: dtp, dorsal trochlear process; ft, fossa tricipitalis; sup, insertion scar for m. supracoracoideus. Not to scale.
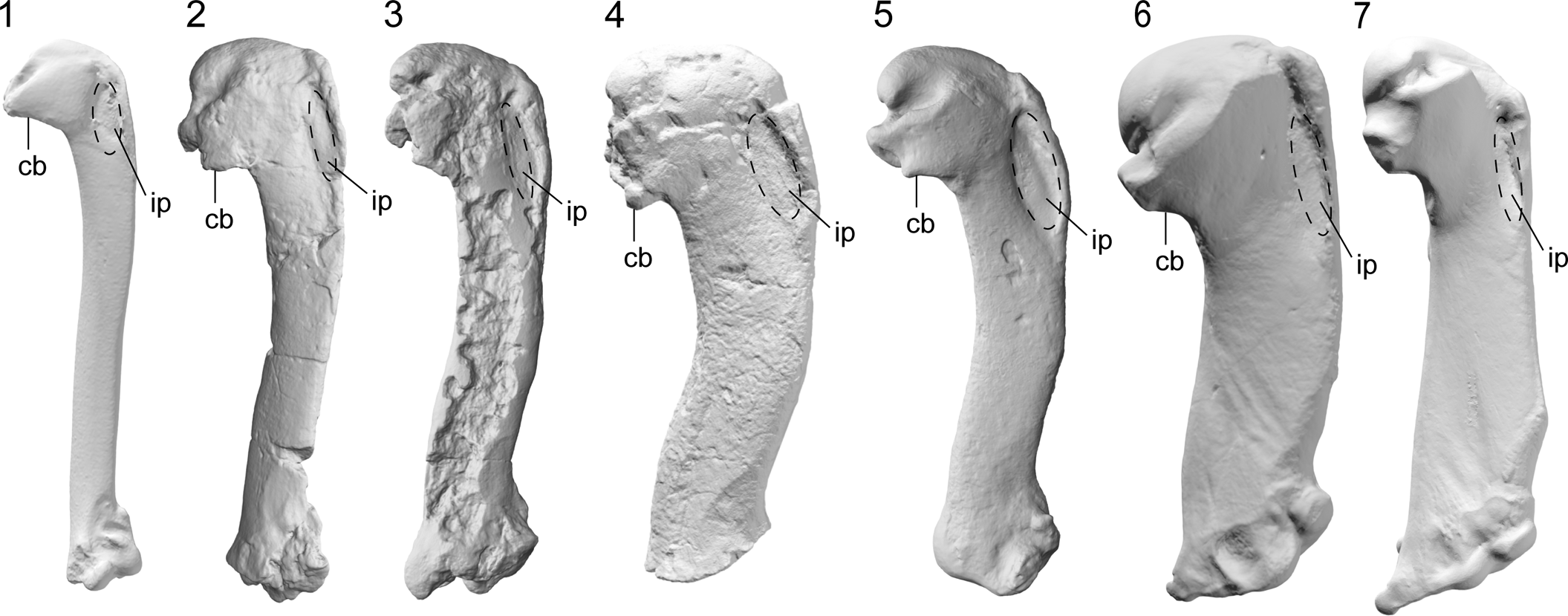
Figure 10. Comparative images of the humerus ventral view (swimming posture): (1) the auk (Alcidae) Alca torda (NMNZ OR.12282) and the penguins (2) Muriwaimanu tuatahi (OU12651), (3) Sequiwaimanu rosieae (CM 2016.6.1), (4) Petradyptes stonehousei n. gen. n. sp. (NMNZ S. 47114), (5) Kaiika maxwelli (OU22402), (6) Pachydyptes ponderosus (NMNZ OR.1450), and (7) Aptenodytes forsteri (NMNZ OR.23039). Abbreviations: cb, crista bicipitalis, ip, impressio m. pectoralis. Not to scale.
Previous attempts to estimate body mass for extinct penguins relied primarily on hindlimb elements (Jadwiszczak, Reference Jadwiszczak2001; Jadwiszczak and Mörs, Reference Jadwiszczak and Mörs2011). While hindlimb dimensions are indeed useful for estimating avian body masses (e.g., Campbell and Marcus, Reference Campbell and Marcus1992; Field et al., Reference Field, Lynner, Brown and Darroch2013), many of the largest reported penguin specimens do not preserve hindlimb material. Our regressions suggest that humerus dimensions also provide reliable estimates of body mass for penguins. Because the underwater flight stroke provides the primary means of penguin locomotion, there is good reason to suspect that the humerus, which serves as the primary attachment point for the major muscles involved in the upstroke and downstroke, may scale relatively tightly to the body mass of the bird that the muscles must propel through the water.
The humerus of Kumimanu fordycei n. sp. is the largest penguin humerus yet discovered, both in terms of its overall length and proximal width. Only the late Eocene Pachydyptes ponderosus Oliver, Reference Oliver1930, has a larger midshaft width, but this primarily reflects the proportionally wider shaft of post-Paleocene taxa (Table 3).
Other very large penguins also have been reported from different elements. The largest previously reported penguin hindlimb element is an isolated tarsometatarsus (MLP 12-I-20-288) from the Eocene La Meseta Formation of Seymour Island, Antarctica. This specimen was assigned to Palaeeudyptes klekowskii Myrcha, Tatur, and del Valle Reference Myrcha, Tatur and del Valle1990, by Acosta Hospitaleche (Reference Acosta Hospitaleche2014), and is ~25% longer than the holotype tarsometatarsus of P. klekowskii, which itself is one of the largest specimens previously known for the species. If correctly referred, this would imply a surprisingly wide range of intraspecific size variation. Regardless of its taxonomic affinities, MLP 12-I-20-288 clearly belonged to an extremely large penguin. Applying regressions based on tarsometatarsus width and depth (Jadwiszczak, Reference Jadwiszczak2001), Acosta Hospitaleche (Reference Acosta Hospitaleche2014) estimated a body mass of 114.4–116.2 kg for this individual. A large partial humerus, preserving only its proximal end (MLP- 12-I-20-288), was also assigned to Palaeeudyptes klekowskii in the same study. Because these bones cannot be directly associated, it remains possible that they belong to two different species, or at least to individuals of different sizes. Our regression based on humerus proximal width (65.2 mm in MLP- 12-I-20-288) yields a body mass estimate of 84.2 kg. In terms of overall length of the humerus itself, scaling using the proportions of smaller complete Palaeeudyptes klekowskii humeri suggests a total length of ~226 mm for this specimen, which would be ~90% the length of the Kumimanu fordycei holotype humerus.
In summary, available comparisons support Kumimanu fordycei as the heaviest known penguin. Future discovery of tarsometatarsi belonging to Kumimanu would allow us to compare mass estimates from Jadwiszczak's (Reference Jadwiszczak2001) regression directly with those from the large Palaeeudyptes tarsometatarsus. We refrain from attempting comparisons of standing height or body length among these taxa, given that very different proportions of stem and crown penguins make such inferences difficult in the absence of nearly complete skeletons (Ksepka et al., Reference Ksepka, Fordyce, Ando and Jones2012).
Nevertheless, the giant body sizes of many stem penguins suggest a foraging niche that is not found in any aquatic bird group today. Because some Paleocene penguins attained giant body size while retaining plesiomorphic flipper features, we speculate that the rapid shift to giant sizes in Paleocene penguins may have been driven primarily by selection for more efficient thermoregulation. Larger endotherms are able to maintain body temperature more efficiently through thermal inertia, and there is a clear positive correlation between body size and both dive depth and dive duration in marine birds (e.g., Boyd and Croxall, Reference Boyd and Croxall1996; Watanuki and Burger, Reference Watanuki and Burger1999; Halsey et al., Reference Halsey, Blackburn and Butler2006). Giant size may have allowed Kumimanu penguins to gain access to foraging areas in colder and deeper waters and to exploit larger prey than smaller contemporaneous penguin taxa.
Although their overall range of body sizes is much lower, the fact that penguins reached their overall peak in body size very early in their evolutionary history stands in marked contrast to patterns observed among marine mammals. Fossil pinnipeds date to the Oligocene but the clade did not reach their peak body sizes until the late Miocene, sirenians date to the early Eocene but did not reach peak body sizes until the Pliocene, and cetaceans date to the early Eocene but only reached peak body size close to the present (Churchill et al., Reference Churchill, Clementz and Kohno2015; Pyenson and Vermeij, Reference Pyenson and Vermeij2016). The unusual trajectory of body size evolution in penguins may be explained in part by the fact that penguins never approached the upper size limits of their marine mammal counterparts, limited by factors such as ovipary. However, another possible factor is that penguins were likely already highly adapted to marine diving before they lost flight, given that all evidence suggests the earliest stem penguins would have been volant wing-propelled divers similar to auks (Simpson, Reference Simpson1946). Thus, the loss of flight would have opened the door for rapid body size increases by removing the competing pressure for limiting weight for take-off.
The early attainment of giant body size by penguins also may have allowed them to take advantage of a limited window of opportunity, which closed as pinnipedimorphs spread through the southern oceans. Giant penguins vanished near the Oligocene-Miocene boundary, and one favored hypothesis is that predation pressure or competition from pinnipedimorphs for food and breeding sites led to selective extinction of very large species whereas smaller penguins thrived from the Miocene onwards (Warheit and Lindberg, Reference Warheit, Lindberg and Burger1988; Warheit, Reference Warheit1992). The lack of spatiotemporal overlap between the largest known fossil penguins and similar-sized marine mammals is consistent with this hypothesis (Ando and Fordyce, Reference Ando and Fordyce2014).
An underappreciated aspect of the early evolution of giant body size in penguins is the potential for improved thermoregulation to increase long-distance dispersal capabilities. Earlier work suggested that penguins originated in Zealandia and dispersed to other continents only after the evolution of the humeral plexus (Thomas et al., Reference Thomas, Ksepka and Fordyce2011). This plexus is only one of several countercurrent heat exchangers present in extant penguins, but is emphasized in paleobiological studies because it is the only one that leaves an osteological correlate that can serve as a proxy of presence/absence in fossil taxa (Thomas et al., Reference Thomas, Ksepka and Fordyce2011). With increasing evidence that giant size also originated while penguins were still restricted to Zealandia, it is interesting to note that the earliest record of a penguin on another continent is the giant Crossvallia unienwillia from Antarctica (Tambussi et al., Reference Tambussi, Reguero, Marenssi and Santillana2005). The humerus of this species is too poorly preserved to infer whether the humeral plexus was present, but the absence of evidence for this structure in Petradyptes, Kaiika, and Kupoupou suggests that the humeral plexus evolved after Crossvallia had already diverged from other penguins. Thus, it is possible that the thermoregulatory benefits of large body size facilitated some key dispersal events in Paleocene penguins, allowing these early titans to explore a wider range of both geographical and ecological space.
Acknowledgments
We thank R.E. Fordyce (OU), R.P. Scofield (CM), M. Florence and H. James (USNM), C. Mehling, M. Norell, J. Cracraft, P. Capainolo, B. Smith, and P. Sweet (AMNH), and R. Prum and K. Zyskowski (YPM) for access to specimens; S. Tennyson, A. Mannering, and J. Wold for their help in finding the Hampden Beach fossils described here; and A. Mannering also for his outstanding work preparing the new specimens. We thank P. Jadwiszczak for sharing photos of Antarctic fossil specimens, K. Dzikiewicz for collecting measurements, C. Kammerer for advice on Greek etymology, and J.-C. Stahl for photography of the new Hampden Beach fossils. DTK was supported by National Science Foundation award DEB-1556615. DJF is supported by UKRI Future Leaders Fellowship MR/S032177/1. TAH and WP were supported by NSF DBI-1759909. The Te Papa Collection Development Fund supported fieldwork, fossil preparation, and storage.
Data availability statement
The morphological dataset and all scripts used for phylogenetic analyses in this study are available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.ttdz08m1r.