



In antiquity, just as nowadays, travelers who were away from home for more than a day would need a place to spend the night. One could find many private and official accommodations along major roads and in the cities of the Roman-Byzantine Empire in the Middle East. For instance, the so-called mansiones of the cursus publicus provided food and accommodation for all those who were officially permitted to use their services.Footnote 1 Although supervised by Roman officials, in general they seem to have been established and financed by local communities, who were required to support traveling officials and military officers.Footnote 2 Some of the mansiones also had mutationes attached to them, stations where transport animals could be exchanged and fed. These mutationes could also stand alone and they were similarly financed and maintained by local residents.
The archaeological project at Samra in Jordan discovered such a road station during excavations of a mound close to the ancient Roman-Byzantine village of Samra and adjoining the Via Nova Traiana (Fig. 1).Footnote 3 Notably, this route represents the most widely attested feature of the Roman annexation of the Nabataean kingdom, which took place in 106 CE and led to the creation of the Province of Arabia.Footnote 4 The construction of the road started shortly after this annexation, between 107 and 114 CE, supervised by governor Gaius Claudius Severus and completed under Hadrian. This north–south highway was built to connect the capital of the new province, Bosra, to the port of Ayla (Aqaba) on the Red Sea, a distance of approximately 430 km.
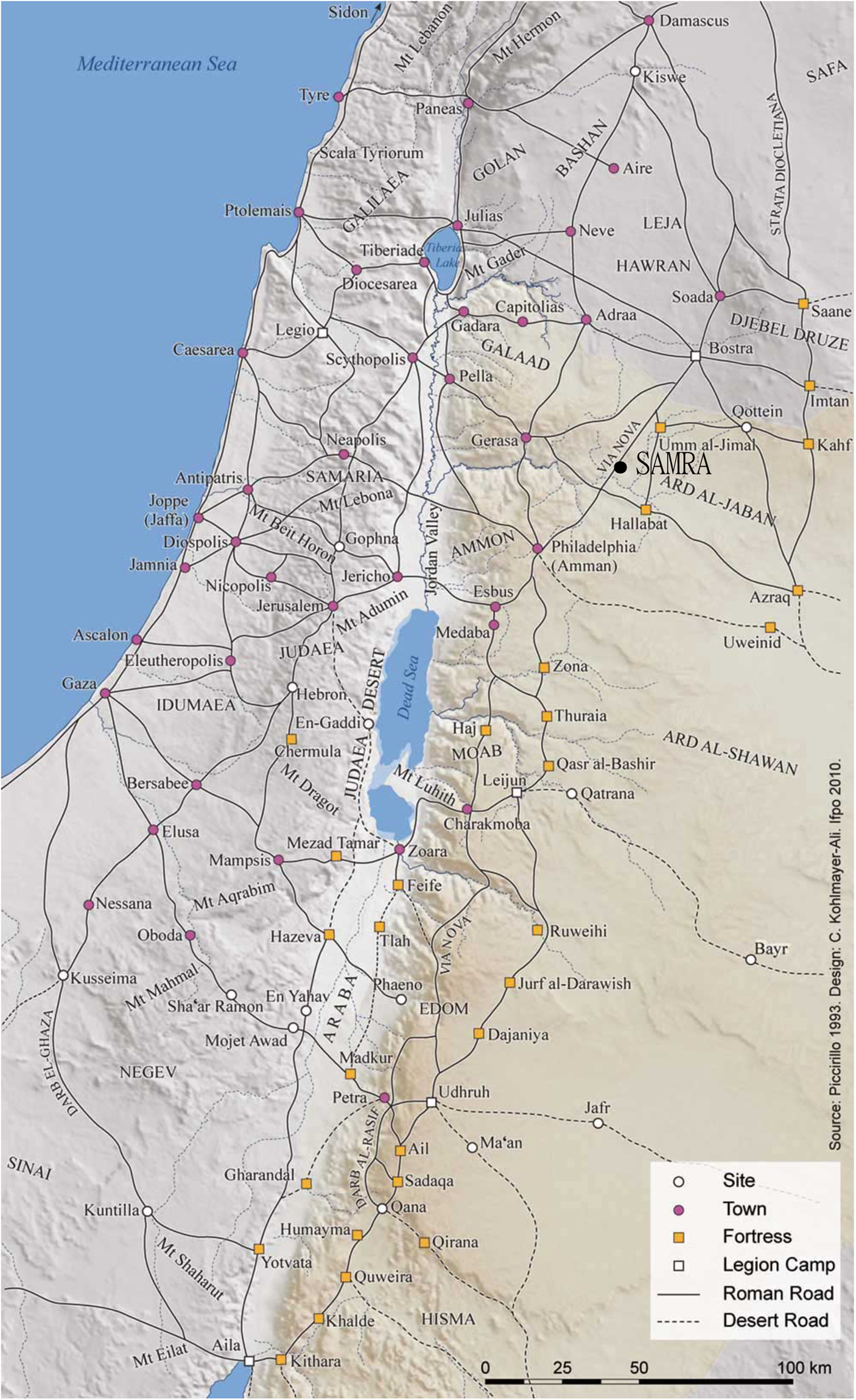
Fig. 1. Location of Samra on the Via Nova Traiana. (Map from Augé Reference Augé and Ababsa2016; © IFPO.)
Given that it is a freestanding construction, and because of its layout and location, the Samra road station was recognized as a Roman mansio. Its occupation did not last long – from 150 to 250 CE – but a surprisingly large quantity of faunal remains were found. This exceptional assemblage facilitates reconstruction of the role of meat in the occupants’ diet and its procurement for travelers along the Via Nova Traiana.
The site
Khirbet es-Samra, a brief history
Referred to as “Hatita” on the Peutinger Table, the site of Khirbet es-Samra, built on the basaltic plateau, was a village built along the Roman highway known as the Via Nova at a point 40 km northeast of Amman.Footnote 5 The location of the mansio away from the village on the edge of the highway, as well as the presence of a fortress, attest to Roman occupation in the 2nd and 3rd c. CE. At the end of the 4th c. CE, the site was reorganized under the emperors Valentinian I, Valens, and Gratian. During the 6th c., the site lost its military function but remained a Christian village. Eleven small churches have been found there, some decorated with mosaics adorned with geometric patterns, animals, and depictions of towns with their defensive walls and their churches, and also bearing Greek inscriptions. The prosperity of this village seems to have peaked in the early 7th c., on the eve of the Muslim conquest. During the 7th and 8th c., under the Umayyads, the village was still Christian. Like many places in the same region, it declined thereafter. The remains of a small mosque, probably dating from the Ayyubid or Mamluk period (13th–15th c. CE), constructed within the ruins of a Byzantine church, were discovered there. The site was probably abandoned during the 15th c. CE and remained deserted until the 20th c. CE. Today it is called Khirbet es-Samra, “the dark ruins,” and since 1981 the site has been studied by the École Biblique et Archéologique Française de Jérusalem and the CNRS.
The mansio discussed in this article was discovered about 600 m from the Byzantine village of Samra (Fig. 2). This type of architectural structure is widespread in the Mediterranean world but much rarer in the Levant. Nevertheless, as pointed out by M.-A. Le Guennec, identifying a Roman road station on the ground is a complex process.Footnote 6 We have no direct information concerning the official status of this building, which could also have been a statio, a taberna or a mutatio. In their publications, numerous archaeologists have raised the question of the use of this type of building for highway service and, in particular, in relation to the cursus publicus. They have questioned the role of uncovered buildings in the Roman occupation of the territories in which they are found, since they are not linked to the official administration.Footnote 7 Here, I will use the term mansio in its broadest sense of the term, meaning a road station or a travel stop.

Fig. 2. Aerial view of the village of Samra, 1964. (© Royal Jordanian Geographical Center.)
The Samra Roman mansio: a brief architectural description
The foundations make the layout of this building absolutely clear, showing a regular and symmetrical structure (30 × 24 m) running along an east–west axis (Fig. 3). The building had been heavily robbed during antiquity and the stones of the walls were probably removed at the beginning of the construction of the Roman village. A collapsed layer of clay debris covers the site in an irregular manner and came from the crude mortar of the walls and ceilings. Below this layer, the original floor is almost intact in most parts of the building. It was made of compact clay with a white surface.

Fig. 3. Top: plan of the road station of Samra (the numbers indicate the room loci); bottom: 3D reconstruction of Samra's mansio. (© L. Monchamps/EBAF.)
The central part of the building has a square plan, a common form attested throughout the Greco-Roman world, in which the rooms (Loci 5, 6, 7, 10, and 11) are distributed around an almost rectangular inner courtyard (Locus 3: 9 × 8 m). The presence of a drain, a bench, and a small podium suggests that Locus 11 would have served as a bathroom. Locus 4 is a small room that includes a square cubbyhole of 2 × 2 m (Locus 15), while in its southwest corner (with an off-center door) is what may have been a stairwell providing access to another floor or a terrace. This small space could, moreover, act as a guard's lodge, since it monitored comings and goings. Off this central space, the distribution of the living quarters is marked by huge stone jambs for the doors.
The space devoted to accommodation (Loci 5, 6, 7, 10, and 11) is easily distinguished from the long rooms added to the south (Locus 9), west (Locus 1), and north (Locus 8), which give the establishment its specific character of a mansio. All the floors were made of lime mortar, except in the courtyard, where they were made of clay. On the western face of the structure is the entrance or reception (Locus 1), where visitors dismounted. The threshold has disappeared but a door was set in the façade at its center. This entrance was directly connected to the pavement of the road. It is not known if the entrance was covered. It may have been where less prestigious means of transport, such as donkeys, were kept. Repeated passage through this entrance has created a trough-like depression on the floor whose irregular shape is indicative of a path. It is assumed that the draft and pack animals crossed the threshold and proceeded to either the right or the left, toward the stables (Loci 8 and 9), and in so doing eroded the beaten earth floor. A narrow indentation in the floor marks the path of the travelers toward the accommodation area via a vestibule (Locus 2), which has a small semicircular tub built in mortared masonry that allowed the travelers to perform their ablutions before moving further inside. Drinking water was drawn from a small cistern to the south, in the curve of the road. The water for the animals and the ablution area was probably taken with buckets from a pool (birkeh) at the back of the building.
The abundant pottery found in the mansio is well dated, thanks to the chronological markers provided by coins, to between 130/150 and 250/280 CE. For the most part, it is tableware rather than related to food preparation and cooking (in the latter category, mainly pots and some saucepans). Locally-made jars are not numerous and imported amphorae are rare, as are storage containers. On the other hand, thin and painted Nabataean plates are well represented. Other types of fineware were found but in smaller quantities, as shown by the fragments of eastern sigillata and plates with a yellow fabric and red painted decoration, or the so-called African ware, with thin walls and dark, pinkish-red varnish. The study of the workshops and the origins of the pottery is still in progress.
The road station was abandoned and not destroyed by fire, perhaps after part of the Roman road fell out of use. The stone walls, rare in this landscape, were certainly reused for the construction of the village.
Methodology
Species identification
Bone specimens were identified down to the lowest possible taxonomic category and anatomical element, or portion thereof. However, where there were few morphological features (such as with many mid-shaft long-bone fragments or fragments of vertebrae or ribs, due to high degrees of bone fragmentation), these fragments were recorded according to three size categories: SH: small-sized herbivore (i.e., unidentified caprine or gazelle size); LH: large-sized herbivore (unidentified equine, cattle, or camel size); and Ind: indeterminate mammalian fragments.
Quantification
The method for determining the number of identifiable specimens used in this study is the most widely used faunal parameter available for the characterization of faunal assemblages from archaeological excavations. The quantification of bone remains is based on the total number of identified specimens (NISP) and on the estimated minimum number of individuals (MNI). The MNI is defined as “the minimum number of [complete] individual animals necessary to account for the specimens observed” and was estimated on the basis of the frequency of the best-represented bones, taking into account the laterality.Footnote 8
Aging
To estimate the age at death and thus obtain a mortality (slaughter) profile, two main methods were used. The first assessed the stage of tooth eruption and analyzed dental wear. For sheep and goats, I refer to the work of Payne, and for pigs to the work of Horard-Herbin.Footnote 9 The second method – epiphyseal fusion of long bones – is less reliable as a result of the various taphonomic processes affecting skeletal remains, especially those of immature individuals and those with a high marrow and trabecula content. For caprines, cattle, and equids, ages of bone fusion published by Barone were used.Footnote 10
Bone surface modifications
Cut and chop marks
Butchery consists of a set, or series of sets, of human activities directed toward the extraction of consumable resources from a carcass. It has a temporal duration, made up of the set and order of activities carried out to extract these resources from a carcass (i.e., butchering pattern).Footnote 11 Consequently, during butchery, anthropogenic marks are the result of several carcass-processing activities, including skinning, dismemberment, or disarticulation, and meat removal (filleting).Footnote 12 Two categories of damage resulting from tool use were identified, namely cut marks and chop marks.Footnote 13 Cut marks are incisions resulting from the cutting movement of a sharp-edged implement on the bone surface. They are elongated, linear striations of variable length and width. Chop marks are defined as broad, deep, and relatively linear depressions that often have a V-shaped cross section. Internal striations within the main groove may be observed. Chop marks are the result of a heavy blow to the bone with a sharp implement.
Burning
Bones may show traces of combustion that can be directly or indirectly linked to human action. It is very unusual for cooking to leave burning marks, unless something goes seriously wrong. Indeed, a bone broth serves generally to recover fat and in such cases most of the bone fragments are not charred. Roasting/grilling may leave burn marks at the ends of bones, and it should not be forgotten that the fresh bones could have been used as fuel. Alternatively, bones may have been burned by accident, by falling into a hearth or being deposited under a fire. The burned bones can be grouped by color, which is directly related to the intensity of combustion.Footnote 14
Carnivore traces
In zooarchaeology, the characteristics of carnivore damage have been studied to determine the effect that these species could have on a faunal assemblage both through the removal/destruction of bones and through alterations made to them. The main traces left by carnivores on bones are chewing, gnawing, score marks, furrows, and tooth punctures.Footnote 15
Results
The faunal list
The Roman levels of this building yielded 5,180 faunal remains which were recovered after sieving; they represented mammals, birds, fish, seashells, and unidentified bones (Table 1). Overall, it was possible to identify 23.0% to the level of species and 38.8% to the level of skeletal element. Therefore, unidentifiable bone fragments form 61.2% of the sample; their abundance is the result of intense weathering, which has resulted in greatly deteriorated bone surfaces and numerous tooth enamel fragments.Footnote 16 Corrosion on the bones’ surface is noted, which has especially affected many body parts of both birds and small mammals, and is linked to micro-organisms which are found more often in soils with a pH between 6 and 7.5, such as basaltic soils.Footnote 17
Table 1. The Samra mansio species list given as number of identified specimens (NISP), frequency of identified specimens (%), and minimum number of individuals (MNI). (Two human teeth were also identified.)
The species consumed
The bone assemblage is largely dominated by the classic domestic triad, caprine/cattle/pig (75.6% of the identified species). Caprine meat (sheep/goat) was widely consumed in many Middle Eastern sites in Roman times, followed by cattle and pig, the bones of which are less abundant.Footnote 18 The mansio of Samra fits perfectly into this regional scheme with a high frequency of sheep/goat bones (Fig. 4).
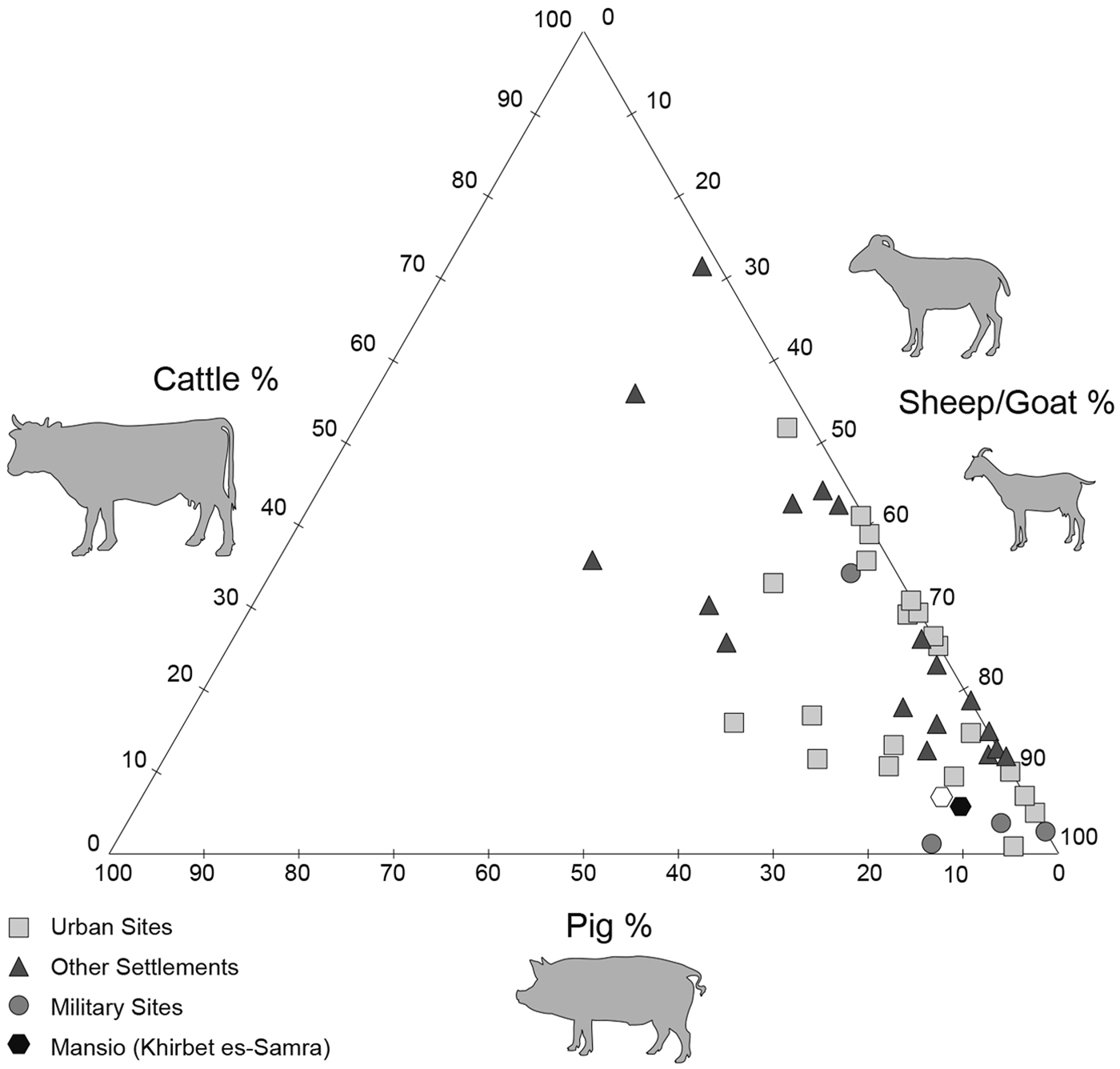
Fig. 4. Tri-plot showing relative frequencies of sheep/goat, cattle, and pig bones from the mansio assemblage and from many assemblages from the Eastern provinces (Syria, Palestine) of Hellenistic and Roman date, arranged by site type. (Drawing by M. Coutureau after King Reference King1999, table J.)
Caprines (sheep/goat)
With 747 remains, sheep and goat bones dominated the domestic species. Distinguishing between the bones of sheep and goats is a notorious challenge in zooarchaeology. Several methodological contributions have been published at different times to facilitate this task, largely relying on a macro-morphological approach.Footnote 19 However, despite the extensive literature, specific separation is not always easy, since a large majority of the criteria are not 100% reliable. In Samra it was not possible to clearly separate the two species, which were placed in a combined sheep/goat category, i.e., caprines.
Anatomical representation indicates that a broad range of skeletal elements were present at the site (Fig. 5). Apart from the small bones (i.e., phalanges, carpals, tarsals), none of the bones (i.e., skull, long bone, vertebrae, and ribs) were complete. Table 2 shows the skeletal profile for caprines where all the anatomical elements are present, suggesting that whole carcasses were processed. Thus for the total NISP (727), we observed 29.4% of elements belonging to the head, 11.8% to the axial skeleton, 17.9% to the forelimb, 24.2% to the hindlimb and finally 16.6% to the feet. It is important to consider these percentages as relative, since they are strongly related to bone fragmentation: for example, for the head, there are a large number of enamel fragments that increase the percentage of the head elements; for the axial skeleton, there are a large number of small fragments of rib which have been placed in the small-sized mammal class since they cannot be linked to a particular species. These reduce the percentage representation of the caprine axial skeleton. However, we can note that the forelimb (the shoulder joint, scapula–humerus–radius) and the hindlimb (pelvis–femur) are overrepresented. This, perhaps, indicates the regular practice of direct provisioning of the site with meat-rich joints of the shoulder (foreshanks) and upper leg.
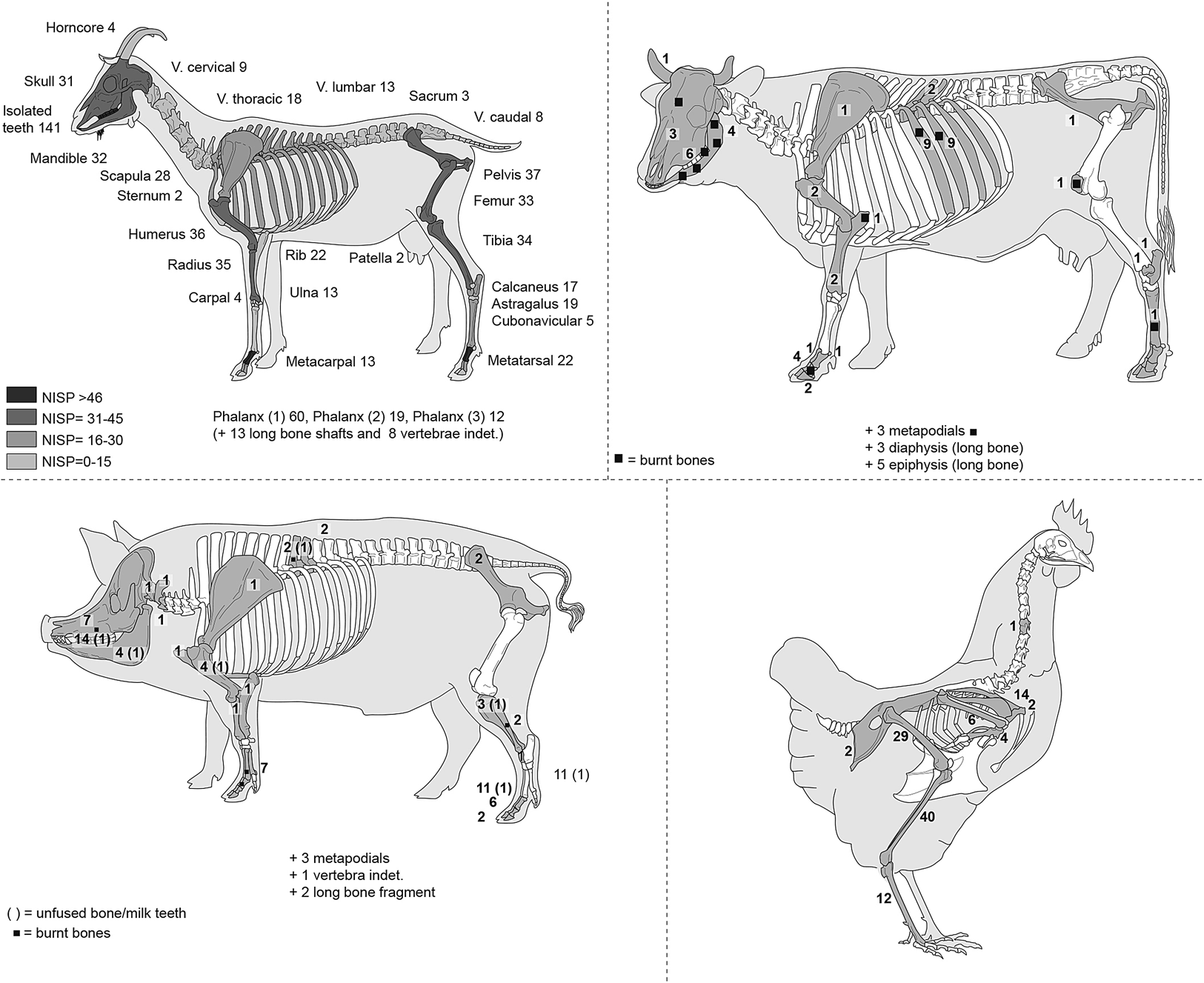
Fig. 5. Element representation in number of identified specimens (NISP) of caprines, cattle, pigs and chickens at the Samra mansio. (Drawing of skeletal profiles by M. Coutureau, available at https://www.archeozoo.org.)
Table 2. Anatomical representation of caprines, cattle, and pigs based on the number of identified specimens (NISP).
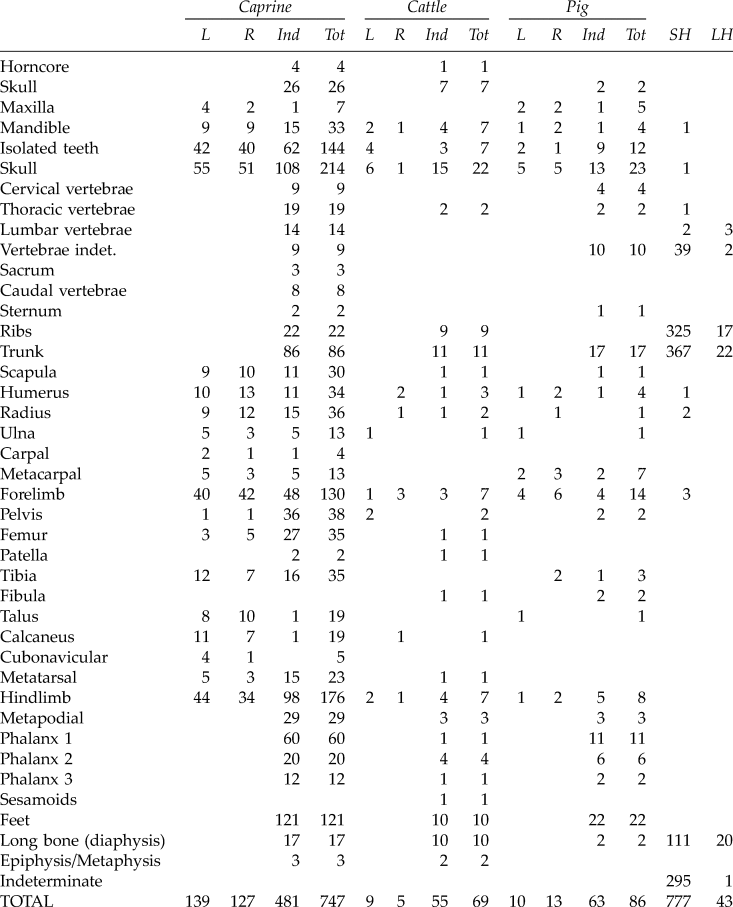
SH = small-size herbivore; LH = large-size herbivore; L = left; R = right; Ind = indeterminate; Tot = total
Twelve individuals (MNI) can be identified from the upper teeth, including nine adults identified from the right upper M1 and three juveniles identified from the right upper D4, two of whom were less than one year old at death.Footnote 20 The poor state of conservation of the lower teeth does not allow us to refine the mortality profile; at most, the lower teeth permit us to isolate three juveniles aged 6–12 months (Payne stage C), two young adults aged 2–3 years (Payne stage E), one adult aged 3–6 years (Payne stages F and G), and one old adult aged 8–10 years (Payne Stage I). Nor do the long bones’ epiphyseal fusion assist in refining the mortality ages; at most we can say that there are at least eight individuals aged over 2–3 years, including three individuals aged over 4–5 years.
Only five bones show butchery traces: three show fine incisions left by a knife – on a distal humerus, a first phalanx, and a coxal bone – while the two others show cleaver marks – on a sternum and a scapula. These indicate basic butchering operations, such as skinning, disarticulation, or filleting. On the other hand, all the skeletal parts bear traces of calcination (n = 73). No bone pathologies were identified in the caprine assemblage, as for the other species.
Pig (Sus scrofa domesticus)
Based on identified bone counts, pigs are the second-most abundant mammal on the site (n = 86), but pork does not seem to have played a major role in the food of the Roman occupants. It is presumed that the pigs at Samra were domestic. Anatomical representation demonstrates that a wide range of body parts were represented at the site, suggesting that complete animals were introduced into the building enclosure (Table 2 and Fig. 5). The slight overrepresentation of cranial (26.7%) and lower limb (25.6%) parts can be explained by the fact that they are more robust elements and so survive better. However, they can also represent anatomical regions which were discarded during the butchering because they are less nutritious. Nevertheless, the sample is too small to be able to identify butchering techniques. A similar pattern of skeletal representation is observed at many Roman-Byzantine sites where pig bones are present. No fully intact mandibles of pigs were encountered in our sample, and the state of loose tooth preservation was generally poor. The dental data that permitted the application of a wear-based aging technique, such as that of Horard-Herbin, which was used here, show the presence of a minimum of five individuals, including three over two years old, an individual about one year old, and a juvenile less than five or six months old.Footnote 21 Deciduous teeth associated with unfused long bones (i.e., humerus, fibula, tibia, and metapodial) indicate the presence of juveniles, testifying to the slaughter of immature individuals for their meat, confirming that pigs were animals exclusively raised for butchery.
Pigs may have been exploited at times of high demand in the seasonal supply of goat meat, or simply to add variety to the diet. Unlike goats and cattle, pigs do not produce any secondary products and are raised only for meat. In addition, they breed much faster than other domestic animals, and, although they do not eat grass, they can be fed on many other things, including waste. The low number of pigs can be explained by environmental conditions in the region, which are not conducive to intensive farming.Footnote 22
Cattle (Bos taurus)
In the ancient Near East, cattle were exploited for two main reasons: most importantly they provided traction for plowing the fields, and secondarily, they were a source of meat. Although cows are associated with milk production in the Western world, this was not their primary usage in the Near East.Footnote 23 Only small numbers of cattle remains (n = 69) occurred in the Roman layers at the site, which confirms that the consumption of cattle, as for pigs, was more occasional than that of caprines or poultry (Table 2 and Fig. 5). The bulk of the cattle bones belong to the head and extremities, i.e., primary butchery waste. The MNI count is three animals, including a calf. At Samra, the small scale of bovine management may be attributed to the scarcity of suitable pasture for cattle.
Chicken (Gallus gallus DOMESTICUS)
A total of 145 remains were determined with certainty as belonging to chickens, while 48 other bird remains that are more fragmented – mostly fragments of long bone – can likely be considered to belong to this species. There is a strong overlap between the small size of the domestic chicken from the premodern period and wild galliformes such as chukar partridge (Alectoris chukar), sand partridge (Ammoperdix heyi), and black francolin (Francolinus francolinus), all natives of southwest Asia. These birds have been identified in particular in levels at Upper Zohar and Tell Hesban.Footnote 24
The skeletal profile for chickens is given in Figure 5 and represents a minimum number of 12, estimated from the right distal end of the tibiotarsus. Among these individuals, there is at least one male individual (cock), indicated by the presence of a spur.
No cut marks or burning traces were observed, and the only bone modification observed consists of occasional damage to articular ends consistent with gnawing by humans, as seen for instance in Upper Zohar.Footnote 25 This can be compared to the known Roman cooking practices (from Apicius), with recipes for whole birds.Footnote 26 Although it is not uncommon to find eggshell fragments at archaeological sites in the Levant (e.g., in Petra and in Gaza), none were identified in the mansio.Footnote 27 The first archaeological evidence for chicken eggs in the Mediterranean is from the 1st c. BCE.Footnote 28 This evidence includes a high percentage of medullary bones, indicative of females during egg-laying.Footnote 29
Domestic species that could have been consumed
Camel (Camelus dromedarius)
It is not surprising to find camel bones (n = 26), as this species played an important role in the human exploitation of the desert, thanks to its exceptional natural ability to tolerate arid environments. Indeed, the dromedary was a beast of burden and played a key role in the Classical period economy at caravan centers, such as Petra in Jordan and Dumat al-Jandal or Al-Yamama in Saudi Arabia.Footnote 30 The skeletal profile for dromedary at Samra is limited and the entire skeleton is not represented: 1 cervical vertebra fragment, 1 scapula fragment, 1 phalange 1, 4 phalanges 2, 5 metapodial fragments, and 13 long bone shafts. The presence of one rib fragment exhibiting a cut mark does not allow us to know with certainty whether this species was consumed.
Equid (Equus sp.)
There are 16 bones that could be attributed to equids. As with the camel specimens, equids are mainly represented by isolated teeth (including a lower deciduous tooth) or tooth fragments (n = 12), phalanges (n = 3), and one left complete metacarpal 3. It is usually difficult to determine precisely whether these isolated equid remains belonged to a horse (Equus caballus), donkey (Equus asinus), or mule, but the small size of the third metacarpal suggests that it belonged to a donkey or a mule.Footnote 31 All these bones derive from the feet or skull: i.e., non-meaty bones. It is impossible to know whether equids were ever eaten in the mansio. The donkey remains represent animals that were probably used as beasts of burden to transport humans and goods.
Dog (Canis familiaris)
Eight bones (1 atlas, 1 lumbar vertebra, 1 proximal metatarsal, 3 phalanges 1, and 2 right distal humerus) were identified as belonging to two adult dogs. These bones are mixed in with those of other species. Though never abundant, dogs are often mentioned in zooarchaeological lists from Classical-period sites in the region.Footnote 32 They may have been used for herding, hunting, or as companions, or they may have been strays. Indeed, it is not uncommon nowadays to observe threatening packs of dogs in villages, feeding on garbage.Footnote 33
The wild species
Gazelle (Gazella sp.)
The Roman level of the mansio contained seven remains belonging to gazelle: 3 phalanx 1, 2 phalanges 2, 1 left talus, and 1 left metatarsal. All of these elements belong to the feet, i.e., a body part very poor in meat. They correspond to leftovers from slaughter, or even from cooking. They also indicate that the occupants of the mansio engaged in hunting. Never abundant, remains of gazelle are nevertheless present in many Roman-Byzantine sites in the Levant.Footnote 34
In the absence of specific elements such as horncore, it is not easy to determine the species. According to current data and the faunal assemblages of Near Eastern archaeological sites, three gazelle species inhabited the region: the mountain gazelle (Gazella gazella) is currently the most common in the region; the Persian gazelle (Gazella subgutturosa) occurs mainly at Mesopotamian sites and more to the east, but it is also present on the Jordanian plateau; and the Dorcas gazelle (Gazella dorcas) occupies a desert environment and is found today in Sinai and the Negev region bordering the Dead Sea.Footnote 35 Nevertheless, some measurements taken on these bone remains suggest individuals of small sizes close to the Dorcas gazelle.
Cape hare (Lepus capensis)
The Cape hare is widely distributed throughout the region in the present day and can occur in a wide variety of habitats such as steppes, agricultural areas, and even the desert, if there is sufficient vegetation.Footnote 36 In Samra, 15 fragments were identified in the Roman layers. Among them, a distal humerus is burned, the result of direct contact with the flame, the rest being protected by the flesh. It can be assumed that the Roman-period inhabitants of the mansio roasted the hare after cutting and removing the legs and the forearms by breaking, as is done today with this species.
Ostrich (Struthio camelus syriacus)
The Middle Eastern ostrich or Arabian ostrich is an extinct ostrich subspecies which once inhabited the Arabian peninsula and the Near East. In the mansio this species is represented by three eggshell fragments. While ostrich bones are quasi-absent at Near Eastern archaeological sites, with the exception of the site of Tell Hesban in Jordan, its eggshell fragments are more or less frequently found.Footnote 37 Ostriches were appreciated not only for the quality of their large eggs, their flesh, and their useful fat, but also for their feathers, especially those of the tail.Footnote 38 The presence of ostrich eggs is interesting, although it is difficult to know whether the eggs were consumed or used as containers.Footnote 39
Fish and seashells
The ichthyological material of Samra is very poor and consists of only four remains belonging to three families: Scombridae are represented by a single vertebra of the genus Euthynnus sp. (a small tuna), a marine species which came from the Red Sea; Cichlidae (tilapia), a freshwater fish, is represented by a ventral spine and a supracleithrum; Cyprinidae, the carp family, are represented by a precaudal vertebra whose genus attribution cannot be specified. Although their remains are rare, fish would also have played a big role in the diet, but the poor post-depositional condition of bone conservation (e.g., from weathering and trampling), associated with lack of sieving, may explain the low representation of fish remains, which are nevertheless abundant on some Roman sites in the region.Footnote 40
In the assemblage, 12 seashells belonging to five species were also identified (Table 1). Their presence suggests that they had probably been collected on purpose because of their natural shape, in order to be used as ornaments.
The case of burning bone
The mansio bone assemblage shows a large number of burned bones (n = 376, 7.3% of NISP) ranging from gray (scorched) to black (carbonized), but with few white bones indicative of intense calcination. These charred bones were found overwhelmingly in the central courtyard (70.5%) and to a lesser extent in the entrance (5.3%) and the vestibule (4.3%). Only the species consumed were affected by calcination: 73 bones of sheep/goat, 16 of cattle, 7 of pigs, 9 of chicken, 3 of gazelle, and 1 of hare. The remainder are indeterminate splinters. Finally, in the case of caprines, it should be noted that burning affected all anatomical parts, including many mandibles and teeth, the latter having certainly exploded into several enamel fragments after being exposed to fire.
Discussion
The bone remains were found in all parts of the building and mainly represent food leftovers linked to human activity (i.e., preparation of the carcass after slaughter and meal preparation) and date from the last occupation of the site. They are especially common in the courtyard (Locus 3) (27.1%), followed by the vestibule (Locus 2) (13.6%) (Fig. 6). This is not surprising if we assume that it was in this inner courtyard that the main cooking activities and food preparation took place. Furthermore, spatial localization does not show any differences for the four major species consumed (Fig. 6).
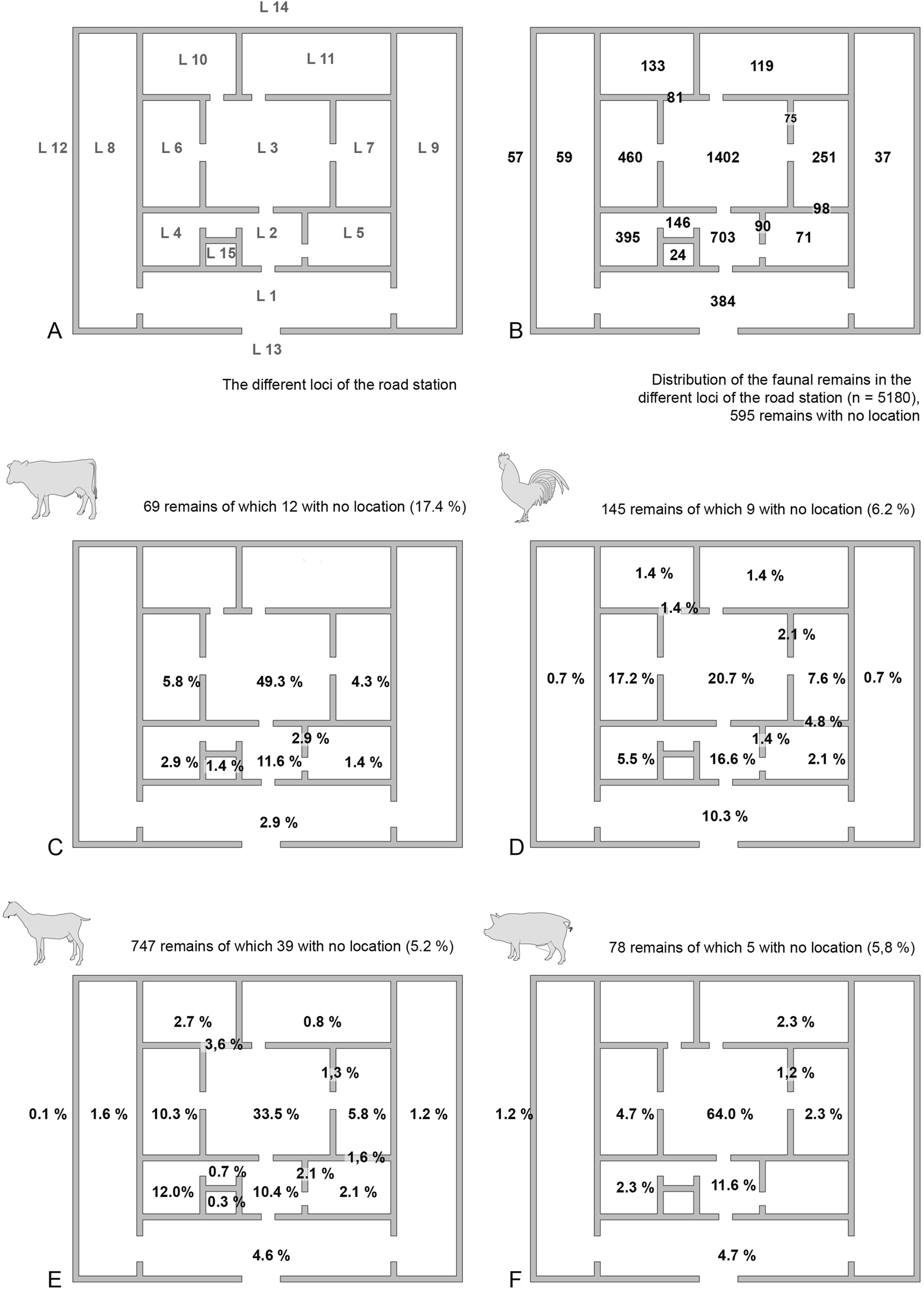
Fig. 6. Spatial distribution of the faunal remains in the different loci of the road station. A: loci numbering; B: distribution of all bone remains; distribution (%) of NISP of cattle (C), chicken (D), caprine (E), and pig (F).
Whole animals may have been brought to the site for subsequent slaughter and consumption. Most caprines were slaughtered at between two and three years of age, at which point they had reached their prime meat weight. This finding is in keeping with a subsistence strategy that stressed the exploitation of locally herded sheep and goats for meat production.
The overrepresentation of certain skeletal elements is notable. In the case of the caprines, it seems that the upper parts of the forelimb (the shoulder joint) were a favored part. This is very interesting, as a similar phenomenon was remarked for caprines in the early Byzantine fort of Upper Zohar, at the Byzantine monastery of Deir ‘Ain ‘Abata in Jordan, and at the Islamic site of Shahr-e Gholgholah in Afghanistan.Footnote 41 Goats are important not only for providing meat but also for secondary products such as milk and wool. The reason for preferring goats to sheep is quite clear, as sheep yield about half of the annual milk of goats and lactate for about three months, while goats can be milked for up to five months.Footnote 42
In contrast to the multipurpose production capacities of sheep, goats, and cattle, domestic pigs are valued for the production of a single commodity – meat. However, in many respects pigs rank far above other farmyard domesticates, as they furnish a higher meat yield, possess flesh of higher fatty and caloric content, yield greater numbers of young per birth, and attain harvest age at a faster rate.Footnote 43 It is known from texts that pork was the preferred meat of ancient Rome, at far as can be inferred from the wide range of pork recipes enumerated by the 1st-c. epicure Apicius, the attention devoted to swine breeding in Varro (Rust. 4), and the explicit phrasing of the Theodosian Code (Cod. Theod. 14.3, 14.4, 14.7).Footnote 44 Concerning the Limes Arabicus, it seems that the physical conditions necessary for swine raising appear to have been met from the Early Roman period onwards. Nevertheless, this area presents fewer pig remains than the more humid northern Mediterranean area, which is undoubtedly linked to the physiological needs of this species. Indeed, the Mediterranean region is more suitable for pig farming than the arid eastern regions, despite the increased rainfall and improved hydraulic technology observed in Roman times.Footnote 45
In the Roman period, meat – most often of poor quality – was usually cut into pieces and then cooked in a pot, which was also used to make broth (elixum) and was accompanied by cereal (barley, wheat, etc.) or vegetable (pea, lentil, vetch, etc.) porridge. In fact, it was a rather highly spiced preparation whose prophylactic virtues remedied the problems of conservation and freshness. The boiled preparation tenderized the old and tough meat. Sometimes the meat was grilled or roasted (assum).Footnote 46 Many bones in the Samra assemblage show traces of fire. It is not only small fragments that were affected by traces of calcination, but also many of the identified remains belonging to all species. It is difficult to explain all these burned bones as simply representing barbecued remains resulting from cooking. Indeed, roasting usually leaves only a few traces of charring on the bone, since the meat is rarely in contact with the flame. It seems more plausible that the burned remains are the result of cleaning activities, such as the periodic burning of rubbish, that took place to reduce the mass of organic debris produced by the occupants and visitors to the station. The excavations revealed pits or ash levels in several rooms, as for example in the courtyard (Locus 3). It is likely that beside the mansio there would have been a refuse heap which may have been collected for secondary use as fertilizer and taken to adjacent cultivated areas. Burned bone remains, just like the animal manure coming from the stables (Loci 8 and 9), would have served as suitable waste for manuring.
Conclusion
The faunal remains from the mansio of Samra have provided valuable insights into the life and economy of a stopover on the Via Nova Traiana. They represent one of the first datasets for this type of Roman building in the Near East. This zooarchaeological study confirms the food-related origin of these bone remains. The food items available to the resident and visiting community were very varied, ranging from caprine (mutton and goat's meat) to chicken, with some pork and beef. Although there is no evidence for dairy products associated with caprines and cattle, it is possible that they were consumed.
Hunted game like gazelle and hare were very occasionally consumed and probably provided an element of variety to the daily diet. The presence of tuna attests to its transport – certainly dried or salted – together with marine shells imported from the Red Sea. Both are a good indicator of the commercial activity that reigned along the Via Nova Traiana. Salting or preparation of fish products in brine (salsamenta or fish sauces such as garum or allec) allowed longer storage.Footnote 47 Although widely attested in other Roman sites in the region, the consumption of beasts of burden such as donkey or camel cannot be clearly established at the site.
The system of food procurement during the Roman period was probably complex. The availability of animals in the countryside was certainly the determining factor in the choice of menu. Indeed, while some goods were traded over long distances, others were only traded over short distances from local rural communities.Footnote 48 Future work will concentrate on understanding the village of Samra, which yielded numerous faunal remains. During the Roman period, a fortress stood on this site and was connected to the mansio; during the Byzantine period, a village grew up around the fortress. The other area of interest for future study is that the village of Samra presents a long occupation, from the Roman to the Ottoman period, and will provide valuable insights into the evolution of the environment and the economy in a semi-desert context in northern Jordan.
Acknowledgments
Thanks to Jean-Baptiste Humbert (EBAF, Jerusalem) and Alain Desreumaux (CNRS, Paris), directors of the archaeological mission of Samra, for entrusting me with the study of this faunal material. Also thanks to Wim Van Neer (Brussels Museum) for the fish identifications, to Michel Coutureau (INRAP) for the drawings, to Liora K. Horwitz (Hebrew University) for judicious comments, and finally to Manon Saenko, Louis de Lisle, and Simon Brelaud.
Supplementary Materials
The Supplementary Materials contain the following tables. Table A: number of the different species and indeterminate remains in the different loci of the road station. Table B: percentage of the different species and indeterminate remains in the different loci of the road station. Table C: caprine bone measurements (mm). Table D: cattle bone measurements (mm). Table E: camel bone measurements (mm). Table F: gazelle bone measurements (mm). Table G: Cape hare bone measurements (mm). To view the Supplementary Materials for this article, please visit https://doi.org/10.1017/S1047759422000034.











 Open access
Open access