


Introduction
Despite a short colon and the lack of a functional caecum as evolutionary adaptations to a strict carnivorous diet, considerable microbial fermentation occurs in the hindgut of domestic cats( Reference Brosey, Hill and Scott 1 ). Microbiota of domestic cats are capable of fermenting a broad range of dietary plant fibres( Reference Brosey, Hill and Scott 1 – Reference Sunvold, Fahey and Merchen 4 ). An in vitro study on the fermentation of various plant fibre sources using faecal inoculum of different species and ruminal fluid of cattle showed that the feline faecal inoculum produced the highest concentrations of total SCFA and acetate, supporting the hypothesis of substantial fermentation activity of the feline colonic microbiota( Reference Sunvold, Hussein and Fahey 5 ). In addition, in vivo research demonstrated that concentrations of volatile fatty acids in the colon of healthy cats were comparable with those measured in the forestomach of ruminants and large intestines of other single-stomached mammals, while concentrations in the small intestine were higher than those reported in other species( Reference Brosey, Hill and Scott 1 ). The study, therefore, provides another indirect proof of the fermentation potential of the feline gut microbiota. As in other species, the supplementation of plant fibre to feline diets has been associated with beneficial effects on several disease conditions (for example, Elliott( Reference Elliott 6 ), Rutgers & Biourge( Reference Rutgers, Biourge, Pibot, Biourge and Elliott 7 ), Freiche et al. ( Reference Freiche, Houston and Weese 8 ) and Fischer et al. ( Reference Fischer, Kessler and de Sá 9 )) and on the general health of the gastrointestinal tract( Reference Buddington, Sunvold, Reinhart and Carey 10 – Reference Bueno, Cappel and Sunvold 12 ). The majority of the information on the benefit of dietary plant fibre is, however, extrapolated from human nutrition as described by Sunvold( Reference Sunvold, Carey, Norton and Bolser 13 ). Dietary and metabolic interspecies differences should be taken into account, and specified in vitro and in vivo studies have been undertaken in dogs and to a lesser extent in cats. A first overview of in vitro and in vivo studies on the effects of dietary plant fibre supplementation to canine and feline diets was done by Sunvold( Reference Sunvold, Carey, Norton and Bolser 13 ) and Buddington and Sunvold( Reference Buddington, Sunvold, Reinhart and Carey 10 ). Additionally, more recent studies in felines are described and discussed in the present review. Besides plant fibres, the importance of animal fibre for carnivorous species has been discussed by Depauw et al. ( Reference Depauw, Hesta and Whitehouse-Tedd 14 ) in cheetahs. Animal fibre is defined by the latter authors as low- to non-digestible (glyco)protein-rich substances that are potential substrates for large-intestinal fermentation( Reference Depauw, Hesta and Whitehouse-Tedd 14 ). Plantinga et al. ( Reference Plantinga, Bosch and Hendriks 15 ) hypothesised that the consumption of whole prey, which is a major source of animal fibre, by feral cats might enhance gut health and affect the microbiota differently as compared with foods of plant origin. In domestic cats, however, little research has been done on animal fibre, despite the fact that animal meal, which is a source of animal fibre, is the main protein source in the majority of the commercially available extruded and wet diets( Reference Yamka, Jamikorn and True 16 , Reference Dozier, Dale and Dove 17 ) and despite the growing popularity of feeding raw meat-based diets containing animal fibre to pets( Reference Kerr, Vester Boler and Morris 18 ). The results of the scarce in vivo research in domestic cats on the effects of animal fibre on nutrient digestibility and faecal fermentation endproduct concentrations are also described in the present review.
Bacterial populations in the gastrointestinal tract
Apparent quantitative and qualitative differences have been shown among bacterial populations present in the different anatomic regions of the gastrointestinal tract of cats( Reference Buddington, Sunvold, Reinhart and Carey 10 , Reference Ritchie, Steiner and Suchodolski 19 ). Qualitatively, the microbiota in different intestinal compartments tend to be more similar within the individual, than between corresponding compartments of different cats, suggesting a large inter-individual variation( Reference Ritchie, Steiner and Suchodolski 19 ). Quantitative data not only depend on host characteristics, such as age, gastrointestinal disease and environment, but also on the sample collection method, and the enumeration technique (culture plating v. molecular techniques). An overview of studies investigating the effects of these factors is given in Tables 1 and 2. For a review of different enumeration techniques, their advantages and disadvantages, see Suchodolski( Reference Suchodolski 20 ).
Table 1 Overview of effects of the host factors age, gastrointestinal (GI) disease and environment on quantitative data of the gastrointestinal microbiota in cats

conc., Concentrations; CFU, colony-forming unit; IBD, inflammatory bowel disease; ↓ , decrease.
* Mean over both diets (34 and 53 % DM crude protein) and different ages of kittens (8, 12 and 16 weeks)
† Median of seventeen IBD-inflicted and ten healthy cats, ranges are 0–4219 bacteria/mm2 for IBD-inflicted and 0–3 for healthy cats; values determined by fluorescence in situ hybridisation.
Table 2 Overview of the effects of sampling and enumeration techniques on quantitative data of the gastrointestinal microbiota in cats

FISH, fluorescence in situ hybridisation.
Microbiota in the different regions of the feline gastrointestinal tract
An overview of the microbiota in different regions of the gastrointestinal tract is given in Fig. 1 and has recently been reviewed by Minamoto et al. ( Reference Minamoto, Hooda and Swanson 21 ).
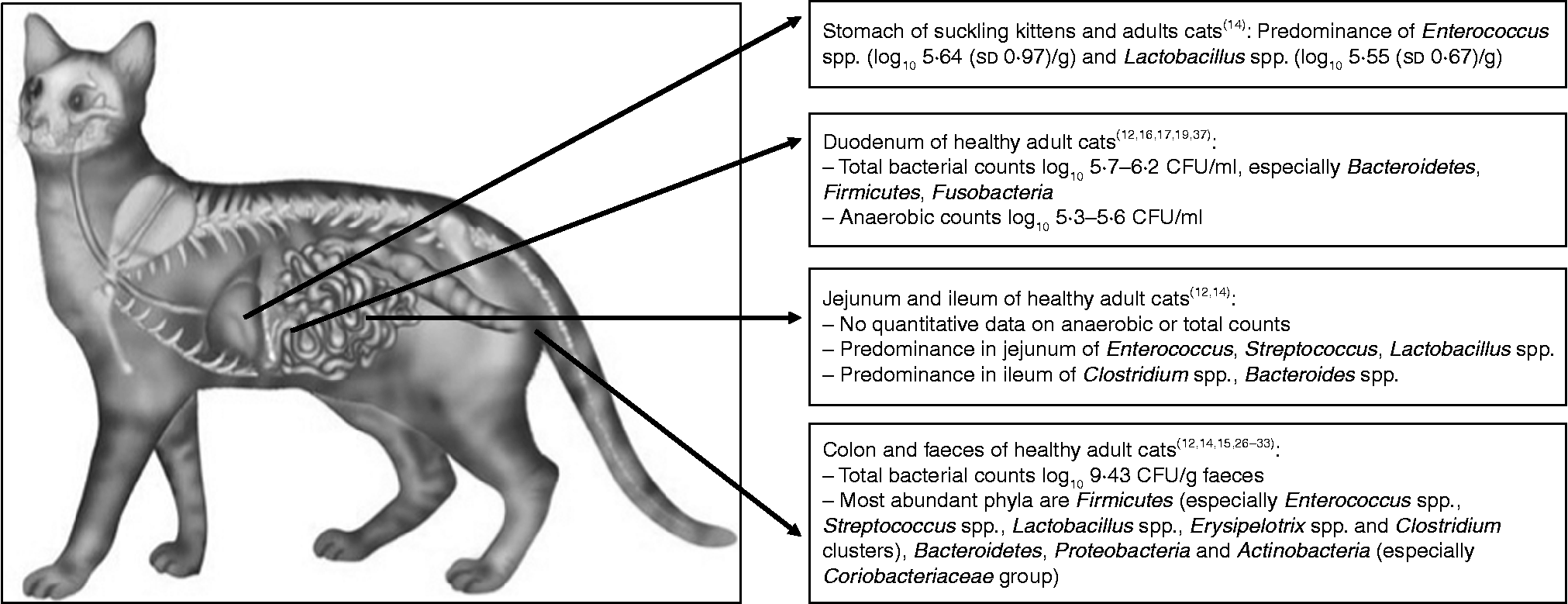
Fig. 1 Overview of the feline microbiota in different regions of the gastrointestinal tract. CFU, colony-forming unit.
Stomach
Microbiota in the stomach contents of suckling kittens and adult cats were studied by Osbaldiston & Stowe( Reference Osbaldiston and Stowe 22 ). The samples were collected after euthanasia, laparotomy and incision of the stomach wall, and culture-plating techniques were used( Reference Osbaldiston and Stowe 22 ). In this part of the gastrointestinal tract, the microbiota mainly constituted of enterococci (log10 5·64 (sd 0·97)). Likewise, all kittens and six out of nine adult cats showed similar counts of lactobacilli (log10 5·55 (sd 0·67)). Comparable counts of five other genera of anaerobic micro-organisms were isolated from the stomach contents of some cats (Streptococcus spp., Staphylococcus spp., Proteus spp., Bacillus spp., Pasteurella spp., Mima spp., Escherichia spp., Clostridium spp., Catenabacterium spp., Eubacterium spp., Bacteroides spp. and Veillonella spp.). No effects of diet were observed between adult cats on a standard feline diet (Hill's Prescription Diet Feline c/d; Hill's Pet Nutrition Inc.) and adult cats on a chemically defined, liquid elemental diet (amino acids, carbohydrates, fats, vitamins and minerals), both fed for 6 weeks( Reference Osbaldiston and Stowe 22 ). In humans, the microbiota of individuals on elemental diets decrease both in number and diversity, due to a lack of continuous supply of nutrients to the large intestine, since these diets are highly digestible( Reference Winitz, Adams and Sudman 23 ). In cats, the reason for the lack of diet effect was postulated to be the different passage rates through the gastrointestinal tract compared with humans.
The elemental diet thus provided a sufficient and continuous supply of nutrients necessary for bacterial survival and replication throughout the whole feline gastrointestinal tract on both diets( Reference Osbaldiston and Stowe 22 ). No information on the number of meals was available in the above-mentioned study. However, this factor could influence the study outcome significantly. No raw meat or ‘natural’ diets were fed to the cats in this study.
Small intestine microbiota: duodenum
The microbiota, determined by culture-plate techniques, in the proximal part of the small intestine have been found to show quantitative and qualitative changes over time, and a considerable variation among individual cats at the same time point, regardless of the sampling technique (direct needle aspiration after laparotomy v. endoscopy; diluted v. undiluted endoscopic samples)( Reference Sparkes, Papasouliotis and Sunvold 24 , Reference Papasouliotis, Sparkes and Werrett 25 ). Despite this variation, both studies found large numbers of bacteria in the proximal part of the small intestine in healthy cats, which is in accordance with the results of Johnston et al. ( Reference Johnston, Lamport and Batt 26 , Reference Johnston, Swift and Forster-Van Hijfte 27 ) and Smith( Reference Smith 28 ). Due to the use of diluted endoscopic samples (Table 2), lower numbers of bacteria were observed by Muir et al. ( Reference Muir, Gruffydd-Jones and Cripps 29 ).
Controversy has arisen about the reliability and the clinical significance of the absolute numbers of bacteria counted from small-intestinal fluid samples( Reference German, Day and Ruaux 30 ). In cats, considerably higher numbers of bacteria were found in the proximal small intestine in comparison with humans( Reference Finegold, Sutter, Mathisen and Hentges 31 ) and dogs( Reference Mentula, Harmoinen and Heikkilä 32 ), possibly as an adaptation to an obligate carnivorous diet( Reference Johnston, Swift and Forster-Van Hijfte 27 ). The causative factors in the carnivorous diet responsible for this high bacterial number in the proximal small intestine are yet to be unravelled. Furthermore, it is suggested that feline host defences to indigenous microbiota may be particularly well developed, and small-intestinal bacterial overgrowth is not a common clinical syndrome in cats with chronic non-obstructive gastrointestinal disease (Table 1)( Reference Johnston, Swift and Forster-Van Hijfte 27 ).
Qualitatively, the most abundant bacterial phylum in the feline duodenal microbiota was Firmicutes, consisting primarily of Clostridiales, detected with both culture-plating as well as molecular techniques( Reference Ritchie, Steiner and Suchodolski 19 , Reference Papasouliotis, Sparkes and Werrett 25 , Reference Johnston, Swift and Forster-Van Hijfte 27 ). The majority of the identified Clostridiales belonged to Clostridium clusters I and XIVa( Reference Ritchie, Steiner and Suchodolski 19 ), the latter of which is known to encompass beneficial butyric acid-producing species( Reference Collins, Lawson and Willems 33 ). Clostridium spp. were found in more than 50 % of the duodenal aspirates together with Enterococcus faecalis, Bacteroides spp., Pasteurella spp., Streptococcus spp., and unidentified Gram-negative aerobic rods( Reference Sparkes, Papasouliotis and Sunvold 24 ). In contrast, Johnston et al. ( Reference Johnston, Lamport and Batt 26 , Reference Johnston, Swift and Forster-Van Hijfte 27 ) found Bacteroides spp. to be the most abundant anaerobic bacteria accompanied by Eubacterium spp. and Fusobacterium spp. Again, Pasteurella spp. were found to be the most abundant aerobic bacteria in duodenal fluid( Reference Johnston, Lamport and Batt 26 ). Ritchie et al. ( Reference Ritchie, Steiner and Suchodolski 19 ) did not find Bacteroides spp. in the duodenum, which could be explained by the low number of cats in which duodenal samples were obtained. The differences in the recovery of Bacteroides among the above-mentioned studies is most probably not affected by diet or age differences, since in all studies commercial wet or dry diets were fed and only young cats with an age between 1 and 2 years were included. Effects of cat sex could be excluded, as Sparkes et al. ( Reference Sparkes, Papasouliotis and Sunvold 24 ) used both female and male cats, whereas Johnston et al. ( Reference Johnston, Lamport and Batt 26 ) used female cats only, and both studies recovered Bacteroides spp. as (one of) the major bacterial species. Body-weight differences between studies are assumed to be small, since in all studies healthy cats were included, so an effect of body weight on the recovery of Bacteroides spp. seems unlikely.
Small intestine microbiota: jejunum and ileum
In the feline jejunum, the predominant species were Enterococcus spp., Streptococcus spp. and Lactobacillus spp., detected with both culture-plating as well as molecular techniques( Reference Ritchie, Steiner and Suchodolski 19 , Reference Osbaldiston and Stowe 22 ). In addition, Osbaldiston & Stowe( Reference Osbaldiston and Stowe 22 ) observed higher frequencies of the latter two species and coliforms in the jejunum as compared with the stomach. In comparison with the microbiota of the stomach, jejunal microbiota showed the presence of three other bacterial species (Micrococcus spp., Klebsiella spp., Enterobacter spp.) and, in one cat, yeasts were cultured from jejunal contents. Again, no differences due to diet were found( Reference Osbaldiston and Stowe 22 ). For more details on the diet types, see the ‘Stomach’ section.
The ileal microbiota consisted mainly of Clostridium spp. and Bacteroides spp.( Reference Ritchie, Steiner and Suchodolski 19 ). For the Clostridiales class, the predominant clusters were the same as in the duodenum( Reference Ritchie, Steiner and Suchodolski 19 ). Papasouliotis et al. ( Reference Papasouliotis, Sparkes and Gruffydd-Jones 34 ) observed significantly higher numbers of bacteria, particularly aerobes, in the ileum compared with the duodenum after culturing intestinal juice, simultaneously aspirated at the two sites. As in the study of Johnston et al. ( Reference Johnston, Lamport and Batt 26 ), Pasteurella spp. were the predominating aerobe species in the duodenum, whereas in the ileum, enterococci and Escherichia coli were the most common aerobes( Reference Papasouliotis, Sparkes and Gruffydd-Jones 34 , Reference Gruffyd-Jones, Papasouliotis, Sparkes, Reinhart and Carey 35 ). As in Ritchie et al. ( Reference Ritchie, Steiner and Suchodolski 19 ), the predominating anaerobes in both segments were Clostridium spp., especially clusters I and XIVa( Reference Papasouliotis, Sparkes and Gruffydd-Jones 34 , Reference Gruffyd-Jones, Papasouliotis, Sparkes, Reinhart and Carey 35 ).
Large intestine and faeces
In the colon and faeces of domestic cats, the most abundant phylum was Firmicutes, which was dominated by Enterococcus spp.( Reference Ritchie, Steiner and Suchodolski 19 , Reference Osbaldiston and Stowe 22 , Reference Desai, Musil and Carr 36 ), Streptococcus spp.( Reference Osbaldiston and Stowe 22 ), Lactobacillus spp.( Reference Ritchie, Steiner and Suchodolski 19 , Reference Osbaldiston and Stowe 22 , Reference Desai, Musil and Carr 36 , Reference Garcia-Mazcorro, Lanerie and Dowd 37 ), Erysipelotrix spp.( Reference Garcia-Mazcorro, Lanerie and Dowd 37 ) or Clostridium clusters( Reference Garcia-Mazcorro, Lanerie and Dowd 37 – Reference Ritchie, Burke and Garcia-Mazcorro 39 ). In two studies, the phyla Bacteroidetes and Proteobacteria were the second and third most abundant, respectively( Reference Ritchie, Burke and Garcia-Mazcorro 39 , Reference Barry, Middelbos and Vester Boler 40 ), whereas in the study of Tun et al. ( Reference Tun, Brar and Khin 41 ) the Bacteroidetes/Chlorobi group was the predominant phylum. In contrast, two studies( Reference Garcia-Mazcorro, Lanerie and Dowd 37 , Reference Handl, Dowd and Garcia-Mazcorro 38 ) observed that the phylum Actinobacteria was the second most abundant in cats, while in the study of Desai et al. ( Reference Desai, Musil and Carr 36 ) even higher numbers of Actinobacteria were found. Likewise, in kittens and geriatric cats, the Coriobacteriaceae group, belonging to this phylum was highly present in the faecal microbiota( Reference Jia, Frantz and Khoo 42 , Reference Jia, Frantz and Khoo 43 ) (see Table 1).
The discrepancy in predominant phyla is probably due to the underestimation of Bifidobacterium spp. when using the 16S rRNA target gene( Reference Ritchie, Steiner and Suchodolski 19 , Reference Ritchie, Burke and Garcia-Mazcorro 39 , Reference Suchodolski, Camancho and Steiner 44 ) (see Table 2). Desai et al. ( Reference Desai, Musil and Carr 36 ) stated that, in general, the overall taxonomic profile is similar in domestic cats to that of most of the studies in mammals. In mammalian faecal microbiota a predominance of bacteria belonging to the phylum Firmicutes has been shown with variation in constituent bacterial species due to animal species( Reference Ley, Hamady and Lozupone 45 ). This finding was confirmed in a recent study using the newer massive parallel 16S rRNA gene pyrosequencing technique on faecal samples of pet dogs and cats( Reference Handl, Dowd and Garcia-Mazcorro 38 ).
On the species level, Sparkes et al. ( Reference Sparkes, Papasouliotis and Sunvold 46 ) observed Bacteroides spp. and Clostridium perfringens as predominant anaerobes, and E. coli, Lactobacillus spp. and Plesiomonas shigelloides as predominant aerobes in the faecal microbiota of cats. Again, Itoh et al. ( Reference Itoh, Mitsuoka and Maejima 47 ) found Bacteroidaceae and Eubacteriaceae (phylum Firmicutes) as the most predominant families in the faecal microbiota of conventionally raised cats, followed by Clostridiaceae, Streptococcacceae and Lactobacillaceae families.
A potential problem with the use of faecal samples to estimate the microbiota in the large intestine is a possible underestimation of Lactobacillus spp., especially when enumerated with general bacterial primers. The use of group-specific primers might circumvent this problem( Reference Ritchie, Burke and Garcia-Mazcorro 39 ). Additionally, the lumen and the mucosa of the gastrointestinal tract harbour bacteria, which are hypothesised to be both represented in the faecal microbiota( Reference Eckburg, Bik and Bernstein 48 ); hence, faecal samples appear to represent the ‘total’ gut microbiota, at least qualitatively. Quantitatively, the faecal excretion of mucosal bacteria might be an underestimation of the actual number of bacteria attached to the mucosa. The fraction of mucosal bacteria that is excreted in the faeces remains to be investigated. In addition, using faecal samples has advantages of non-invasive sampling techniques( Reference Desai, Musil and Carr 36 ) and a larger availability compared with intestinal fluids( Reference Handl, Dowd and Garcia-Mazcorro 38 ).
Effects of dietary fibre on the gastrointestinal microbiota
The results of the studies on the effects of supplementing fructo-oligosaccharides (FOS), short-chain FOS (sc-FOS), galacto-oligosaccharides (GOS), sc-FOS+GOS and lactosucrose( Reference Sparkes, Papasouliotis and Sunvold 24 , Reference Barry, Middelbos and Vester Boler 40 , Reference Sparkes, Papasouliotis and Sunvold 46 , Reference Johnston, Lamport and Ballèvre 49 – Reference Biagi, Cipollini and Bonaldo 53 ) on the faecal microbiota are depicted in Table 3. In most of the studies, an increase in faecal Bifidobacterium spp. was observed. Barry et al. concluded that FOS might be a useful fibre source for promotion of feline gastrointestinal health based on changes in the faecal microbiota( Reference Barry, Wojcicki and Middelbos 51 ). The lack of a significant treatment effect in the studies of Sparkes et al. ( Reference Sparkes, Papasouliotis and Sunvold 24 , Reference Sparkes, Papasouliotis and Sunvold 46 ) and a rather low response (increase of log10 0·5) in the study of Kanakupt et al. ( Reference Kanakupt, Vester Boler and Dunsford 52 ) might have been due to the low inclusion level of (sc-)FOS. When GOS or a sc-FOS + GOS combination was supplemented, the increase in Bifidobacterium spp. counts was higher (increase of log10 0·7 and 0·9, respectively)( Reference Kanakupt, Vester Boler and Dunsford 52 ). In contrast, the decreased numbers of bacteria in cats fed diets supplemented with sc-FOS in the study of Johnston et al. ( Reference Johnston, Lamport and Ballèvre 49 ) might also have been due to other differences in diet composition (different level of protein, fat and other carbohydrates). Additionally, the number of animals used in this experiment was very low (three for the control diet and four for the ‘FOS diet’)( Reference Johnston, Lamport and Ballèvre 49 ).
Table 3 Overview of the effects of dietary fibre on the feline gastrointestinal microbiota

FOS, fructo-oligosaccharides; CFU, colony-forming unit; ↑ , increase; ↓ , decrease; sc, short-chain; GOS, galacto-oligosaccharide.
* Mean of lecithinase-positive and -negative Clostridia at all time points.
† Differences between specified time point and day before administration are calculated.
‡ Absent before lactosucrose administration.
In two feline studies, the effects of a synbiotic formulation on the faecal microbiota of healthy cats were studied( Reference Garcia-Mazcorro, Lanerie and Dowd 37 , Reference Biagi, Cipollini and Bonaldo 53 ). The first study used a mixture of seven probiotic strains and a blend of FOS and arabinogalactans( Reference Garcia-Mazcorro, Lanerie and Dowd 37 ), whereas in the second study a combination of GOS and a specific Bifidobacterium strain was used( Reference Biagi, Cipollini and Bonaldo 53 ). No changes in major bacterial phyla were discovered between faecal samples before, during or after administration of the synbiotic supplement( Reference Garcia-Mazcorro, Lanerie and Dowd 37 ). During product administration, probiotic species were detected in eleven out of twelve cats, and abundances of Enterococcus spp. and Streptococcus spp. were increased in at least one time point during administration and decreased back to baseline values after discontinuation of the supplementation( Reference Garcia-Mazcorro, Lanerie and Dowd 37 ). Likewise, in another study, cats were supplemented with a probiotic for 4·5 weeks, and the supplemented species (Lactobacillus acidophilus) was recovered from faeces of cats during the supplementation period, but not before or after the supplementation was ceased( Reference Marshall-Jones, Baillon and Croft 54 ). In contrast, 10 d after cessation of daily synbiotic administration, a significant increase in faecal bifidobacteria content was observed compared with counts before supplementation( Reference Biagi, Cipollini and Bonaldo 53 ).
In vitro and in vivo fermentation studies
As stated previously, intestinal fermentation might be an important process in healthy domestic cats. A literature overview of the fermentation studies done in vitro and in domestic cats is given in the next sections. Furthermore, for several disease conditions, dietary fibre can exert beneficial effects. An overview of the effects of dietary fibre on a broad variety of diseases was beyond the scope of the present review. Therefore, the focus is on diseases in which dietary protein restriction is a treatment cornerstone, such as chronic kidney disease (CKD).
In vitro fermentation studies
All in vitro fermentation studies done in cats used fresh faecal samples as a source of inocula( Reference Sunvold, Fahey and Merchen 3 , Reference Sunvold, Fahey and Merchen 4 , Reference Barry, Wojcicki and Bauer 55 , Reference Rochus, Bosch and Vanhaecke 56 ). Samples were immediately placed in a sterile sample bag( Reference Barry, Wojcicki and Bauer 55 ) or within 15 min post-voiding in a container pre-filled with carbon dioxide and kept on crushed ice before incubation (maximum 4 h post-voiding of first sample)( Reference Rochus, Bosch and Vanhaecke 56 ). A recent study with canine faeces revealed that chilling the faecal samples for 24 h in crushed ice maintained the fermentation characteristics of substrates compared with incubation with fresh faecal inoculum( Reference Bosch, Wrigglesworth and Cone 57 ). Furthermore, in another in vitro study from the same group, faeces appeared to be applicable for inoculum preparation for in vitro screening purposes, although the fermentation in the proximal and transverse colon might be overestimated( Reference Bosch, Pellikaan and Rutten 58 ). These inocula were used to ferment different fibre sources. In the studies of Sunvold et al. ( Reference Sunvold, Fahey and Merchen 3 , Reference Sunvold, Fahey and Merchen 4 ), fermentation was described based on organic matter disappearance from the culture media and the concentrations and ratios of SCFA. Only one study in cats investigated the interdependency of fermentation characteristics of select fermentable fibres( Reference Rochus, Bosch and Vanhaecke 56 ). The main conclusions of the in vitro studies are compiled in Table 4.
Table 4 Overview of in vitro fermentation studies of plant fibres using feline faecal inoculum

↑ , Increase; FOS, fructo-oligosaccharides.
Besides the source (intestinal fluid v. faeces) of the inocula, another important factor that influences the outcome of in vitro fermentation studies is the diet of the inoculum donors( Reference Sunvold, Fahey and Merchen 4 , Reference Barry, Wojcicki and Bauer 55 ). This statement contrasts with the above-mentioned findings of Osbaldiston & Stowe, where no effect of diet on the microbiota composition was found (see the ‘Stomach’ and ‘Jejunum and ileum’ sections)( Reference Osbaldiston and Stowe 22 ). However, the latter study used culture techniques, which underestimate the diversity of the microbiota( Reference Ritchie, Steiner and Suchodolski 19 ), whereas in the in vitro system, the endproduct and metabolite concentrations and the volumes of gas produced upon fibre fermentation are measured, hence the metabolism of the microbiota as a whole is evaluated. In the study of Barry et al. ( Reference Barry, Wojcicki and Bauer 55 ), cats were adapted to different dietary fibre sources (cellulose, FOS, pectin) in vivo before fermenting these fibres in vitro. Overall, in vivo adaptation to FOS or pectin resulted in higher in vitro SCFA concentrations and more gas produced as compared with adaptation to cellulose( Reference Barry, Wojcicki and Bauer 55 ). The differences found in the present study were of a great magnitude and most probably biologically relevant considering careful extrapolation to in vivo situations (for example, total in vitro gas produced in ml/g DM averaged over the three adaptation diets: 0·0 for cellulose v. 90·5 for FOS v. 61·1 for pectin; total SCFA in mmol/g DM averaged over the three adaptation diets: 0·1 for cellulose v. 5·0 for FOS v. 2·8 for pectin)( Reference Barry, Wojcicki and Bauer 55 ). Sunvold et al. ( Reference Sunvold, Fahey and Merchen 4 ) compared in vitro organic matter disappearance and SCFA production from fermentation of different fibre sources, using inocula from cats fed a diet without supplemental fibre or supplemented with beet pulp. In vitro fermentation of fibrous substrates by faecal microbiota from cats increased when fermentable fibre was included in the donor diet( Reference Sunvold, Fahey and Merchen 4 ).
In vivo studies
The in vivo effects of dietary fibre, including oligosaccharides and animal fibre, on nutrient intake and digestibility, faecal characteristics, the morphology and function of the colon, N and energy metabolism, and some disease conditions are compiled in this section.
Effects of dietary fibre on nutrient intake, nutrient digestibility and faecal characteristics: viscous fibres
Sunvold et al. performed in vivo investigations following in vitro fermentation trials( Reference Sunvold, Fahey and Merchen 3 , Reference Sunvold, Fahey and Merchen 4 ). These authors concluded that the in vitro method appeared to be a good estimator of in vivo fermentation with the exception of the most fermentable fibres (pectin and gums with high viscosity). However, the in vivo fermentation calculation, based on the comparison of organic matter in food and faeces, was lower than could be predicted in vitro because of a decrease in digestibility of the other nutrients in the diet( Reference Sunvold, Fahey and Merchen 4 ).
Besides difficulties of extrapolating in vitro data, another disadvantage of supplementing viscous fibres in vivo was an increased defecation frequency and a poor stool quality with a supplementation level of 9·5 % total dietary fibre( Reference Sunvold, Fahey and Merchen 3 ). Loose stools with a strong odour were also confirmed by Bueno et al. ( Reference Bueno, Cappel and Sunvold 11 , Reference Bueno, Cappel and Sunvold 12 ) when a pectin–gum arabic blend was included in the diets (total dietary fibre 8·6 %). It has to be noted that the doses used in the above-mentioned experiments are very high. Barry et al. ( Reference Barry, Wojcicki and Middelbos 51 ) supplemented a lower dose of pectin (4 % as fed) to domestic cats and observed softer faeces as compared with the control group, cellulose. The decrease was, however, small and the authors concluded that pectin, like FOS, might be a useful fibre source for the promotion of feline gastrointestinal health( Reference Barry, Wojcicki and Middelbos 51 ). Sunvold et al. ( Reference Sunvold, Fahey and Merchen 3 ), on the contrary, advised the use of a moderately fermentable fibre source, such as beet pulp, in feline diets (see the ‘Effects of dietary fibre on nutrient intake, nutrient digestibility and faecal characteristics: moderate (beet pulp) and less fermentable fibre sources’ section). In a recent feline study, another viscous fibre source, guar gum, was used as the sole source of fermentable soluble fibre, supplemented to a moderate-protein diet( Reference Rochus, Janssens and Van de Velde 59 ). The apparent protein digestibility coefficient tended to be lower in guar gum- (71·8 (sem 3·6) %) than in cellulose (79·7 (sem 1·0) %)-supplemented cats, and the faecal and plasma metabolites from protein fermentation (for example, faecal ammonia in mg/l: 390 for guar gum v. 227 for cellulose; plasma isovaleryl- +2-methylbutyrylcarnitine in μmol/l: 0·32 (sem 0·06) for guar gum v. 0·21 (sem 0·05) for cellulose) were higher in the former cats, which confirmed highly viscous fibres to be less suitable soluble fibre sources for in vivo use in domestic cats( Reference Rochus, Janssens and Van de Velde 59 ).
Effects of dietary fibre on nutrient intake, nutrient digestibility and faecal characteristics: fructans, mannanoligosaccharides, galacto-oligosaccharides and lactosucrose
Studies on the effect of fructans (FOS, oligofructose, inulin), mannano-oligosaccharides (MOS) and GOS on faecal characteristics (consistency score and pH) are shown in Table 5 ( Reference Terada, Hara and Kato 50 – Reference Biagi, Cipollini and Bonaldo 53 , Reference Hesta, Janssens and Debraekeleer 60 – Reference Aquino, Saad and Santos 62 ). Differences in results might be explained by differences in the levels of supplementation among studies: for example, up to 9·0 % as fed oligofructose in Hesta et al. ( Reference Hesta, Janssens and Debraekeleer 60 ); maximum 0·6 % DM MOS in Aquino et al. ( Reference Aquino, Saad and Santos 62 ); 1 % synbiotic in Biagi et al. ( Reference Biagi, Cipollini and Bonaldo 53 ).
Table 5 Overview of the effects of dietary fructans, mannanoligosaccharides (MOS), galacto-oligosaccharides (GOS) and lactosucrose on faecal characteristics in cats

OF, oligofructose; FOS, fructo-oligosaccharides; ↑ , increase; sc, short-chain.
* Consistency score: 1 represented watery diarrhoea; 3, normal consistency; and 5, constipation(60).
† Consistency score: 1 represented dry, hard pellets; 2, hard, dry, formed stool; 3, soft, formed, moist stool; 4, soft, unformed stool that assumes form of container; 5, watery liquid that can poured( Reference Barry, Wojcicki and Middelbos 51 ).
Decreased faecal consistency (looser stools) can have implications for the host animal, for the use of fibres in the pet food industry and for the appreciation of the foods by pet owners. On the contrary, the effects on stool consistency can be advantageous in the treatment of cats with constipation( Reference Freiche, Houston and Weese 8 ). A decrease in faecal pH is caused by an increased production of bacterial endproducts, such as lactic acid and SCFA, in the hindgut. This decrease can exert several effects both on the microbiota and the host animal, such as stimulation of the growth of beneficial bacteria like Lactobacillus spp.( Reference Bergman 63 ) or an increased mineral absorption from the hindgut( Reference Raschka and Daniel 64 , Reference Xiao, Li and Min 65 ). Likewise, the absorption of ammonia from the hindgut can be reduced by decreasing pH. Lactic acid molecules produced can be converted to weaker acids, such as acetic, propionic and butyric acids, by cross-feeding bacteria( Reference Duncan, Louis and Flint 66 ), which prevents a severe decrease in pH and the development of lactic acidosis. The latter disease condition can be observed in cats with gastrointestinal disease( Reference Packer, Moore and Chang 67 ).
The above-mentioned SCFA have been associated with different beneficial effects on the hosts' general health( Reference Salminen, Bouley and Boutron-Ruault 68 ) and the function of the gastrointestinal tract( Reference Bueno, Cappel and Sunvold 11 , Reference Bueno, Cappel and Sunvold 12 ). Besides faecal characteristics, the in vivo studies listed in Table 5 also investigated the effects of various oligosaccharides on nutrient digestibility. In the study of Hesta et al. ( Reference Hesta, Janssens and Debraekeleer 60 ), the apparent digestibility of protein decreased significantly as the level of fructan inclusion increased (from 87·0 % with 0 % inulin to 82·8 and 77·3 % with 3 and 6 % inulin inclusion, respectively; 83·1 % with 3 % oligofructose inclusion in the diet), which was confirmed with the supplementation of sc-FOS + GOS to feline diets( Reference Kanakupt, Vester Boler and Dunsford 52 ) (from 84·2 % in control to 81·9 % in sc-FOS + GOS). The latter decrease( Reference Kanakupt, Vester Boler and Dunsford 52 ) was lower than compared with the study of Hesta et al. ( Reference Hesta, Janssens and Debraekeleer 60 ), most probably caused by the lower inclusion levels of the fibres. The decreased protein digestibility was probably due to a higher faecal excretion of bacterial protein with a higher level of fructan in the diet( Reference Hesta, Janssens and Debraekeleer 60 ) (from 25·4 % with 0 % inulin to 31·1 and 35·2 % with 3 and 6 % inulin inclusion, respectively; 32·8 % with 3 % oligofructose inclusion in the diet). Low-level MOS supplementation did not affect nutrient digestibility when supplemented to a wet diet, but slightly improved the DM digestibility if supplemented to a dry commercial diet( Reference Aquino, Saad and Santos 62 ) (from 73 (sd 4) % in control to 76 (sd 4) % in a 0·6 % MOS diet). In addition, a beneficial effect on palatability was only seen with MOS supplementation to the dry diet (for example, 11 % increase in DM intake between control and the 0·6 % dry diet)( Reference Aquino, Saad and Santos 62 ). According to these authors, MOS is thus preferably supplemented to dry diets( Reference Aquino, Saad and Santos 62 ).
A decreased ileal protein digestibility might result in an increased large-intestinal protein and amino acid fermentation( Reference Rochus, Janssens and Van de Velde 59 , Reference Hendriks, van Baal and Bosch 69 ). Besides SCFA, microbial degradation of amino acids can result in putrefactive endproducts, such as branched-chain fatty acids, valeric acid, ammonia (NH3) and phenolic compounds (for example, indole, p-cresol, phenol)( Reference Mafra, Barros and Fouque 70 ). Some putrefactive endproducts, such as polyamines, appear to be required for normal development and repair of the gastrointestinal tract( Reference Wang and Johnson 71 , Reference Loser, Eisel and Harms 72 ). However, many of these compounds are suggested to be related to colorectal disease in humans and rats( Reference Corpet, Yin and Zhang 73 – Reference Pedersen, Brynskov and Saermark 75 ). Different sources of dietary fibres have, therefore, been applied in domestic cats in an attempt to reduce the production and excretion of these potentially harmful substances and to attempt the reduction of faecal odour( Reference Terada, Hara and Kato 50 – Reference Biagi, Cipollini and Bonaldo 53 , Reference Hesta, Hoornaert and Verlinden 61 , Reference Barry, Hernot and Van Loo 76 ). Faecal concentrations of ammonia (339 (sd 210) μg/g wet faeces before supplementation), indole (48 (sd 19) μg/g wet faeces before supplementation), ethylphenol (20 (sd 6) μg/g wet faeces before supplementation) and urinary ammonia (17 (sd 8) mg/ml urine before supplementation) were reduced significantly on day 14 of lactosucrose administration( Reference Terada, Hara and Kato 50 ) (to 162 (sd 34), 30 (sd 12), 8 (sd 3) μg/g wet faeces and 10 (sd 4) mg/ml urine, respectively). This decrease might be explained by the concomitant decrease in counts of Clostridia ( − 0·25 log numbers per g faeces for the average of lecithinase-positive and -negative Clostridia, difference between before and after 14 d of lactosucrose administration) and Enterobacteriaceae ( − 1·6 log numbers per g faeces, difference between before and after 14 d of lactosucrose administration) due to the lactosucrose supplementation, as both groups are known to produce these putrefactive substances. In addition, the environmental ammonia (ammonia in room from 22 (sd 2) parts per million (ppm) before administration to 16 (sd 1) ppm after 14 d of administration) and the faecal odour (no quantitative data available) decreased remarkably during administration( Reference Terada, Hara and Kato 50 ). Likewise, supplementation of oligofructose led to decreased faecal concentrations of histamine, spermidine and indole( Reference Barry, Hernot and Van Loo 76 ) (quantitative data not available), and a synbiotic combination of GOS and a Bifidobacterium strain decreased faecal ammonia concentrations (in μmol/g faecal DM) even 10 d post-supplementation( Reference Biagi, Cipollini and Bonaldo 53 ) (from 353 before to 288 at 1 d post- administration and 281 at 10 d post-administration). In contrast, Hesta et al. ( Reference Hesta, Hoornaert and Verlinden 61 ) found no effects of FOS supplementation to cats on twenty-seven different odour components, and Kanakupt et al. ( Reference Kanakupt, Vester Boler and Dunsford 52 ) observed no differences in faecal protein catabolites among control, sc-FOS-, GOS- and sc-FOS + GOS-supplemented cats. In the latter study, no differences in protein catabolite-producing bacteria were observed( Reference Kanakupt, Vester Boler and Dunsford 52 ). Increased faecal concentrations of ammonia (small differences), 4-methyl phenol and indole were observed when feline diets were supplemented with FOS (differences compared with cellulose in mmol or μmol/g of faecal DM: 0·1, 2·1 and 1·0, respectively) and pectin (differences compared with cellulose in mmol or μmol/g of faecal DM: 0·1, 1·9 and 0·7, respectively), possibly due to the fast fermentation of both supplements( Reference Barry, Wojcicki and Middelbos 51 ). Different outcomes in the studies might again be explained by different sources and inclusion levels of fibres.
Effects of dietary fibre on nutrient intake, nutrient digestibility and faecal characteristics: moderate (beet pulp) and less fermentable fibre sources
The effects of moderately and less-fermentable fibre sources on nutrient intake have been studied by different groups. Fekete et al. ( Reference Fekete, Hullár and Andrásofszky 77 ) observed slightly different DM intake among diets supplemented with high levels (10 % DM) of beet pulp (263 (sd 42) g/4 d), peanut hulls (274 (sd 31) g/4 d) and alfalfa meal (257 (sd 37) g/4 d). In contrast, Sunvold et al. ( Reference Sunvold, Fahey and Merchen 3 ) did not observe differences in DM, organic matter or N intake between beet pulp- and cellulose-supplemented cats and cats on a control diet without supplemented fibre. Both studies used a similar inclusion level of the dietary fibre sources. Likewise, the inclusion of cellulose at a high level (17 % DM) did not alter food intake in cats( Reference Prola, Dobenecker and Mussa 78 , Reference Prola, Dobenecker and Kienzle 79 ) and the addition of psyllium husks and seeds did not decrease diet acceptance in cats with constipation( Reference Freiche, Houston and Weese 8 ).
In all the above-mentioned studies nutrient digestibility was studied as well. In peanut hull- and alfalfa meal-supplemented cats, a decreased DM digestibility ( − 22·4 and − 7·2 % compared with the control diet) was seen( Reference Fekete, Hullár and Andrásofszky 77 ), which was confirmed in healthy beet pulp-( Reference Sunvold, Fahey and Merchen 3 ) and cellulose( Reference Prola, Dobenecker and Mussa 78 )-supplemented cats ( − 7·6 and − 15 % (average of the three types of cellulose used) compared with control). Likewise, another study confirmed a decreased DM digestibility when diets of overweight cats were supplemented with beet pulp ( − 11 % compared with control), wheat bran ( − 13 % compared with control) or sugarcane fibre ( − 21 % compared with control)( Reference Fischer, Kessler and de Sá 9 ). Only peanut hull and alfalfa meal supplementation decreased protein digestibility( Reference Fekete, Hullár and Andrásofszky 77 ) ( − 10 and − 2 % compared with control, respectively). Likewise, Fischer et al. ( Reference Fischer, Kessler and de Sá 9 ) observed different effects on protein and fat digestibility depending on the chemical composition of the supplemented fibre source. No other studies in which cats were supplemented with peanut hulls, alfalfa meal, wheat bran or sugarcane fibre were found. Conclusively, moderately fermentable fibre sources, such as beet pulp, appear to be beneficial for normal-weight cats. In contrast, low-fermentable fibres, such as sugarcane fibre, might have adequate properties in low-energy weight-loss diets( Reference Fischer, Kessler and de Sá 9 ). However, the high water-binding activity of the latter fibre source, comparable with that of long-fibre cellulose( Reference Prola, Dobenecker and Kienzle 79 ), can lead to extremely dry faeces, limiting the inclusion level in the diet( Reference Fischer, Kessler and de Sá 9 ). This problem might be overcome by using short-fibre or microcrystalline cellulose( Reference Prola, Dobenecker and Kienzle 79 ). An important factor in determining the outcome of the supplementation of fibres is the inclusion level, which was very high in the above-mentioned studies. Therefore, more research using lower levels is warranted.
Effects of dietary fibre on nutrient intake, nutrient digestibility and faecal characteristics: animal fibre
Differences in nutrient digestibility and fermentation endproducts between a high-protein, extruded feline diet and a raw and cooked beef-based diet, which is a source of animal fibre, were studied by Kerr et al. ( Reference Kerr, Vester Boler and Morris 18 ). The extruded diet had a much higher protein content (57 % crude protein) compared with a conventional extruded feline diet (for adult cats between 32 and 35 % crude protein) to mimic the composition of the beef-based diets. The effect of processing of the diets was investigated. However, due to a different ingredient composition between the extruded and beef-based diets, this comparison was biased. The extruded diet contained, for example, chicken meal as the major protein source. The latter might have contained a considerable amount of animal fibre from cartilage, for example, and might be considerably less digestible than beef meat. Two other studies that investigated the effects of feeding raw-meat diets on nutrient digestibility and faecal characteristics in domestic cats were found( Reference Vester, Beloshapka and Middelbos 80 , Reference Kerr, Beloshapka and Morris 81 ). Comparisons of results between both studies are again biased by the use of different ingredient sources, such as different sources of plant fibre. Further studies using diets with an equal ingredient composition of all diets are necessary to study the effects of animal fibre fermentation on nutrient digestibility, the host's metabolism and the interaction with plant fibres in domestic cats.
Effects of dietary fibre on colonic morphology and function
The influence of fibre fermentation and concomitant SCFA production on colonic morphology was studied in adult healthy cats by Bueno et al. ( Reference Bueno, Cappel and Sunvold 11 , Reference Bueno, Cappel and Sunvold 12 ), and in overweight cats by Fischer et al. ( Reference Fischer, Kessler and de Sá 9 ). Beneficial effects on morphology (colonic weight, mucosal cell density, crypt structure), function (mucosal tissue energetics, transport of SCFA, etc.) and microbiota of the colon (less pathogenic bacteria) were observed( Reference Bueno, Cappel and Sunvold 11 , Reference Bueno, Cappel and Sunvold 12 ). According to these authors, the supplementation of a moderately fermentable fibre source to feline diets generated the best combination of beneficial effects on morphology, function and microbiota. In contrast, in overweight cats, the supplementation of beet pulp, wheat bran or sugarcane fibre did not affect the histological image of the colon biopsies( Reference Fischer, Kessler and de Sá 9 ). Nevertheless, these fibres demonstrated other beneficial properties upon in vivo fermentation, as stated in other parts of the present review.
Effects of dietary fibre on nitrogen and energy metabolism
The principle of the N trap is that blood urea concentrations decrease when fermentable fibre is supplemented to the diet. Fermentable fibre stimulates the growth of and provides energy to the anaerobic micro-organisms in the large intestine( Reference Younes, Garleb and Behr 82 ). For bacterial protein anabolism, not only an energy source, such as fermentable fibre, but also a source of N should be available( Reference Bliss 83 ). N sources include undigested dietary protein entering the large intestine, endogenous protein and blood urea( Reference Younes, Garleb and Behr 82 ). Bacterial protein is not absorbed in the large intestine, but is excreted in the faeces( Reference Rutgers, Biourge, Pibot, Biourge and Elliott 7 ). When blood urea is the major N source to the intestinal microbiota, blood urea concentrations decrease and a decreased N excretion by the kidneys is observed, while N excretion in the faeces increases. Furthermore, fermentable fibre increased caecal blood flow in rats( Reference Younes, Garleb and Behr 82 ), which might enhance the passive diffusion of urea from the blood to the intestine. The N trap hypothesis has been proven in the rat( Reference Younes, Garleb and Behr 82 , Reference Younes, Rémésy and Behr 84 ) and dog( Reference Howard, Kerley and Sunvold 85 ), whereas in the cat tendencies towards an N shift from urine to faeces were found using diets supplemented with oligofructose( Reference Hesta, Hoornaert and Verlinden 61 , Reference Barry, Hernot and Van Loo 76 ). Verbrugghe et al. ( Reference Verbrugghe, Hesta and Gommeren 86 – Reference Verbrugghe, Hesta and Daminet 88 ) studied the effects of oligofructose and inulin on glucose and amino acid metabolism in domestic cats. In the first study( Reference Verbrugghe, Hesta and Gommeren 86 ), a control diet was tested against a prebiotic diet with 2·5 % of a blend of oligofructose and inulin. Diets did not affect fasting plasma glucose and insulin concentrations, blood glucose and insulin responses to glucose administration, or area under the glucose and insulin curves. In contrast, a decreased mean blood glucose concentration and area under the curve were achieved by supplementing sugarcane fibre to diets of overweight cats( Reference Fischer, Kessler and de Sá 9 ). Despite an apparent absence of effects on carbohydrate metabolism in the study of Verbrugghe et al. ( Reference Verbrugghe, Hesta and Gommeren 86 ), analysis of plasma acylcarnitine profiles revealed higher propionylcarnitine concentrations when the prebiotic diet was fed, suggesting colonic fermentation and propionate absorption. Prebiotic supplementation reduced methylmalonylcarnitine and aspartate aminotransferase concentrations, both indicating reduced gluconeogenesis from amino acids( Reference Verbrugghe, Hesta and Gommeren 86 ). Further studies confirmed the amino acid-sparing potential of propionic acid from dietary fructan fermentation( Reference Verbrugghe, Janssens and Meininger 87 ) and signs of sparing of the amino acid valine with propionylated high-amylose maize starch supplementation provided that protein intake was sufficient( Reference Rochus, Janssens and Cools 89 ). The quantification of the amino acid-sparing potential of propionic acid in healthy and disease-inflicted cats remains to be done. In contrast, this mechanism was not confirmed with guar gum supplementation, due to its high viscosity( Reference Rochus, Janssens and Van de Velde 59 ). The guar gum's high viscosity impaired the small-intestinal protein digestion, causing a large load of undigested protein being fermented in the large intestine, which biased the proper assessment of the amino acid sparing by the produced propionic acid( Reference Rochus, Janssens and Van de Velde 59 ). Further research using guar gum with lower viscosity might be warranted. The applicability of low-viscous guar gum might be restricted to dry extruded feline diets or homemade diets, since, for the production of canned diets, the gelling properties of highly viscous guar gum are advantageous.
Effects of dietary fibre on diseases with dietary protein restriction as a treatment cornerstone, such as chronic kidney and liver disease
CKD is a very common disease in middle-aged to elderly cats( Reference Ross, Osborne and Kirk 90 ). The cornerstones of the dietary management of CKD are a modification of the quantity (decrease) and quality (increase) of protein and a restriction of the dietary P intake( Reference Polzin, Osborne and Ross 91 ). Liver disease encompasses a range of different aetiologies, such as hepatic lipidosis and portosystemic shunts. The symptomatic treatment of liver disease, specifically in case of hyperammonaemia and hepatic encephalopathy, is based on dietary protein restriction( Reference Bunch, Nelson and Guillermo Couto 92 , Reference Center 93 ). Additionally, the supplementation of dietary fibre can exert beneficial effects on these disease conditions and the mechanisms behind these effects are explained in the next paragraphs.
The N trap principle, explained in the previous section, might be advantageous in animals suffering from hyperammonaemia (liver disease) or azotaemia (CKD), since an increase in the faecal N excretion and decreases in blood urea and ammonia concentrations can be achieved( Reference Younes, Garleb and Behr 82 ). Until now, no studies investigating the N trap principle have been performed in cats with CKD. As mentioned above (‘Effects of dietary fibre on nutrient intake, nutrient digestibility and faecal characteristics: fructans, mannanoligosaccharides and galacto-oligosaccharides’ section), a decrease in large-intestinal pH occurs when SCFA and lactate are produced upon fermentation of dietary fibre. Another consequence of this decrease, besides the ones mentioned above, is that the overload of protons (H+), that are present in a more acidic environment, leads to the ionisation of the ammonia (NH3) molecules present to ammonium (NH4 +) ions( Reference Younes, Garleb and Behr 82 ). The absorption of NH4 + ions from the intestine to the bloodstream is far less effective, and the majority of these ions are excreted in the faeces( Reference Cummings 94 ). That way, the ammonia concentration in the blood decreases, which benefits patients with hyperammonaemia( Reference Rutgers, Biourge, Pibot, Biourge and Elliott 7 ). Furthermore, less urea will be produced in the liver, resulting in lower blood urea N concentrations, beneficial to azotaemic patients. Additionally, the recently suggested amino acid-sparing potential of propionic acid( Reference Verbrugghe, Hesta and Gommeren 86 – Reference Rochus, Janssens and Cools 89 ) (see above) might be advantageous to patients whose diseases urge a dietary protein restriction, such as kidney( Reference Polzin, Osborne and Ross 91 ) or liver( Reference Bunch, Nelson and Guillermo Couto 92 , Reference Center 93 ) disease patients with evidence of hyperammonaemia or hepatic encephalopathy.
Conclusions
Despite the growing interest in dietary fibre supplementation to feline nutrition, research on this topic remains scarcer in cats compared with dogs and humans. The newest molecular techniques for qualitative and quantitative assessment of the microbiota have been applied to feline samples of different parts of the feline gastrointestinal tract. Therefore, a detailed assessment of the complex and diverse microbiota is available in the literature. Furthermore, in vitro batch culture systems are assumed to be suitable for screening several indigestible substrates for fermentation kinetics and endproduct concentrations and this technique has been used in several animal species. However, the use of this technique with feline inoculum remains scarce, despite the fact that valuable estimations of the fermentation potential and kinetics of various fibre sources for felines can be gained non-invasively. Research on in vivo fermentation is rather scarce in cats, despite the fact that fermentation endproducts might appear to exert different beneficial effects on the host animal. More research is warranted to reveal potential (plant or animal) fibre sources that can be used on a large scale in feline nutrition for healthy and diseased cats.
Acknowledgements
The present review was written as a part of the postgraduate study of the first author (K. R.), which was funded by the Institute for Promotion of Innovation through Science and Technology in Flanders (IWT; grant no. 091050 to K. R.). The IWT had no role in the design, analysis or writing of the article.
K. R. was responsible for the manuscript drafting. M. H. and G. P. J. J., promoter and co-promoter of K. R., respectively, contributed to the manuscript drafting.
There are no conflicts of interest.






