


The Neoproterozoic–Palaeozoic transition (NPT) around 600 Ma ago was a critical time interval when the Earth experienced fundamental change, manifested as climatic extremes – ‘snowball Earth’ – followed by the emergence and rapid diversification of animals – ‘Cambrian explosion’. How animals and environments co-evolved, and what caused these fundamental changes to the Earth system during the NPT, is a great scientific puzzle, which has been a rapidly developing frontier of interdisciplinary research between bio- and geosciences. South China preserves a complete stratigraphic succession of the NPT developed in various facies ranging from shallow to deep marine realms with extraordinarily well-preserved, successive fossil biotas in various taphonomic settings (Zhu, Reference Zhu2010; Fig. 1), making it a key area and global focus of studies in the field over recent decades. Indeed, the current narrative of early animal evolution has largely been based on the fossil biotas from South China. These include: (1) the world's oldest microscopic animal fossils with cellular details from the early Ediacaran Weng'an biota (Doushantuo Formation); (2) putative macroscopic animal fossils preserved as carbonaceous imprints from the early Ediacaran Lantian, Wenghui and Miaohe biotas (also Doushantuo Formation); (3) typical late Ediacaran faunas, preserved in dark limestone (Shibantan biota) and as large and poorly mineralized tubular animal fossils (Gaojiashan biota), both from the Dengying Formation; (4) phosphatized small shelly and soft-bodied animal fossils from the early Cambrian Meishucun and Kuanchuanpu faunas; and (5) Cambrian fossil Lagerstätten (Chengjiang, Guanshan and Kaili faunas) with typical Burgess Shale-type soft-bodied preservation.
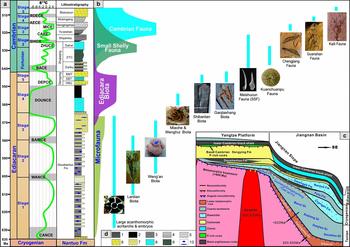
Figure 1. (a) Ediacaran and Cambrian stratigraphy of South China. Ediacaran chemostratigraphy is after Zhu et al. (Reference Zhu, Lu, Zhang, Zhao, Li, Zhao and Zhao2013), and lower Cambrian chemostratigraphy is after Zhu, Babcock & Peng (Reference Zhu, Babcock and Peng2006). Ediacaran lithostratigraphy is based on the successions in the Yangtze Gorges area, Yichang, western Hubei, and lower Cambrian lithostratigraphy is based on the successions in the eastern Yunnan area. (b) Episodic macroevolutionary pattern and distribution of major fossil biotas of South China from Ediacaran and early Cambrian time, showing a long and cryptic prelude and two major radiative episodes of the Cambrian explosion of animals with an evolutionary interlude of the Ediacara biota. Fossil representing the large acanthomorphic acritarchs is Tianzhushania showing cell cleavage within the envelope (image courtesy of P. Liu); fossil image of the Lantian biota is Lantiannella laevis, a putative animal (image courtesy of Z. Chen); fossil image of the Weng'an biota is Eocyathispongia qiania, a sponge-grade animal; fossil image of the Miaohe and Wenghui biotas is Eoandromeda octobrachiata, a putative diploblastic-grade animal; fossil image of the Shibantan biota is Charniodiscus, a characteristic Ediacara organism (image courtesy of Z. Chen); fossil image of the Gaojiashan biota is Conotubus, a large tubular animal (image courtesy of Y. Cai); fossil images of the Meishucun fauna are Anabarites trisulcatus and Protohertzina anabarica; fossil image of the Kuanchuanpu fauna is a phosphatized embryo Olivooides multisulcatus (image courtesy of M. Steiner); fossil image of the Chengjiang fauna is Haikouella lanceolata; fossil image of the Guanshan fauna is Hallucigenia hongmeia; and fossil image of the Kaili fauna is Globoeocrinus globulus. (c) A conceptual tectonic–sedimentary model of the Neoproterozoic – lower Cambrian successions across the Yangtze Platform from shallow shelf to the deep basin. (d) Legend for the lithostratigraphic log: 1 – bedded cherts; 2 – black shale; 3 – silty shale; 4 – siltstone; 5 – sandstone; 6 – thick-bedded and massive dolostone; 7 – thin- or medium-bedded dolostone; 8 – limestone; 9 – phosphorite; 10 – phosphatic nodules.
In addition to their unique archives of early animal evolution, lithologically the composite stratigraphic successions of South China provide the potential to apply various sedimentary geochemical proxies for documenting environmental changes. For example, frequent and dramatic carbon isotope perturbations and ocean redox evolution, evidenced from South China successions, have been suggested to reflect global changes in seawater chemistry that may have been closely related to contemporaneous biological evolutionary events (e.g. Zhu et al. Reference Zhu, Strauss and Shields2007; Li et al. Reference Li, Love, Lyons, Fike, Sessions and Chu2010; Sahoo et al. Reference Sahoo, Planavsky, Kendall, Wang, Shi, Scott, Anbar, Lyons and Jiang2012; Lu et al. Reference Lu, Zhu, Zhang, Shields, Li, Zhao, Zhao and Zhao2013; Shields-Zhou & Zhu, Reference Shields-Zhou and Zhu2013; Chen et al. Reference Chen, Ling, Vance, Shields-Zhou, Zhu, Poulton, Och, Jiang, Li, Cremonese and Archer2015).
However, many questions remain unanswered. In particular, taphonomic or facies biases and poor age constraints, both local and global, hamper direct interpretation of these Chinese palaeobiological and geochemical data. In order to use these archives to their best advantage, South China has been a target area of numerous multidisciplinary research programmes in this field. Following previous thematic issues on the NPT of South China published by Palaeogeography Palaeoclimatology Palaeoecology, Acta Palaeontologica Sinica, Precambrian Research and Bulletin of Geosciences (Zhu et al. Reference Zhu, Strauss and Shields2007; Zhu, Reference Zhu2010; Fatka et al. Reference Fatka, Steiner, Weber and Zhu2012; Shields-Zhou & Zhu, Reference Shields-Zhou and Zhu2013; Maletz et al. Reference Maletz, Steiner, Weber and Zhu2014), this thematic issue of Geological Magazine presents the most recent advances in the field based on studies of material from China, mostly South China. These studies cover a range of aspects including chronostratigraphy, geochronology, palaeobiology and sedimentary geochemistry. We hope that these new contributions will help to deepen current understanding about the NPT of the Earth–life system. New insights and the significant details from these contributions are outlined below.
1. Advances in Ediacaran – early Cambrian chronostratigraphy and geochronology
The absence of a high-resolution Ediacaran – early Cambrian global chronostratigraphic framework was hitherto a major reason for controversies surrounding models of the evolution of life and environment during the NPT. Since biostratigraphy is of limited usefulness for intra-basinal and inter-basinal stratigraphic correlation between Ediacaran – lower Cambrian successions, owing mainly to low species diversity, taxonomic uncertainty, and the sporadic and endemic palaeogeographic distribution of the fossils, it is generally agreed that high-resolution Ediacaran – early Cambrian chronostratigraphy requires integration of chemo- and biostratigraphic data with precise age constraints. The well-exposed and fossiliferous Ediacaran successions in the Yangtze Gorges area and lower Cambrian successions in the eastern Yunnan area of South China have long shown great potential for establishing such a high-resolution global chronostratigraphy. In recent decades, a generalized Ediacaran – early Cambrian chronostratigraphic model has been established (Fig. 1). However, the implications of the model for intra-basinal and inter-basinal correlations are still problematic, owing largely to the absence of age constraints for some prominent carbon isotopic anomalies, as well as for the first and last appearances of potential biostratigraphic index fossils, thus limiting attempts to calibrate bio- and geo-events during this critical transition.
In this special issue, Zhou et al. (Reference Zhou, Li, Xiao, Lan, Ouyang, Guan and Chen2017: this issue) report a SIMS zircon U–Pb age of 609±5 Ma for a tuffaceous bed immediately above the upper phosphorite unit of the lower Ediacaran Doushantuo Formation in the northern Yichang area, South China. Integrated litho-, bio- and chemostratigraphic correlations suggest that this unit is correlative with the upper Doushantuo phosphorite unit in the Weng'an area of central Guizhou. Therefore, these new age constraints indicate that the Weng'an biota could be as old as 609±5 Ma.
Yang et al. (Reference Yang, Li, Zhu and Condon2017a : this issue) report intriguing SIMS zircon Pb–Pb ages of 553.6±2.7 Ma and 546.3±2.7 Ma from ash beds in the middle of the upper Ediacaran Dengying Formation of eastern Yunnan, which are comparable to the ID-TIMS zircon U–Pb age of 550.55±0.75 Ma from an ash bed at the base of the Dengying Formation in the Yangtze Gorges area. These new dates imply that the base of the Dengying Formation is diachronous, thus requiring further work to resolve the complex transition between the Doushantuo and Dengying formations in different facies over the Yangtze Platform. The new dates from the middle Dengying Formation also suggest that the Shibantan Ediacara fossil assemblage is younger than the White Sea Assemblage and temporally overlaps with the Nama Assemblage.
Since ratification of the Cambrian GSSP in Newfoundland, defining the base of the Cambrian System on most continents outside the stratotype section has been a challenging task because of facies controls on occurrences of the index trace fossil Treptichnus pedum, as well as the absence of age or isotope constraints at the stratotype section. Therefore, other chronostratigraphic markers are badly needed. The Asteridium–Heliosphaeridium–Comasphaeridium (AHC) acritarch assemblage was considered as a potential biostratigraphic marker for the boundary interval. To test its validity, Ahn & Zhu (Reference Ahn and Zhu2017: this issue) present new data on the stratigraphic range of the AHC acritarch assemblage, and its spatial and temporal relationships to other bio- and chemostratigraphic information from the Yanjiahe Formation, Yangtze Gorges area, South China. The result indicates that the AHC assemblage correlates temporally with the Anabarites trisulcatus–Protohertzina anabarica (small shelly fossils) Assemblage Zone, and correlates spatially with the large negative carbon isotope anomaly at the basal Cambrian (BACE). This suggests that the AHC acritarch assemblage can be used as an additional and important chronological reference for global correlation of the Cambrian base.
Whereas the Ediacaran–Cambrian transition witnessed the rapid diversification of the Cambrian evolutionary fauna, the Cambrian–Ordovician transition records the further radiation of the Cambrian evolutionary fauna into the Palaeozoic evolutionary fauna. In order to accomplish a detailed chronostratigraphy for the Cambrian–Ordovician transition, Li et al. (Reference Li, Zhang, Chen, Zhang, Chen, Huang, Peng and Shen2017: this issue) report a high-resolution C-isotopic profile of the Wa'ergang section, western Hunan, South China. The Wa'ergang section has been proposed as a potential GSSP for the base of Cambrian Stage 10, the uppermost stage of the Cambrian System, and so these new data provide a chemostratigraphic reference point for global correlation of the Cambrian–Ordovician transition. Additionally, Guo et al. (Reference Guo, Deng, Hu and Wang2017: this issue) report high-resolution δ13Ccarb data for Ediacaran–Cambrian successions in the Keping area of the Tarim Basin, testing the validity of the chemostratigraphy for inter-basinal correlations.
2. New palaeobiological information about the early evolution of animals
The upper Ediacaran Dengying Formation of South China is characterized by a thick succession of carbonate rocks, predominantly dolostone, representing deposition in highly alkaline, seemingly barren shallow seas. However, abundant typical Ediacara fossils (the Shibantan biota) have recently been discovered in the dark grey limestone interval of the middle Dengying Formation in the Yangtze Gorges area. Unlike their preservation usually in siliciclastic rocks, the Shibantan fossils preserved in limestone provide fresh insights into the anatomy and the mode of life of Ediacara fossils. In this issue, Shen et al. (Reference Shen, Xiao, Zhou, Dong, Chang and Chen2017: this issue) describe a new Ediacara fossil, Curviacus ediacaranus gen. et sp. nov., from the Shibantan biota. C. ediacaranus exhibits a modular body construction consisting of serially arranged and crescent-shaped chambers, which is broadly similar to the co-occurring Yangtziramulus zhangii and other Ediacara modular fossils, such as Palaeopascichnus, providing new information about the diversity of the late Ediacaran modular organisms.
Chambered structures are also very common in Dengying dolostone and are similar to older, Tonian-age structures that were proposed to be fossils of sponges or sponge-grade organisms. Through detailed petrographic analyses and 3-D representations of these chambered structures within the Dengying dolostone, Luo, Pan & Reitner (Reference Luo, Pan and Reitner2017: this issue) argue that the structures result from microbial activities. This interpretation is consistent with the widespread occurrence in the Dengying dolostones of microbiolites and stromatolites, reminiscent of the microbe-dominated ecosystems of the Precambrian. The distinct microbially induced sedimentary structures (MISS) reported by Liu & Zhang (Reference Liu and Zhang2017: this issue) from the shelly fossil-bearing phosphorite of the Zhujiaqing Formation in eastern Yunnan suggest that this microbially dominated ecosystem may have lasted into earliest Cambrian time, persisting until the arrival of metazoan-dominated ecosystems after 520 Ma. Furthermore, a continuous evolutionary transition from late Precambrian to Cambrian times is supported by a multi-layered tubular microfossil Feiyanella manica gen. et sp. nov. from the lowest Cambrian Kuanchuanpu Formation, southern Shaanxi, South China. As reported by Han et al. (Reference Han, Cai, Schiffbauer, Hua, Wang, Yang, Uesugi, Komiya and Sun2017: this issue), F. manica shows profound similarities in morphology and reproduction to the characteristically late Ediacaran cloudinids (Cloudina and Conotubus).
During the past decades, more than 20 metazoan phyla or clades including primitive vertebrates have been discovered from the Cambrian fossil Lagerstätten of South China, particularly from the Chengjiang, Guanshan and Kaili faunas, providing invaluable information to improve our understanding of the early evolution of animals. Ongoing excavations continually reveal new fossil material discoveries. Three articles in this special issue report such new material. First, Zeng et al. (Reference Zeng, Zhao, Yin and Zhu2017: this issue) describe extraordinarily well-preserved appendages of a trilobite species, Hongshiyanaspis yiliangensis (Zhang & Lin in Zhang et al. Reference Zhang, Lu, Zhu, Qian, Lin, Zhou, Zhang and Yuan1980), from the Xiazhuang fossil assemblage within the Hongjingshao Formation (Cambrian Stage 3) near Kunming, Yunnan. This is the seventh trilobite species with complete appendages to be described and reconstructed in the world, and only the second one in China after Eoredlichia intermedia from the Chengjiang Lagerstätte. The new data support the mandibulate affinities of trilobites. Another new report is of a crown group fossil priapulid worm, Xiaoheiqingella sp., from the Cambrian Guanshan Lagerstätte (Cambrian Stage 4) near Kunming, Yunnan (Hu et al. Reference Hu, Zhu, Zhao and Steiner2017: this issue). The new discovery represents the third occurrence of the crown group fossil priapulids after the Cambrian Chengjiang Lagerstätte and the Pennsylvanian Mazon Creek Lagerstätte. The third new fossil report is by Yang et al. (Reference Yang, Zhao, Babcock and Peng2017b : this issue), who describe a new vauxiid sponge (Vauxiidae, Verongida), Angulosuspongia sinensis gen. et sp. nov., from the Kaili Formation (Cambrian Stage 5), Jianhe, Guizhou. This is the first discovery of vauxiid fossils reported from outside Laurentia, providing additional information towards a better understanding of early demosponge evolution.
3. Geochemical constraints on the evolution of redox conditions and seawater chemistry
Increasing evidence suggests that the rise of animals and their subsequent rapid diversification were directly linked to increased oxygenation of the atmosphere and ocean during the NPT (Och & Shields-Zhou, Reference Och and Shields-Zhou2012). However, the exact timing and mechanism of this oxygenation event, and how it links to the early evolution of animals, remain elusive. Recent multi-proxy geochemical studies from South China have indicated a stepwise increase in oxygenation during Ediacaran and early Cambrian times, and that modern-like oxygen levels were reached for the first time in Earth history at ~ 521 Ma, coinciding with the radiation of animals seen in fossil records from South China (Chen et al. Reference Chen, Ling, Vance, Shields-Zhou, Zhu, Poulton, Och, Jiang, Li, Cremonese and Archer2015). Meanwhile, other studies have proposed a more dynamic history of redox evolution during Ediacaran and early Cambrian times, suggesting that spatio-temporal heterogeneity of redox conditions may have exerted a strong impact on the early evolution of animals (e.g. Jin et al. Reference Jin, Li, Algeo, Planavsky, Cui, Yang, Zhao, Zhang and Xie2016; Och et al. Reference Och, Cremonese, Shields-Zhou, Poulton, Struck, Ling, Li, Chen, Manning, Thirlwall and Strauss2016; Sahoo et al. Reference Sahoo, Planavsky, Jiang, Kendall, Owens, Wang, Shi, Anbar and Lyons2016). Furthermore, statistical analyses of global iron-speciation data suggest only limited oxygenation through the Ediacaran and Cambrian periods (Sperling et al. Reference Sperling, Wolock, Morgan, Gill, Marcus, Halverson, Macdonald, Knoll and Johnston2015). These inconsistent results may have been caused by discrepancies among the different geochemical proxies, spatiotemporal heterogeneity of depositional environments and poor stratigraphic constraints on some of the studied successions, thus requiring more surgical investigations.
In this issue, Wei et al. (Reference Wei, Ling, Li, Wei, Wang, Chen, Zhu, Zhang and Yan2017: this issue) focus on redox evolution through a lower Cambrian succession (northwestern Guizhou), which was deposited on the shelf margin of the Yangtze Platform. Through analyses of nitrogen and sulphur isotopes in combination with trace elements (Mo and U) based on the drill core material, they conclude that seawater along the Yangtze shelf margin was: (1) anoxic and ferruginous with low sulphate concentration (relatively deep chemocline) during the Cambrian Fortunian and early Age 2; (2) anoxic and sulphidic (shallow chemocline and anoxic photic zone) during the late Cambrian Age 2 and early Age 3; and (3) anoxic and ferruginous during the middle and late Cambrian Age 3. The results also suggest that redox evolution on the Yangtze shelf margin was probably controlled by changing biological activity through feedbacks on the local marine environment.
In contrast to the lower Cambrian succession in the outer shelf environment, Jin et al. (Reference Jin, Li, Algeo, Cheng, Lei, Zhang and Shi2017: this issue) focused their studies on a lower Cambrian succession at the Wangjiaping section (near Yichang, western Hubei) from an inner shelf environment (in the paper, the authors consider it to represent an outer shelf setting). The high-resolution Fe–C–S–Al–Ti geochemical data document a redox transition from dominantly euxinic to ferruginous conditions with locally high sulphate availability during Cambrian Stage 3. Furthermore, relatively lower δ34Spy and lower total organic carbon (TOC) values, and higher positive correlation between TOC and Fepy/FeHR than deeper water successions reported previously suggested that euxinic conditions at Wangjiaping were in response to increasing marine productivity and organic matter-sinking fluxes. These data further indicate that spatial heterogeneity of redox conditions in the Yangtze shelves played an important role in the distribution of early animals.
In addition to redox evolution, Feng et al. (Reference Feng, Huang, Lu and Zhang2017: this issue) present major element data for the Fulu iron formation (IF) within Neoproterozoic glacial successions of South China. The data suggest that the iron source for the Fulu IF comprised a binary mixture between a hydrothermal source and detrital loads, although high-temperature hydrothermal input may also have contributed some iron to the IF. The data also indicate that the Fulu IF precipitated across an iron chemocline that separated a weakly oxic surface ocean from an oxygen-depleted deep ocean during the Neoproterozoic glaciation. Furthermore, the paper by Zhu & Jiang (Reference Zhu and Jiang2017: this issue) reports rare earth element (REE) data for the Ediacaran Doushantuo phosphorite, which contains the well-preserved phosphatized microfossils known as the Weng'an biota at Weng'an, central Guizhou. The results indicate a complex diagenetic history of the phosphatic grains, and suggest that the phosphorite formed under oxic bottom-water conditions with the involvement of hydrothermal fluids, providing new insight into the Ediacaran phosphorites and phosphatic fossils.
Acknowledgements
Papers in this special issue are mostly by members of the National 973 programme “From snowball Earth to the Cambrian explosion: Coevolution of life and environment around 600 myr ago” funded by the Ministry of Science and Technology of China (grant no. 2013CB835000). We particularly wish to thank the all the reviewers and our colleagues, without whose help this special issue would not be possible.

