


Introduction
The Spoon-billed Sandpiper Calidris pygmeus is a ‘Critically Endangered’ species with a population now estimated at less than 120 pairs (BirdLife International 2015, Syroechkovskiy et al. in prep.). Its breeding range is restricted to Chukotka and Koryakya in the far north-east of Russia, where it nests along a 4,500-km discontinuous narrow strip of coastal tundra (Tomkovich et al. Reference Tomkovich, Syroechkovskiy, Lappo and Zöckler2002, Zöckler and Lappo Reference Zöckler and Lappo2008, Zöckler et al. Reference Zöckler, Syroechkovskiy and Atkinson2010a). From here, it migrates down the eastern coast of Asia, through the Russian Far East, Korean Peninsula and along the Yellow Sea coast of China, before wintering along the coasts of Bangladesh, Myanmar, the Inner Gulf of Thailand, Vietnam, south and east China, with occasional records from India, Sri Lanka and Malaysia (BirdLife International 2001, Chowdhury Reference Chowdhury2010, Zöckler et al. Reference Zöckler, Syroechkovskiy and Bunting2008, Reference Zöckler, Htin Hla, Clark, Syroechkovskiy, Yakushev, Daengphayon and Robinson2010b).
A major decline in the breeding population of Spoon-billed Sandpipers was detected between 1977 and 2000 (Tomkovich et al. Reference Tomkovich, Syroechkovskiy, Lappo and Zöckler2002) and has continued, with the rate of decline estimated at 25% per annum (Zöckler et al. Reference Zöckler, Syroechkovskiy and Atkinson2010a). Surveys indicate that although breeding pairs successfully fledge young, recruitment to the known breeding populations is low (Zöckler et al. Reference Zöckler, Syroechkovskiy and Atkinson2010a). There is evidence that first year birds spend their first full summer in the non-breeding grounds (Tomkovich Reference Tomkovich1995, Zöckler et al. Reference Zöckler, Syroechkovskiy and Atkinson2010a, Eiam-Ampai et al. Reference Eiam-Ampai, Nimnuan, Sonsa, Sutibut and Round2011). Birds are apparently recruited to breeding populations near the areas in which they fledged. Low return rates of young birds to the breeding grounds suggest they suffer high mortality at this stage, or on passage (Zöckler et al. Reference Zöckler, Syroechkovskiy and Atkinson2010a). However, observations indicate juveniles can in some rare incidences disperse from their birth sites (Zhang Lin and P. Tomkovich in litt.). Low recruitment of birds could explain part or all of the observed decline in the species. Although not studied in detail, it has been suggested that adult mortality is also most likely to occur on the non-breeding areas, both on wintering and stop-over areas (Tong et al. Reference Tong, Lin, Li, Zöckler and Clark2012).
A major threat on non-breeding areas appears to be to be hunting or trapping. Trapping for larger waders on wintering areas has been found to be prevalent, with Spoon-billed Sandpipers being caught or poisoned as by-catch (Zöckler et al. Reference Zöckler, Htin Hla, Clark, Syroechkovskiy, Yakushev, Daengphayon and Robinson2010b, Chowdhury et al. Reference Chowdhury, Foysal, Das, Mohsanin, Diyan and Alam2011, Tong et al. Reference Tong, Lin, Li, Zöckler and Clark2012, Martinez and Lewthwaite Reference Martinez and Lewthwaite2013). In the Gulf of Mottama, Myanmar, at least 53 hunters were active in winter and 20 hunters on the east coast claimed to have captured over 150 Spoon-billed Sandpipers between 2000 and 2010 (Zöckler et al. Reference Zöckler, Htin Hla, Clark, Syroechkovskiy, Yakushev, Daengphayon and Robinson2010b). On Sonadia Island, Bangladesh, at least 25 hunters were active in winter and eight hunters claimed to have captured 22 Spoon-billed Sandpipers between October 2009 and April 2010 (Chowdhury Reference Chowdhury2010). In China, hunting is ongoing and widespread. Martinez and Lewthwaite (Reference Martinez and Lewthwaite2013) reported 460 mist nets between Zhuhai and Leizhou at mudflats in Guangdong Province. The prevalence of hunting along the remainder of the coast is unknown, but it has certainly been observed at a number of other sites. In addition to hunting, shorebirds and other coastal waterbirds are sometimes killed by poisoned bait, as has been observed in the Gulf of Mottama (Zöckler et al. Reference Zöckler, Htin Hla, Clark, Syroechkovskiy, Yakushev, Daengphayon and Robinson2010b), but also at sites in Jiangsu Province (Tong et al. Reference Tong, Lin, Li, Zöckler and Clark2012). Hunting mitigation measures and engagement with communities have been initiated at some sites, with some apparent early success (Zöckler et al Reference Zöckler, Htin Hla, Clark, Syroechkovskiy, Yakushev, Daengphayon and Robinson2010b). Other potential or actual threats include habitat loss (mostly to coastal development) and increasing levels of pollution in non-breeding areas. Coastal development and reclamation of mudflats are taking place across the passage and wintering range, with analysis of satellite images showing widespread loss of intertidal habitat since the early 1980s (Ma et al. Reference Ma, Melville, Liu, Chen, Yang, Ren, Zhang, Piersma and Li2014, MacKinnon et al. Reference MacKinnon, Verkuil and Murray2012, Murray et al. Reference Murray, Clemens, Phinn, Possingham and Fuller2014).
In order to target advocacy and protection efforts at conserving important habitats in important sites and reducing winter hunting pressures, a good knowledge of the areas used by non-breeding birds is required. To meet this need, between 2005 and 2013, field surveys in six countries were undertaken to improve knowledge of the non-breeding distribution of the Spoon-billed Sandpiper. While some sites are now well known, the species has such an extensive and poorly delineated non-breeding range that it is possible that important wintering sites remain undiscovered, hindering conservation efforts (Zöckler et al. Reference Zöckler, Htin Hla, Clark, Syroechkovskiy, Yakushev, Daengphayon and Robinson2010b).
It is impractical to survey the entire coastline in the species’ wintering range. However, a species distribution model can be used to model the suitability of the coastline and identify potential wintering sites, allowing field surveys to be better targeted. Species distribution models (SDMs; Guisan and Zimmermann Reference Guisan and Zimmermann2000) have proved to be highly useful in predicting new sites for poorly known species (e.g. Donald et al. Reference Donald, Buchanan, Collar, Abebe, Gabremichael, Mwangi, Ndang’ang’a, Spottiswoode and Wondafrash2010, Le Lay et al. Reference Le Lay, Engler, Franc and Guisan2010, Drew et al. Reference Drew, Wiersma and Huettmann2011) and in making assessments for conservation management (e.g. Rödder et al. Reference Rödder, Engler, Bonke, Weinsheimer and Pertel2010, Cushman and Huettmann Reference Cushman and Huettmann2010). Comparing environmental conditions at sites where Spoon-billed Sandpipers have been recorded with conditions across the potential wintering range might enable us to detect areas that could support unknown wintering populations of the species.
Here, we describe recent surveys that have updated our knowledge of the winter distribution (November–February) of the Spoon-billed Sandpiper, describing counts and documenting observations on the species’ ecology. These were combined with previous records of the species in a database, with coordinates where available (Bunting and Zöckler Reference Bunting, Zöckler, Boer, Galbraith and Stroud2006). We then used species distribution models, trained using the results of field surveys, with the aim of identifying unrecognised wintering areas, thereby helping to efficiently target conservation resources to the most important areas for the species. We provide a spatially geo-referenced GIS file of the predicted potential distribution to help target survey effort.
Methods
Field surveys
Field surveys were undertaken in six countries along the East Asian-Australasian flyway between 2005 and 2013. In January 2005, potential wintering sites in India were surveyed, followed by Bangladesh (January 2006), Thailand (December 2006) and Myanmar (six surveys between January 2008 and February 2013). Surveys also took place in Vietnam (Red River Delta in 2009 and the Mekong Delta in 2011). Since 2009, key sites on the southern Chinese coast have been surveyed. Regular winter surveys in Bangladesh began in October 2009 under the Bangladesh Spoon-billed Sandpiper Conservation Project (e.g. Bird et al. Reference Bird, Lees, Chowdhury, Martin and Haque2010, Chowdhury et al. Reference Chowdhury, Foysal, Das, Mohsanin, Diyan and Alam2011).
Areas for field surveys were selected using two sources of information. First, we collated records dating back to the 19th century (BirdLife International 2001) with more recent records supplied by visiting birdwatchers and local ornithologists. Second, visual assessment of Landsat Geocover Mosaic data (MDA Federal 2004), satellite images and aerial photos on Google Earth were used to identify areas of potentially suitable estuaries with large exposed mudflats and connected sandbars with little or no mangroves.
At each site, teams of surveyors looked for suitable habitats or flocks of small waders (calidrids and charadriids), whether feeding or roosting. Local knowledge collected from fishermen was often used to identify sites of shorebird concentrations, particularly temporary sand/mud flats offshore. Variation in the nature of sites meant that methods could not be standardised between sites, and there was variation in the amount of ground surveyed and number of surveys undertaken. Ideally, areas were surveyed at high tide when roosting birds could be carefully checked using a telescope. Training was given to surveyors to help them identify Spoon-billed Sandpipers. Surveys were mostly undertaken from boats with shallow draft that were able to rest level on intertidal substrates for long periods during low tide, providing observers with a stable platform from which to survey intertidal areas. An absence of boats in some areas prevented surveys from being undertaken.
In the Gulf of Mottoma, Myanmar, teams of surveyors fanned out across the tidal mudflats, when the tide receded. Each wader flock encountered was scanned with telescopes and the number of each species counted. Attention was paid to the behaviour of birds, especially feeding activities. Observations on prey items were recorded from many wintering sites, through direct observations using telescopes and based on photographs of feeding birds. Several scans could be made in the same area if multiple flocks were present. Surveyors also estimated the total number of small waders (calidrids and charadriids) seen during each survey. The proportion of Spoon billed Sandpipers was estimated by fitting a generalised linear model with binomial errors (Zöckler et al. Reference Zöckler, Htin Hla, Clark, Syroechkovskiy, Yakushev, Daengphayon and Robinson2010b) to the flock scan data. The total number of Spoon-billed Sandpipers was estimated by multiplying the average proportion of Spoon-billed Sandpipers observed by an estimate of the total number of small waders on the mudflats (Zöckler et al. Reference Zöckler, Htin Hla, Clark, Syroechkovskiy, Yakushev, Daengphayon and Robinson2010b).
Records from these field surveys were compiled into a central database, together with ad hoc records documented in literature or sightings which authors were aware of, based on their own knowledge or connections (BirdLife International 2001, Bunting and Zöckler Reference Bunting, Zöckler, Boer, Galbraith and Stroud2006). Information on site, date, coordinates and count were compiled where available. Additionally, surveyors assessed current or potential threats to sites, identifying whether there was evidence of hunting, development, or ongoing habitat change. Proposed infrastructure developments on sites were identified from local sources and documented plans.
Species distribution modelling
We modelled the potential winter distribution of Spoon-billed Sandpipers using geo-referenced records from the database from the period 2000 to 2013. These were largely observations collected during the field surveys, but also contained ad hoc sightings. Records prior to 2000 were not considered due to the potential for sites to have changed considerably over time. We took presence in one year as a presence, meaning no account was taken on the number of years in which a bird was present at a site. Accurate records of which sites have been visited in each year are not available (observers are generally more likely to report presences). Models were built using maximum entropy implemented in Maxent (Phillips et al. Reference Phillips, Anderson and Schapire2006) to link records to environmental variables. Maxent uses a machine learning technique to estimate the predicted distribution (likelihood of occupancy between 0 and 1) that is most evenly spread out across the study area, given constraints set by the training data. It has been shown to perform well with low sample sizes (Franklin Reference Franklin2009).
Models were run at 1-km resolution and the modelled area was a 0.1° strip along the coastline of South-East Asia. Coastline was derived from the bounds of values of the SPOT VGT data on which the models were based (see below), with the coast being defined as where reflectance values occurred next to No Data values. An area within a bounding box of 86.6° to 111.7° E and 5.0° to 23.3° N (dotted line in Figure 1) was used for the models. The area selected for the model covered the majority of wintering sites that were in close proximity to each other (assessed visually). This was done to balance between capturing the majority of wintering records, while minimising the variation in environmental conditions. It resulted in the exclusion of some areas that were remote from the majority of other records (e.g. Wenzhou and Min Jiang in China). However, the inclusion of all wintering areas would potentially reduce the efficacy of any final models. Models used species records and environmental data from November to February. Of 414 Spoon-billed Sandpiper records, just 85 1-km squares were occupied, with many cases of multiple records from locations. Environmental conditions at these 85 cells were compared to those at 10,000 pseudo-absences.

Figure 1. Winter distribution of Spoon-billed Sandpipers, based on records from 2005 to 2013. Size of dot indicates maximum number of birds seen in any single winter (Nov–Feb) in each 0.25 degree grid square. The dotted line depicts the area used for modelling (see Figure 2).
Environmental data summarised climate, land cover, tidal and offshore conditions. Our variable selection was based on measures which were recorded in a consistent way across the entire extensive study area. We selected readily available variables that could be measured in an objective way, across the entire study area now and in the future. Two bioclimatic variables from Worldclim (http://www.worldclim.org), mean temperature of the coldest quarter and precipitation of the coldest quarter, were included. Land cover was described from SPOT VGT data (http://www.vito-eodata.be/), with each of five bands (B0, B2, B3, MIR and NDVI) used. B0 measures reflectance between 0.43 - 0.47 µm (blue light), B1 measures reflectance between 0.61 - 0.68 µm (red light), B2 measures 0.78 - 0.89µm (near infra-red) and B3 measures 1.58 - 1.75 µm (mid or short wave infra-red). NDVI (Normalised Difference Vegetation Index) is derived from B3 and MIR, and is a measure of photosynthetic activity (http://www.vgt.vito.be/). For each band we extracted the maximal value in a middle 10-day period of each month for November 2008-February 2009, November 2009-February 2010 and November 2010-February 2011, and averaged these to produce a single value per band. Tide height (the average difference between high and low water) was extracted from topex calculations of tidal range (Ray et al. 1999). Offshore conditions were described by mean monthly average chlorophyll a concentration from Aqua MODIS ocean colour data at 4-km resolution (http://oceancolor.gsfc.nasa.gov/cgi/l3). Values were extrapolated to the adjacent land using Euclidean allocation. Distance to coastline (as defined above, and always positive as SPOT data were not available from areas on the seaward side of the coast) was also included as a variable.
This gave a total of 10 variables (two climatic, five land cover, ocean chlorophyll, tide height and distance to coast) in the initial model. Models were built by sequentially removing the variables that contributed the least to each model, based on percentage contribution values of < 1%. The regularisation multiplier was altered as appropriate to produce smooth response functions to reduce over-fitting of the models. Once a final model had been obtained, 10 replicates were run for cross-validation. Model fit was assessed by ROC/AUC statistics (area under the receiver operating characteristic curve). The AUC is a continuous threshold independent statistic for which a value of 0.5 indicates random discrimination between presences and absences, and values of 1 indicating perfect discrimination. An imbalance in the number of absences compared to presences can inflate AUC values (Lobo et al. Reference Lobo, Jiménez-Valverde and Real2008), but a value ≥ 0.9 indicates a good model.
Results
Field surveys
Based on records from the 2005–2013 surveys, we estimate a minimum total of 242 and a maximum of 378 Spoon-billed Sandpipers across the entire winter distribution (Table 1 and Figure 1). This figure is based on absolute counts in some areas and extrapolations (using the methods described above) in other sites that were too extensive to survey in their entirety (Table 1). Totals for Myanmar and Bangladesh together account for up to 80% of the wintering Spoon-billed Sandpiper population (Table 1, Figure 1). Notable was the upper part of the Gulf of Mottama in Myanmar where the total population estimate based on flock counts in 2010 was 160–220, based on 191 sightings of Spoon-billed Sandpipers in scans of flocks totalling 33,562 small waders and a total of 35,000 small waders estimated to be in the area by observers. Absolute numbers wintering on the island of Nan Thar in the state of Rakhine (near the border with Bangladesh) fluctuated between 14 and 35 birds with 20–25 in recent years. The large Asian River deltas of the Ganges-Brahmaputra, Ayeyarwaddy, Red River and Mekong all held three or more birds on a regular basis. Sites in China, Vietnam, and Thailand all held relatively low numbers. A summary of all wintering sites is presented in Table 1 and Figure 1.
Table 1. Number of Spoon-billed Sandpipers observed at sites surveyed since 2005. Minimum and maximum counts from different years over the reporting period 2005–2013 except for * where total number of Spoon-billed Sandpiper in Gulf of Mottama has been extrapolated, based on flock counts in 2010–2012. Wintering population estimates are based on counts 2011–2013.

Species distribution models
The performance of the model was convincing according to the AUC of 0.946 ± 0.028. Neither of the climate variables contributed more than 1% to the models, so were dropped from the model along with mid infra-red reflectance (Table 2). Offshore chlorophyll was of particular importance, and examination of the relationship (Figure S1 in the online supplementary material) suggests that birds were more likely to occur in areas where concentrations of chlorophyll offshore were higher. Relationships with reflectance from SPOT are more difficult to interpret, but the negative association with NDVI (Figure S1) might simply reflect areas of extensive unvegetated land (i.e. estuarine mud) being more likely to be occupied than vegetated areas inland. Areas in the top 5% of values for the model included the areas along the coast of Bangladesh, western Myanmar, the outer regions of the Gulf of Mottoma in Myanmar and the Inner Gulf of Thailand (Figure 2). Most of the larger areas identified as suitable are already known to be used by wintering Spoon-billed Sandpipers, but there would appear to be less extensive but still potentially suitable areas south of the Mekong delta. Small parts of the coast of western Myanmar, eastern Bangladesh and the Guangxi and Guangdong regions of China may also merit further investigation. Figure S2, a geo-referenced .tif file for GIS, is provided to allow detailed interrogation of the areas predicted to be potentially occupied.
Table 2. Variables retained in the final model with their permutation importance values and percentage contributions.
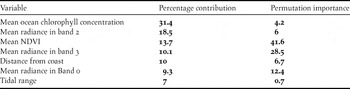
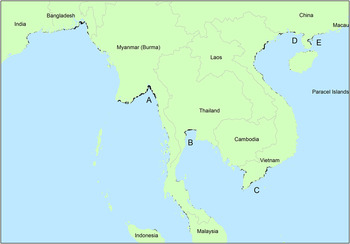
Figure 2. Modelled potential distribution of Spoon-billed Sandpiper habitat. Map shows the 5% of the study area with the highest predicted likelihood of occupancy. The predicted distribution of the Spoon-billed Sandpiper on which Figure 2 is based is available as a .tif for GIS in Figure S2. A indicates Gulf of Mottama, Myanmar, B is Inner Gulf of Thailand, C is south Mekong delta, D is Guangxi and E indicates Guangdong.
Discussion
The species is widely distributed in winter over an area stretching from Bangladesh to Eastern China just south of the Yangtze River estuary. The majority of birds seem to winter in Myanmar, where 60% of the population has been observed. At least 80% of the entire global population appears to winter in Myanmar and Bangladesh. Surveys of wintering Spoon-billed Sandpipers were undertaken at sites in six of the flyway countries over a nine-year period. Collating records of these birds allows us to build up a database of areas used by wintering individuals, pointing to areas of conservation priority for the species. Difficulties in standardisation of survey methodologies between and within sites, not least due to shifting sediments in estuaries, means that for many sites it is difficult to produce accurate population trends over multiple years. However, these surveys have allowed crude estimates of abundance to be calculated and broadly allow us to detect if populations continue to decline. From our surveys, the estimated population in the wintering areas surveyed is between 242 and 378 individuals. This is consistent with current estimates derived from the breeding area of fewer than 120 breeding pairs (Syroechkovskiy et al. in litt.).
Descriptive observations made during surveys and photos taken suggest that Spoon-billed Sandpipers fed on a broad variety of benthos, including small crabs, shrimps, polychaetes and amphipods, and often small flies. Birds were observed feeding in shallow waters or mudflats with puddles, but did not appear to visit or feed in the areas of deep and fine mud found in many shallow areas of estuaries.
Our species distribution model had a relatively high AUC value and correctly identified the majority of the known wintering areas as being highly suitable, suggesting that it provides a good description of the distribution of Spoon-billed Sandpipers. The model was built to assess the potential distribution of the species, rather than to interpret habitat associations, but the association with chlorophyll was notable. As we do not have geo-referenced data on where Spoon-billed Sandpipers do not occur, we were unable to undertake a thorough assessment of model performance and assess both specificity and sensitivity. The majority of suitable areas identified by our model were around sites at which birds are regularly recorded (Table 1), suggesting the main wintering areas are already known. This is supported by the scarcity of records away from major, known wintering grounds. However, since the species occurs in small numbers across extensive estuary systems, it may have been overlooked at some sites. For example, in 2013, just one individual each was reported from two sites in Malaysia (David Bakewell in litt.), one in the Eastern Ayeyarwaddy Delta in November 2013 (Moses and Zöckler Reference Moses and Zöckler2013), two in 2014 at the same site (Saw Moses pers. comm.) and one in Cambodia in January 2014 (Simon Machmut pers. comm.). Substantial populations might remain in China: in 2008, local bird watchers and photographers mentioned wintering birds in the Min Jiang estuary in Fujian Province. Up to 14 birds have since been recorded at this site in winter (Bai et al. Reference Bai2015), although none were seen in 2013. After intensive surveys in 2010 and 2011, further wintering sites were discovered west of Min Jiang (Fujian), in Beihai (Guangxi), on the island of Hainan and Fangchengang near the Vietnamese border. These mostly involved sightings of one or two individuals, but in Lei Zhou Peninsula towards Hainan (Guangdong), up to five were observed in December 2012 (Martinez and Lewthwaite Reference Martinez and Lewthwaite2013). Up to eight birds were recorded in 2013 in Wenzhou in Zhejiang Province (MT), making this both the most northerly and easterly known wintering site. The model highlighted a number of previously unsurveyed sites with small areas of potentially suitable habitat which may merit further investigation. These include parts of the coast of western Myanmar and eastern Bangladesh and the southern part of the Mekong delta. Potentially suitable areas were also indicated in the Guangxi and Guangdong regions, which coincide with areas identified as potentially suitable habitat from a visually interpretation of satellite images. Future field surveys should be targeted at these areas. Field surveys would ideally also be made at locations predicted to be unoccupied to test the veracity of the model. However, practicalities and expense means that the areas highlighted as potentially suitable should be prioritised.
Difficulties in survey methods, including the speed at which estuaries alter due to storms and the highly dynamic coastal tidal systems of some sites (authors’ pers. obs.), means we cannot make an accurate assessment of population changes within most sites. However, the indications of declining populations in the breeding areas (Zöckler et al. Reference Zöckler, Syroechkovskiy and Atkinson2010a) are supported by robust surveys. Apparent population declines have been noted across the wintering areas, although they appear to be less severe than those documented on the breeding grounds. For example, in Bangladesh the species was regularly recorded around Hatia Island in large numbers from the late 1980s to early 1990s, with the highest single count of 289 birds in 1989 from Moulevir Char (Howes and Parish Reference Howes and Parish1989). Smaller numbers (three to c.30) were then recorded between the late 1990s and early 2000s, with the westernmost record of four birds on 24 February 1992 from Egg Island on the edge of the Sundarbans (Thompson et al. Reference Thompson, Harvey, Johnson, Millin, Rashid, Scott, Stanford and Woolner1993, Thompson and Johnson Reference Thompson and Johnson2003). A survey on Egg Island and adjacent islands in January 2013 found no Spoon-billed Sandpipers (Chowdhury et al. Reference Chowdhury, Diyan, Zöckler, Foysal and Lemke2014). On Sonadia Island, regular surveys began in 2009 and records reflect a consistent build up during northward migration: counts of 13 in January and 25 in March 2010 were followed by 15 and 11 in January and 27 and 24 in March 2011 and 2012. Up to eight birds were recorded at the tip of Teknaf peninsula in 2006, two in 2010 but no birds have been recorded there since at least 2012 (Thompson et al. Reference Thompson, Chowdhury, Haque, Khan and Halder2014, Chowdhury et al. Reference Chowdhury, Diyan, Zöckler, Foysal and Lemke2014). Wintering numbers from Nan Thar in Myanmar declined from 35 in 2008 to only 14 in 2009 and 2010 but increased to 20 in 2013 (Zöckler et al. Reference Zöckler, Zaw Naing, Moses, Nou Soe and Htin Hla2014). In Vietnam, the Red River Delta was a regular wintering site with up to 17 birds recorded in the mid-1990s (Jonathan Eames pers. comm.), but no birds were found during a 10-day survey in January 2010 and only one bird in 2013, despite intensive searching. The ubiquity of the declines across all sites seems to indicate that the declines are genuine, rather than artefacts of variation in observer effort, especially as the species has had an increasing profile in recent years. Spoon-billed Sandpipers are surprisingly difficult to identify in flocks of waders especially when roosting, but the development of very high resolution telescopes and digital cameras for birdwatching has greatly increased the ability of general birdwatchers to identify the species with certainty. It has, however, required targeted surveys to find most of the new locations for the species. Chance findings of Spoon-billed Sandpipers cannot be relied upon to locate the remaining wintering areas and it must be assumed that they are not wintering in areas regularly visited by birdwatchers. Given the extremely difficult terrain in which most birds have been found, it is likely that the survey methods will need to be tailored for each site rather than there being a survey protocol that can be used across the wintering range.
Evidence of a link between changes in habitat and declines in Spoon-billed Sandpipers is lacking, not least because of the small number of detailed studies on the species. Habitat loss is a threat at a number of occupied sites (e.g. Chowdhury Reference Chowdhury2010, Chowdhury et al. Reference Chowdhury, Foysal, Das, Mohsanin, Diyan and Alam2011, Zöckler et al. Reference Zöckler, Zaw Naing, Moses, Nou Soe and Htin Hla2014), and reclamation of crucial intertidal areas on the passage have rendered them unsuitable for waders (Murray et al. Reference Murray, Clemens, Phinn, Possingham and Fuller2014). Therefore, sites which are currently occupied should be rigorously protected to prevent loss or alteration on a precautionary basis. The species distribution models did identify pockets of potentially suitable habitat that have not yet been surveyed. Targeting survey effort to these areas, whilst maintaining regular ongoing monitoring at other known sites, would be a priority. However, it would not appear that extensive areas of potentially suitable habitat still exist. Habitat loss on stop-over sites and wintering areas, together with hunting pressure (Zöckler et al. Reference Zöckler, Htin Hla, Clark, Syroechkovskiy, Yakushev, Daengphayon and Robinson2010b), might be the main driver for population declines in this species. It is also possible that some sites have become degraded in a way that is not easily detectable; e.g. the gradual reclamation by expanding shrimp farms, as observed in Thailand, Myanmar and Bangladesh (Zöckler et al. Reference Zöckler, Syroechkovskiy and Bunting2008), or that Spoon-billed Sandpiper distributions are driven by fine-scale factors in the environment or landscape that are not adequately captured by our model. Similarly, we cannot discount the role of habitat loss on passage sites and on the breeding grounds in causing declines, although loss on breeding areas might be less likely as a cause of decline, as breeding grounds have apparently largely remained unchanged and research from the breeding grounds (Zöckler et al. Reference Zöckler, Syroechkovskiy and Atkinson2010a) points to threats along the migration route. The species might also be vulnerable to a range of other, less obvious, threats at passage and wintering sites, such as pollution or the impact of agricultural run-off: topics which also merit research.
Protection through site designations is much desired but alone is insufficient, and needs to be enforced. Hunting and trapping of birds is widespread and observed in almost all major sites. The species urgently requires stronger or better enforced protection in China and Vietnam. In China, Spoon-billed Sandpipers are on the “list of terrestrial wildlife under state protection, which are beneficial or of important economic or scientific value” under the Law of the People’s Republic of China on the Protection of Wildlife, but they are not listed as State Key Protected species. Hunting appears to be an important factor in the declines of Spoon-billed Sandpipers in many winter sites. In the Gulf of Mottama, at least 53 hunters were active in winter, having caught annually an estimated 20,000 birds (Zöckler et al. Reference Zöckler, Htin Hla, Clark, Syroechkovskiy, Yakushev, Daengphayon and Robinson2010b). On the East Coast of the Gulf, 20 hunters caught a minimum of 150 Spoon-billed Sandpipers in total between 2001 and 2010 (BANCA 2011a) and 84 between 1989 and 2011 on the West coast of the Gulf (BANCA 2011b). On Sonadia Island, Bangladesh, at least 25 hunters were active in winter and eight hunters claimed to have captured a total of 22 Spoon-billed Sandpipers between October 2009 and April 2010 (Chowdhury Reference Chowdhury2010). This means actions will also need to be taken at the local or community scale, targeting damaging practices such as indiscriminate hunting and trapping of birds. In these areas, in addition to enforced legislative protection of sites, conservation efforts should be targeted towards reducing hunting pressure and supporting alternative livelihoods to the hunting of water birds for food. Mitigation and engagement with communities has been initiated at some sites, with early evidence of success (Zöckler et al. Reference Zöckler, Htin Hla, Clark, Syroechkovskiy, Yakushev, Daengphayon and Robinson2010b, BANCA 2011a). Expanding this action to other areas where Spoon-billed Sandpipers winter should be considered a priority for conservation, and our species distribution model could be used to guide survey efforts to identify any unknown populations. Law enforcement has been applied in South China, where hunting has been widespread (Martinez and Lewthwaite Reference Martinez and Lewthwaite2013).
Regular monitoring is recommended to closely watch the state of wintering populations. Over the past eight years, coordinated monitoring efforts have been carried out regularly at known sites with increasing efficiency, and searches for new areas focused on Myanmar (Zöckler et al. Reference Zöckler, Htin Hla, Clark, Syroechkovskiy, Yakushev, Daengphayon and Robinson2010b, Reference Zöckler, Zaw Naing, Moses, Nou Soe and Htin Hla2014), Bangladesh (Chowdhury et al. 2010, 2013, Chowdhury and Foysal Reference Chowdhury and Foysal2015), Thailand (Round Reference Round2006, Nimnuan and Daengpayon Reference Nimnuan and Daengpayon2008, Sripanomyom et al. Reference Sripanomyom, Round, Savini, Trisurat and Gale2011) and Vietnam (this study). These surveys have been based around January counts in an attempt to minimise movement of birds between sites. Although we do not yet know enough about the species’ movements, we observed little fluctuation in numbers of Spoon-billed Sandpipers and little movement between sites during this period.
A species distribution model described the known distribution of the species well, highlighting all major sites known to be used by wintering birds. A few new potential areas were identified in Bangladesh, China, Myanmar and Vietnam. Field surveys should be targeted at these locations urgently to determine if birds are present, and initiate conservation actions as required to mitigate any threats. The protection of sites that are found to be occupied should be encouraged, and is likely to benefit many other species in addition to Spoon-billed Sandpipers. Protection could also be applied to sites which have not yet been surveyed, based on the precautionary principle, especially if large numbers of shorebirds are known to occur there. This protection, whether legal or community based, should be extended to preventing hunting in addition to habitat loss.
Supplementary Material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0959270915000295
Acknowledgements
For help with surveys we would like to thank S. Balachandran, A. Nayak, G. Maheshwaran, M. Fanck, K. Webb, A. Sharma (in India); E. Ul Haque, A. Islam, Z. Islam, S. Islam, M. L. Khan, J. Schwahn, O. Grell, C. Bock, Paul M. Thompson, Md. Foysal, M. Abdullah Abu Diyan, Omar Shahadat, Nazim Uddin Prince, Sakib Ahmed (in Bangladesh); A. Moe, Go Pan, S. Moses, N. Lwin, L. Win, Z. Lunn, K. Eberhardt, G. Chisholm, R. Kelsey, A. Keaveney, S. Daengphayon, P. Round, H. Andretzke, N. Yakushev, E. Koblik, A. Bräunlich, T. Noah, S. Klasan, R. Robinson (in Myanmar); Nguen Bao (in Vietnam), and J. Martinez (in South China). We would like to thank Kei Dan Ren Foundation for Nature, Tokyo; the German Ornithological Society (DO-G); Manfred Hermsen Foundation in Bremen; Bird Life’s Preventing Extinction Programme and BirdLife–Asia; the Convention on Migratory Species (CMS); the BBC Wildlife Fund; British Trust for Ornithology (BTO); the Darwin Initiative UK; East Asian Australasian Flyway Partnership (EAAFP); Lighthouse Foundation in Hamburg; National Audubon; Ocean Park Conservation Foundation in Hong Kong; the Royal Society of Protection of Birds (RSPB) UK; IUCN Save our Species (SOS); US Fish and Wildlife Service, and several private donors, who all supported the surveys. We thank Jan Engler, Charlie Moores and Paul Donald for comments that greatly improved an earlier version of this manuscript.
This paper is dedicated to the late Dr Tony Htin Hla, who was passionate about the conservation of Spoon-billed Sandpipers in Myanmar.






