



Introduction
Known as the earliest large-bodied, nonmammalian, probainognathian cynodonts discovered in South America, Chiniquodontidae is a clade characterized by the long secondary bony palate extending back to or beyond the posterior end of the tooth row, posterior sectorial postcanines with backwardly recurved main cusp, and the long pterygoid processes ending in thin projections (Huene, Reference Huene1936; Romer, Reference Romer1969a, Reference Romerb; Abdala and Giannini, Reference Abdala and Giannini2002). The clade is represented in the Middle to Late Triassic of the Santa Maria Supersequence, southern Brazil (Huene, Reference Huene1936; Romer, Reference Romer1969a; Sá-Teixeira, Reference Sá-Teixeira1982; Martinelli et al., Reference Martinelli, Soares and Schwanke2016, Reference Martinelli, Kammerer, Melo, Neto, Ribeiro, Da-Rosa, Schultz and Soares2017; Schmitt et al., Reference Schmitt, Martinelli, Melo and Soares2019), and the Chañares and Ischigualasto formations, western Argentina (Romer, Reference Romer1969b, Reference Romer1973; Martínez and Forster, Reference Martínez and Forster1996). More recently, the range of the family was extended to Africa, with the description of new specimens and the discovery of two species in Madagascar and Namibia (Abdala and Smith, Reference Abdala and Smith2009; Kammerer et al., Reference Kammerer, Flynn, Ranivoharimanana and Wyss2010; Mocke et al., Reference Mocke, Gaetano and Abdala2020). Consequently, the chiniquodontids have long geochronological and wide geographical distributions, strengthening biostratigraphic correlations among South American and African basins.
However, due to poor preservation and incompleteness of the first-discovered chiniquodontid specimens, diagnosing individual species has proven difficult. Consequently, the family has undergone several revisions, exclusion of species, and synonymization of genera (Huene, Reference Huene1936; Crompton, Reference Crompton1955; Hopson and Kitching, Reference Hopson and Kitching1972; Hopson, Reference Hopson, Schultze and Trueb1990; Battail, Reference Battail1991; Abdala and Giannini, Reference Abdala and Giannini2002; Martinelli et al., Reference Martinelli, Kammerer, Melo, Neto, Ribeiro, Da-Rosa, Schultz and Soares2017). Considering this, the study and reanalysis of Chiniquodontidae is relevant, and the increase in the number of chiniquodontid specimens can be achieved with the study of the unprepared fossils already in collections and the use of new technologies, e.g., computed tomography (CT-scan).
The CT-scan is a nondestructive method that allows the accurate visualization of external structures, e.g., bony sutures, often not visible on the fossil due to mechanical or chemical preparations or coverings of rock matrix and sediment (Rowe et al., Reference Rowe, Macrini and Luo2011; Souto et al., Reference Souto, Murta-Fonseca, Machado, Lopes and Fernandes2019; Wallace et al., Reference Wallace, Martínez and Rowe2019; Kerber et al., Reference Kerber, Martinelli, Rodrigues, Ribeiro, Schultz and Soares2020; Benoit et al., Reference Benoit, Nxumalo, Norton, Fernandez, Gaetano, Rubidge and Abdala2022). In addition, CT-scan images allow the visualization of internal cavities and morphological structures, and the generation of high-quality three-dimensional models (e.g., Cnudde and Boone, Reference Cnudde and Boone2013; Cunningham et al., Reference Cunningham, Rahman, Lautenschlager, Rayfield and Donoghue2014), producing high-quality data for phylogenetic and functional analyses. Consequently, it is an important tool in the evaluation of new fossil specimens and the revision of already published material. Nevertheless, only two chiniquodontid specimens have CT-scan data available: MCP 1600 PV, the holotype of ‘Probelesodon’ kitchingi Sá-Teixeira, Reference Sá-Teixeira1982 (present study) and PVSJ 411, the holotype of ‘Probelesodon’ sanjuanensis Martínez and Forster, Reference Martínez and Forster1996 (see Rowe and Sill, Reference Rowe and Sill2001).
The present paper provides new information about the morphology of one of the best- preserved chiniquodontid specimens (MCP 1600 PV, ‘Probelesodon’ kitchingi holotype), based on three-dimensional (3D) models obtained through CT-scan images. We redescribe the specimen, revising the known morphological information and improving available data. In addition, the taxonomic status of the specimen is discussed, as well as the synonymization of species within the family.
Chiniquodontidae—background
Chiniquodon theotonicus Huene, Reference Huene1936 and ‘Belesodon magnificus’ Huene, Reference Huene1936 were the first described chiniquodontids, based on two sets of incomplete and poorly preserved specimens (one for each species), collected at the locality of Chiniquá, Rio Grande do Sul state, Brazil. The holotype of Chiniquodon theotonicus (GPIT 1050) is an incomplete skull with preserved sectorial postcanines and an associated humerus (Huene, Reference Huene1936; Romer, Reference Romer1969a). In contrast, ‘Belesodon magnificus’ was based on a large and nearly complete, but very poorly preserved skull (GPIT 40, 26 cm in basal skull length), with associated postcranial materials (Huene, Reference Huene1936). Subsequent expeditions in the region led to the discovery of new specimens of Chiniquodon theotonicus including two skulls (MCZ 3614 and 3615), a pair of mandibles, and some postcranial elements, as well as one skull of ‘Belesodon magnificus’ (MCZ 1533), all collected near Candelária municipality, Rio Grande do Sul state, Brazil (Romer, Reference Romer1969a). However, they also presented numerous breaks on the bone surface, giving a ‘crackle-ware’ effect (Romer, Reference Romer1969b). Despite these new discoveries, the two species and the family lacked proper diagnoses due to poor preservation and deformation of the materials.
The situation changed with the description of ‘Probelesodon lewisi’ Romer, Reference Romer1969b based on nine well-preserved skulls of different sizes (basal skull length range 7.2–15.5 cm) and postcranial elements from the Chañares Formation (early Late Triassic), Argentina. Those materials provided new information about the morphology of Chiniquodontidae, but a formal diagnosis was first proposed by Hopson and Kitching (Reference Hopson and Kitching1972). The authors also included in the family the species Aleodon brachyrhamphus Crompton, Reference Crompton1955 from the Manda Beds of Tanzania, despite its divergent postcanine morphology in relation to the already described chiniquodontids, and its former description as a gomphodont (but see Martinelli et al., Reference Martinelli, Kammerer, Melo, Neto, Ribeiro, Da-Rosa, Schultz and Soares2017).
The composition of the family was further expanded with the description of ‘Probelesodon minor’ Romer, Reference Romer1973 from the Chañares Formation, ‘Probelesodon’ kitchingi from the middle-early Late Triassic of the Santa Maria Supersequence, Brazil, and ‘Probelesodon’ sanjuanensis from the lower half of the Ischigualasto Formation (Late Triassic), Argentina. In addition, Hopson (Reference Hopson, Schultze and Trueb1990) included Cromptodon mamiferoides Bonaparte, Reference Bonaparte1972 from the Cerro de las Cabras Formation of Argentina, and Battail (Reference Battail1991) added Cistecynodon parvus Brink and Kitching, Reference Brink and Kitching1953 from the Burgersdorp Formation of South Africa, and Prozostrodon brasiliensis Bonaparte and Barberena, Reference Bonaparte and Barberena2001 (first coined as Thrinaxodon brasiliensis Barberena, Bonaparte, and Teixeira, Reference Barberena, Bonaparte and Teixeira1987) from the Santa Maria Supersequence, Brazil. Finally, the range of the family was extended to Madagascar and Namibia with the description of Chiniquodon kalanoro Kammerer et al., Reference Kammerer, Flynn, Ranivoharimanana and Wyss2010 (‘Isalo II’ beds, early Carnian) and Chiniquodon omaruruensis Mocke, Gaetano, and Abdala, Reference Mocke, Gaetano and Abdala2020 (upper Omingonde Formation, possible early Carnian).
A formal diagnosis for Chiniquodon theotonicus was provided by Abdala (Reference Abdala1996), who revised the family and proposed that it is composed of only three species: ‘Belesodon magnificus,’ Chiniquodon theotonicus (including ‘Probelesodon lewisi’ and ‘Probelesodon minor’ as junior synonyms), and ‘Chiniquodon kitchingi’ (formerly ‘Probelesodon’ kitchingi). Subsequently, enhancing this study, Abdala and Giannini (Reference Abdala and Giannini2002) performed allometric and comparative analyses with the available sample of chiniquodontids from Argentina and Brazil, and concluded that all can be arranged into a skull growth series; therefore, the differences observed can be related to ontogenetic variation. As a result, ‘Belesodon magnificus’ (represented by a large skull), ‘Probelesodon lewisi,’ ‘Probelesodon minor,’ and ‘Probelesodon’ kitchingi were considered junior synonyms of Chiniquodon theotonicus. In addition, the authors recognized ‘Probelesodon’ sanjuanensis as a distinct species and proposed the new combination Chiniquodon sanjuanensis based on its highly arched zygomatic arch, with the ventral border strongly curved and extending to a level above the middle of the orbit, and the presence of a serrated posterior edge of the main cusp of at least the last postcanine.
Following Abdala and Giannini (Reference Abdala and Giannini2002), Chiniquodontidae is characterized as follows. Carnivorous eucynodonts with an overall cranial morphology similar to those of Thrinaxodon spp., but showing a more robust zygomatic arch; characteristic angulation (≥ 110°) between the ventral edge of the maxillary zygomatic process and the anteroventral margin of the jugal; enlarged pterygoid flanges (pterygoid processes) ending in a thin projection and directed posteroventrally; elongated secondary bony palate; and the posterior sectorial postcanines with backwardly recurved main cusps, lacking cingula or with tiny lingual cingular cusps. Based on this diagnosis, Cromptodon mamiferoides, Cistecynodon parvus, Prozostrodon brasiliensis, and Aleodon brachyrhamphus were excluded from Chiniquodontidae (Abdala and Giannini, Reference Abdala and Giannini2002).
Until recently, the composition of the family was restricted to Chiniquodon theotonicus, Chiniquodon sanjuanensis, and Chiniquodon kalanoro, but with the description of Aleodon cromptoni Martinelli et al., Reference Martinelli, Kammerer, Melo, Neto, Ribeiro, Da-Rosa, Schultz and Soares2017 from the Santa Maria Supersequence (Pinheiros-Chiniquá Sequence), the genus Aleodon Crompton, Reference Crompton1955 was referred to the family again. This inclusion is supported by phylogenetic analyses that recovered Chiniquodon theotonicus and Aleodon spp. as a monophyletic group (Hopson and Kitching, Reference Hopson and Kitching2001; Martinelli et al., Reference Martinelli, Kammerer, Melo, Neto, Ribeiro, Da-Rosa, Schultz and Soares2017). However, despite the presence of a main cusp highly recurved posteriorly, most postcanines of Aleodon sp. also bear a lingual platform (Martinelli et al., Reference Martinelli, Kammerer, Melo, Neto, Ribeiro, Da-Rosa, Schultz and Soares2017), a feature not included in the diagnosis stated by Abdala and Giannini (Reference Abdala and Giannini2002).
Geological setting
The specimen MCP 1600 PV was collected in a Triassic outcrop named Rincão do Pinhal, in the central region of Rio Grande do Sul state, southern Brazil (Fig. 1.1). The region corresponds to the Santa Maria Supersequence (SMS), and according to Horn et al. (Reference Horn, Melo, Schultz, Philipp, Kloss and Goldberg2014), this geological setting is composed of four third-order sequences, from base-to-top: (1) Pinheiros-Chiniquá Sequence, (2) Santa Cruz Sequence, (3) Candelária Sequence, and (4) Mata Sequence. The two lowermost sequences are composed of ephemeral fluvial (braided) deposits overlain by dry mudflat facies (loess deposits), whereas the Candelária Sequence is characterized by a sheet delta association (underwater deposits), indicating a change from arid to humid conditions (Horn et al., Reference Horn, Goldberg and Schultz2018; Corecco et al., Reference Corecco, Pereira, Soares and Schultz2020). The Mata Sequence is characterized by cross-stratified, conglomeratic sandstones of fluvial origin, with abundant silicified logs, from which fossil vertebrates are still unknown (Horn et al., Reference Horn, Melo, Schultz, Philipp, Kloss and Goldberg2014).

Figure 1. (1) Exposures of the rocks of the Santa Maria Supersequence and the outcrop where MCP 1600 PV was found; the outcrop is represented by the gray rectangle in the detail, near Paraíso do Sul municipality. (2) Chrono- and biostratigraphy of Triassic units with vertebrate Assemblage Zones (AZ) of the Santa Maria Supersequence. Modified from Martinelli et al. (Reference Martinelli, Kammerer, Melo, Neto, Ribeiro, Da-Rosa, Schultz and Soares2017). The chronostratigraphic dates are based on detrital zircons analyses performed by Langer et al. (Reference Langer, Ramezani and Da-Rosa2018) and Philipp et al. (Reference Philipp, Schultz, Kloss, Horn, Soares and Basei2018).
Four faunal associations for Middle–Late Triassic outcrops from southern Brazil were established in recent years and they can be correlated with Argentinian and African stratigraphic units (e.g., Soares et al., Reference Soares, Schultz and Horn2011; Horn et al., Reference Horn, Melo, Schultz, Philipp, Kloss and Goldberg2014; Martinelli et al., Reference Martinelli, Soares and Schwanke2016, Reference Martinelli, Kammerer, Melo, Neto, Ribeiro, Da-Rosa, Schultz and Soares2017, Reference Martinelli, Escobar, Francischini, Kerber, Müller, Rubert, Schultz and Da-Rosa2021; Schmitt et al., Reference Schmitt, Martinelli, Melo and Soares2019; Abdala et al., Reference Abdala, Gaetano, Martinelli, Soares, Hancox and Rubidge2020; Schultz et al., Reference Schultz, Martinelli, Soares, Pinheiro, Kerber, Horn, Pretto, Müller and Melo2020; Fig. 1.2). The Dinodontosaurus Assemblage Zone (AZ; Ladinian-early Carnian, Pinheiros-Chiniquá Sequence) is correlated, in part, with the tetrapod faunas of the Chañares Formation (see discussions by Martinelli et al., Reference Martinelli, Kammerer, Melo, Neto, Ribeiro, Da-Rosa, Schultz and Soares2017 and Ezcurra et al., Reference Ezcurra, Fiorelli, Martinelli, Rocher and von Baczko2017), upper Omingonde Formation from Namibia, and with the Isalo II beds from Madagascar. The Santacruzodon AZ (early Carnian, Santa Cruz Sequence) is the least taxonomically diverse AZ, but presents some faunal correlation with the Isalo II beds (late Ladinian–early Carnian) and similar absolute dates (see below) with the lower part of the Chañares Formation; it also shares some faunal components with the Dinodontosaurus AZ (Schmitt et al., Reference Schmitt, Martinelli, Melo and Soares2019). The Hyperodapedon AZ (Carnian, Candelária Sequence) fauna is correlated with the Ischigualasto Formation and the Riograndia AZ (early Norian, Candelária Sequence) is associated with the lower fauna of the Los Colorados Formation (Langer et al., Reference Langer, Ribeiro, Schultz and Ferigolo2007, Reference Langer, Ramezani and Da-Rosa2018; Desojo et al., Reference Desojo, Fiorelli, Ezcurra, Martinelli and Ramezani2020; Schultz et al., Reference Schultz, Martinelli, Soares, Pinheiro, Kerber, Horn, Pretto, Müller and Melo2020; Martinelli et al., Reference Martinelli, Escobar, Francischini, Kerber, Müller, Rubert, Schultz and Da-Rosa2021).
In addition, maximum absolute ages were obtained in some sites from the Rio Grande do Sul (Langer et al., Reference Langer, Ramezani and Da-Rosa2018; Philipp et al., Reference Philipp, Schultz, Kloss, Horn, Soares and Basei2018), indicating maximum depositional ages of 236.1 Myr to the Santacruzodon AZ (Santa Cruz do Sul and Venâncio Aires municipalities), 233.23 Myr to the Hyperodapedon AZ (Cerro da Alemoa site), and 225.42 Myr to the Riograndia AZ (Linha São Luiz site, Faxinal do Soturno municipality). The region where MCP 1600 PV was collected corresponds to the Dinodontosaurus AZ of the Pinheiros-Chiniquá Sequence (Santa Maria Supersequence). It was found in the same outcrop (see Materials and methods) where the dicynodont Dinodontosaurus Romer, Reference Romer1943 occurred, and it is stratigraphically lower than the Santa Cruz Sequence and therefore has a maximum age of 236.1 Myr.
Materials and methods
Analyzed specimen
MCP 1600 PV (‘Probelesodon’ kitchingi holotype) is a well-preserved skull (Figs. 2–5) from the Rincão do Pinhal locality, Agudo municipality, Rio Grande do Sul state, Brazil (Fig. 1). The specimen was collected by the priest Abrahão Cargnin in 1977, and it was associated with disarticulated materials of distinct taxa (dicynodonts, other cynodonts, and archosauromorphs), reflecting a postmortem association (Sá-Teixeira, Reference Sá-Teixeira1982).

Figure 2. Chiniquodon kitchingi (Sá-Teixeira, Reference Sá-Teixeira1982) comb. nov. (MPC 1600 PV), skull in dorsal view: (1) photograph; (2) schematic drawing. The black arrow indicates the serrated suture between the frontal and nasal bones. fr = frontal; ju = jugal; la = lacrimal; ma = maxilla; na = nasal; pa = parietal; pma = premaxilla; ppf = pterygoparoccipital foramen; prf = prefrontal; po = postorbital; sm = septomaxilla; sq = squamosal. Scale bar = 5 cm.
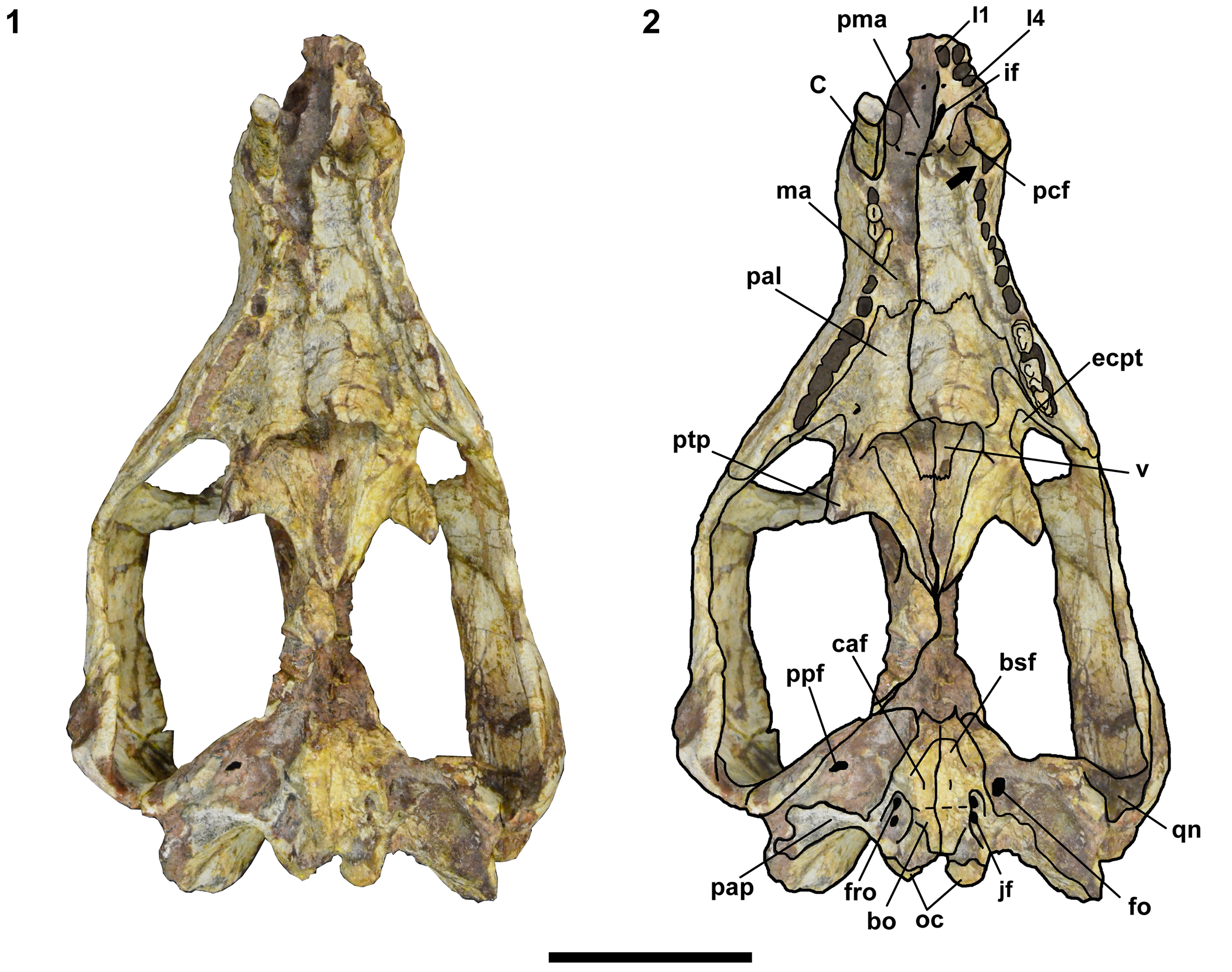
Figure 3. Chiniquodon kitchingi (Sá-Teixeira, Reference Sá-Teixeira1982) comb. nov. (MPC 1600 PV), skull in ventral view: (1) photograph; (2) schematic drawing. The black arrow indicates the space on the canine alveolus. bo = basioccipital; bsf = basisphenoid; C = upper canine; caf = carotid artery foramen; ecpt = ectopterygoid; fo = fenestra ovalis; fro = fenestra rotunda; I# = upper incisor; if = incisive foramen; jf = jugular foramen; ma = maxilla; oc = occipital condyle; pal = palatine; pap = paroccipital process; pcf = paracanine fossa; pma = premaxilla; ppf = pterygoparoccipital foramen; ptp = pterygoid process; qn = quadrate notch; v = vomer. Scale bar = 5 cm.

Figure 4. Chiniquodon kitchingi (Sá-Teixeira, Reference Sá-Teixeira1982) comb. nov. (MPC 1600 PV), skull in left lateral view: (1) photograph; (2) schematic drawing. The black arrow indicates the characteristic angulation between the jugal and maxillary bones of chiniquodontids. Abbreviations as in Figures. 2 and 3; plus ib = internarial bar; lbc = lambdoidal crest; pfr = prefrontal; pt = pterygoid. Scale bar = 5 cm.
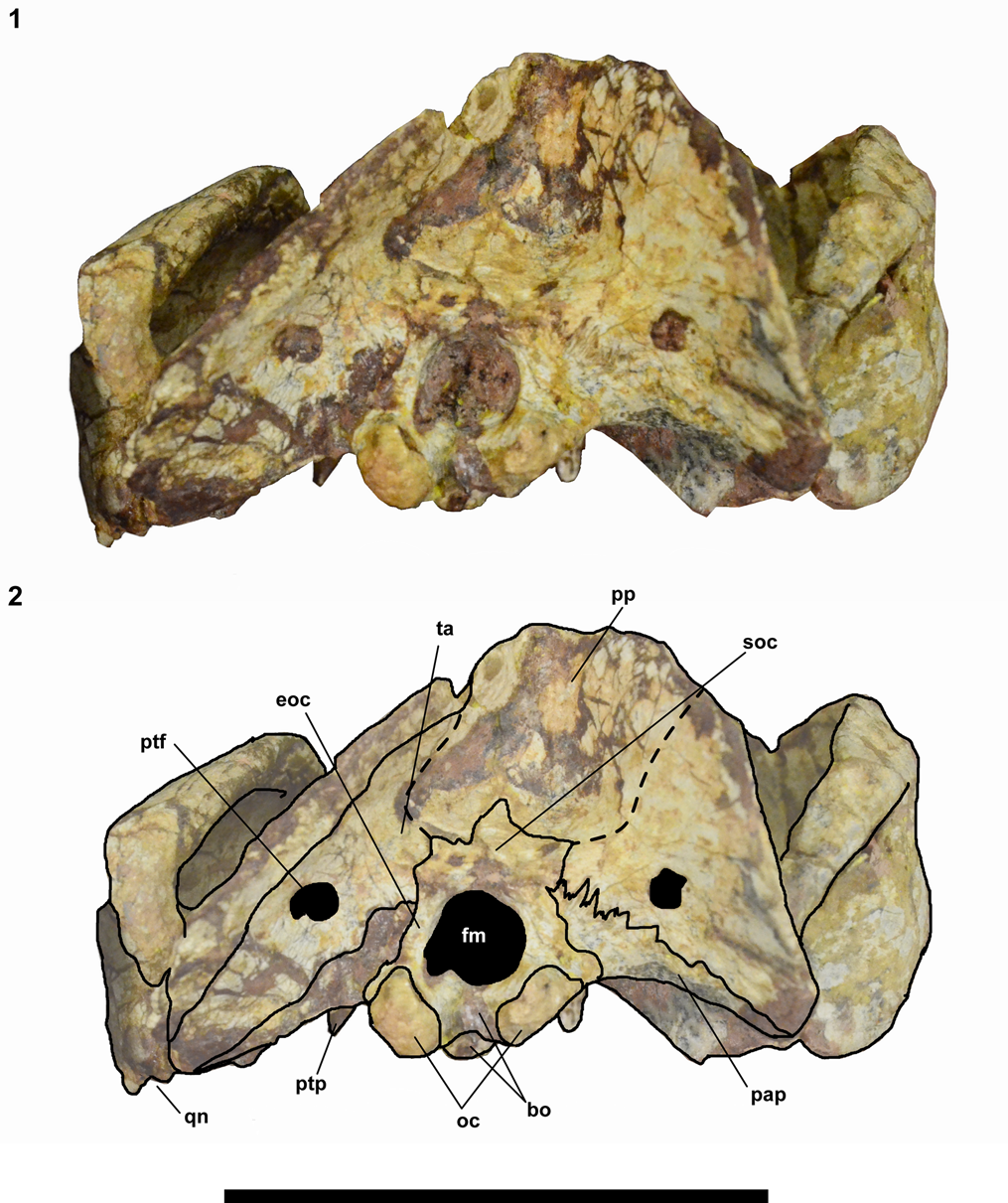
Figure 5. Chiniquodon kitchingi (Sá-Teixeira, Reference Sá-Teixeira1982) comb. nov. (MPC 1600 PV), skull in occipital view: (1) photograph; (2) schematic drawing. Abbreviations as in Figure 3; plus eoc = exoccipital; fm = foramen magnum; pp = postparietal; ptf = postparietal foramen; soc = supraoccipital; ta = tabular. Scale bar = 5 cm.
It was studied first-hand, and for comparative morphological analyses, we used descriptions, schematic drawings, and photography of the specimens available in the literature (e.g., Romer, Reference Romer1969a, Reference Romerb, Reference Romer1973; Martínez and Forster, Reference Martínez and Forster1996; Abdala and Giannini, Reference Abdala and Giannini2002; Martinelli et al., Reference Martinelli, Kammerer, Melo, Neto, Ribeiro, Da-Rosa, Schultz and Soares2017). Some specimens from Argentina and Brazil were studied first-hand by AGM (e.g., PVL 4167, 4444, 4674, 4675, PVSJ 411, MCZ 3614, 3615, UFRGS-PV-1331-T, and NHM-UK PV R8429).
A total of 22 skull measurements were taken directly from the specimen MCP 1600 PV with the aid of a caliper (Fig. 6). The measurement parameters were taken from Sá-Teixeira (Reference Sá-Teixeira1982; eight measurements) and Abdala and Giannini (Reference Abdala and Giannini2002; seven measurements). We also added seven measurements: orbital diameter (OD), orbital opening height (OH), temporal fenestra length (TFL), the distance between the pterygoid processes (PTPW), maximum snout height (SH), and pterygoid transverse processes length (PTPL). The source of each skull measurement parameter is summarized in Table 1.

Figure 6. Chiniquodontidae, skull measurement parameters. Modified from Abdala and Giannini (Reference Abdala and Giannini2002). See text for abbreviations.
Table 1. Measurements (in cm) of the skull of MCP 1600 PV Chiniquodon kitchingi (Sá-Teixeira, Reference Sá-Teixeira1982) comb. nov., holotype. * = measurement parameters from Abdala and Giannini (Reference Abdala and Giannini2002); ** = taken from Sá-Teixeira (Reference Sá-Teixeira1982).

Computed tomography
The CT-scan was performed at the Instituto do Cérebro, São Lucas Hospital (PUCRS, Brazil), using the medical scanner Pet CT Multislice 16d Discovery. The fossil was scanned with a resolution of 512 x 512 pixels, 0.625 mm between slices, 140 kV and 380 mA, resulting in 676 slices. Fossils from the same outcrop and region usually contain a high concentration of calcite (e.g., Holz and Schultz, Reference Holz and Schultz1998; Pavanatto et al., Reference Pavanatto, Da-Rosa, Müller, Roberto-Da-Silva, Ribeiro, Martinelli and Dias-Da-Silva2020). This mineral interacts with X-rays, resulting in artifacts on CT-scan images, which make it difficult to accurately visualize internal and external structures (Cnudde and Boone, Reference Cnudde and Boone2013). To try to mitigate possible artifacts, the protocol of Hoffmann et al. (Reference Hoffmann, Rodrigues, Soares and Andrade2021) was followed.
Visualization of the CT-scan images, segmentation of each bone, and generation of the 3D models were performed using the software Avizo 7.1 Standard Edition (Thermo Fisher Scientific, 2012). The gray values of the images were altered to better visualize the density differences between rock matrix and bone elements. Bone segmentation was performed using the ‘segmentation editor.’ Each bone was manually selected with a different color, using the coronal view. In addition, three videos of the 3D models were generated (see Supplementary Data 1–3).
Repositories and institutional abbreviations
GPIT, Institut und Museum für Geologie und Paläontologie der Universität, Tübingen, Germany; GSN, Geological Survey of Namibia, Windhoek, Namibia; MCN PV, Museu de Ciências Naturais, Coleção de Paleovertebrados, Porto Alegre, RS, Brazil; MCP, Coleção de Fósseis, Museu de Ciências e Tecnologia, Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre, Brazil; MCZ, Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts, USA; NHM-UK, The Natural History Museum, London, UK; PUCRS, Pontifícia Universidade Católica do Rio Grande do Sul, Porto Alegre, Brazil; PULR, Museo de Antropología, Universidad Nacional de La Rioja, Argentina; PVL, Colección Paleontología de Vertebrados Lillo, Universidad Nacional de Tucumán, Argentina; PVSJ, Instituto y Museo de Ciencias Naturales, San Juan, Argentina; UFRGS-PV, Paleovertebrate Collection, Universidade Federal do Rio Grande do Sul, Brazil.
Systematic paleontology
Probainognathia Hopson, Reference Hopson, Schultze and Trueb1990
Family Chiniquodontidae Huene, Reference Huene1936
Genus Chiniquodon Huene, Reference Huene1936
Type species
Chiniquodon theotonicus Huene, Reference Huene1936.
Diagnosis
Probainognathian cynodonts with an elongated secondary bony palate, with the posterior palatine lateral projections reaching beyond the level of the tooth row; elongated basicranial ventral bar; horizontal S-shaped and concave-convex suture between the jugal and squamosal on the zygomatic arch; long pterygoid transverse process; sectorial postcanines with main cusps recurved posteriorly, lacking a lingual platform and mesial and distal denticulated margins; distal region of the humerus posteromedially expanded (modified from Abdala, Reference Abdala1996 and Abdala and Giannini, Reference Abdala and Giannini2002).
Chiniquodon theotonicus Huene, Reference Huene1936
Holotype
GPIT 1050, partial skull and humerus.
Diagnosis
Chiniquodontid cynodont with a stout skull and a short and wide snout in relation to the total skull length; a zygomatic arch with a slightly arched ventral border, reaching the level of the ventral orbital edge; sectorial postcanines having no, or very reduced, cingula, and lacking mesial and distal denticulated cutting edges. Modified from Abdala and Giannini (Reference Abdala and Giannini2002).
Occurrence
Brazilian specimens are known from several outcrops in Rio Grande do Sul state, discovered in rocks of the Dinodontosaurus AZ (late Ladinian to early Carnian), Pinheiros-Chiniquá Sequence, Santa Maria Supersequence. Argentinian specimens come from La Rioja Province, Argentina, from the Massetognathus-Chanaresuchus AZ of the Chañares Formation (early Carnian).
Referred material
GPIT 40; UFGRS-PV-66-T, 275-T, 1331-T; MCN PV 2755; MCZ 1533, 3035, 3614, 3615, 3776, 3777, 3779, 3781, 4002, 4020, 4100, 4296, 8823; PVL 4167, 4444, 4448, 4674, 4675; PULR 12, 18, 100, 101, 102; NHM-UK-R 8429.
Remarks
According to Abdala and Giannini (Reference Abdala and Giannini2002), two specimens could represent a different species: the skull MCZ 3615 (Romer, Reference Romer1969a) and the lower jaw MCN PV 2755 (Abdala et al., Reference Abdala, Ribeiro and Schultz2001) due to the presence of posterior postcanines with small lingual cingula, consisting of two tiny cusps. However, considering the poor preservation of the specimens and the possibility of variation in postcanine morphology within the same species, the authors decided to include them in Chiniquodon theotonicus until new materials are found. Also, this feature was reported in the specimens known from the Santacruzodon AZ (Abdala et al., Reference Abdala, Ribeiro and Schultz2001; Martinelli et al., Reference Martinelli, Ribeiro, Campo and Soares2018) so they might represent a different morphotype, with a broad biostratigraphic interval.
Chiniquodon kitchingi (Sá-Teixeira, Reference Sá-Teixeira1982) new combination
Figures 2–5, 7–9, 10.1

Figure 7. Colorized 3D model of the skull of Chiniquodon kitchingi (Sá-Teixeira, Reference Sá-Teixeira1982) comb. nov. with segmented bones: (1) ventral and (2) dorsal views. The black arrow indicates the small crest formed by the sutures between the frontals. Abbreviations as in Figures 2, 3, and 5; plus pr + op = prootic + opisthotic.
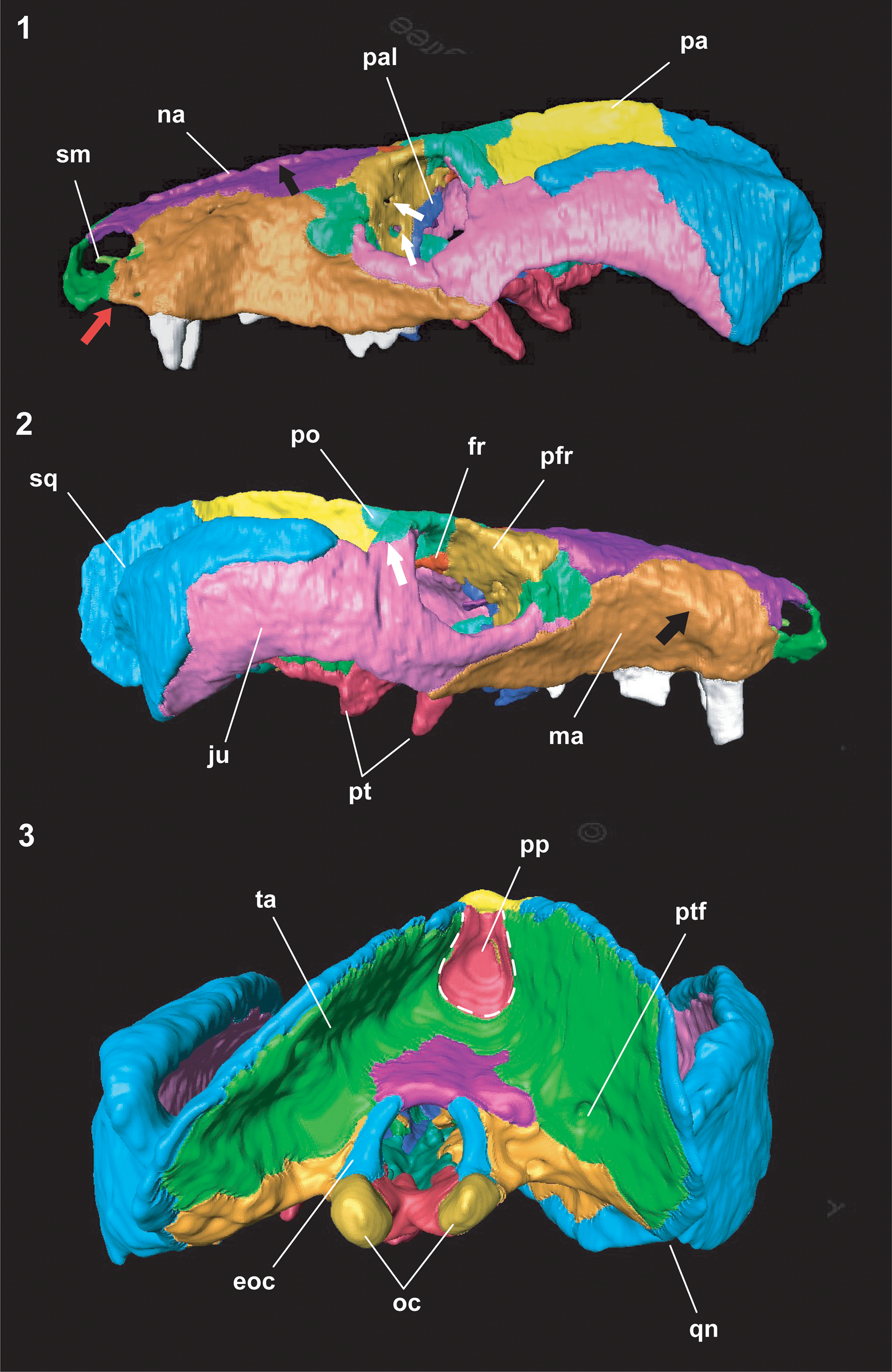
Figure 8. Colorized 3D model of the skull of Chiniquodon kitchingi (Sá-Teixeira, Reference Sá-Teixeira1982) comb. nov. with segmented bones: (1) left, (2) right, and (3) occipital views. The black arrow in (1) indicates the crest formed on the suture between the nasals; the two white arrows indicate the two foramina on the prefrontal bones, and the red arrow indicates the step on the maxilla. In (2), the white arrow points to the suture between the jugal and the postorbital, and the black arrow indicates the canine root crest. Abbreviations as in Figures 2–5.

Figure 9. Chiniquodon kitchingi (Sá-Teixeira, Reference Sá-Teixeira1982) comb. nov.: (1) colorized 3D model of the skull in right lateral view with jugal and squamosal excluded; (2) colorized 3D model and (3) photograph of the basicranium. Abbreviations as in Figures 2–5 and 7; plus ept = epipterygoid; io = interorbital opening; orb = orbitosphenoid; tri = trigeminal nerve foramen. Scale bar = 5 cm.

Figure 10. Comparison of selected skulls of Chiniquodon from South America: (1) skull of MCP 1600 PV (holotype of ‘Probelesodon kitchingi’) in lateral (reversed) and dorsal views; (2) skull and articulated jaws of PVL 4444 in lateral and dorsal views, from the Massetognathus-Chanaresuchus AZ of the Chañares Formation, Argentina; (3) skull and articulated jaws of UFRGS-PV-1331-T (ex UFRGS-PV-0066-Tg) in lateral and dorsal views, from the Dinodontosaurus AZ of Brazil. Scale bars = 2 cm.
Holotype
MCP 1600 PV, skull with dentition.
Diagnosis
Chiniquodontid cynodont with a more gracile general skull shape (longer and narrower snout) in comparison to Chiniquodon theotonicus (see Fig. 10); ‘arrow’-shaped frontals with lateral expansions toward the prefrontals; ectopterygoid not in contact with maxilla; rounded snout tip; medial contact between the posterior rami of the postorbitals; and a highly angled suture (nearly 90°) between the jugal and squamosal bones.
Occurrence
Rincão do Pinhal locality, Agudo municipality, Rio Grande do Sul state, Brazil; Dinodontosaurus AZ (late Ladinian to early Carnian), Pinheiros-Chiniquá Sequence, Santa Maria Supersequence.
Description
Premaxilla.—Most of the right premaxilla is broken, and the left one is almost complete. The external nasal openings are clearly separated by a thin internarial bar formed by both premaxillae. The extranasal process (i.e., the dorsal facial process) is short and does not contact the nasal. The facial suture between the premaxilla and maxilla is not clear on both sides of the skull, but it was possible to infer its position based on the CT-data (Fig. 8.1). Only the left incisive foramen is preserved (Fig. 3). It is long and narrow, separated from the right one, in its anterior portion, by a thin lamina of bone (Fig. 3). The incisive foramen is apparently enclosed within the premaxilla, with no participation of the maxilla. In ventral view, the two premaxillae form a ‘pointed arch,’ where the incisors are located. There is no evidence of the pointed prominence formed by the anterior portions of the premaxillae described for Chiniquodon theotonicus specimens (Romer, Reference Romer1969a, Reference Romerb): in MCP 1600 PV, the snout is round in lateral view (Fig. 4). Both paracanine fossae are present, anteromedially to the canines, and posteriorly delimited by the maxillae (Fig. 3).
Septomaxilla.—The left septomaxilla is preserved as a curved and slender bone inside the nasal opening cavity (Fig. 4). The facial process is short and does not contact the nasals dorsally.
Maxilla.—The maxillae have a roughly triangular shape in lateral view and form the greater part of the lateral surface of the snout (Fig. 4). Delimitation of the maxillae by the nasals is evident, starting on the posterior border of the nasal openings, diverging on the widest part of the nasals at ~90° downward, and reaching the lacrimal and jugal posteriorly.
The snout presents an expansion close to the nasal cavities due to the enlarged canine roots (visible in lateral and dorsal views; Figs. 4, 7.2). A step is present on the ventral margin of the maxillae between the last incisors and the canine (Figs. 4, 8.1). This feature is also present in some Chiniquodon theotonicus specimens from the Chañares Formation (Argentina) and Chiniquodon omaruruensis (see Mocke et al., Reference Mocke, Gaetano and Abdala2020). Right below the posterior border of the orbit, the ventral margin of the maxillae contacts the jugals, forming a suborbital angulation of ~120° (Fig. 4), which is one of the synapomorphies of chiniquodontids (Abdala and Giannini, Reference Abdala and Giannini2002).
At least three discrete foramina (Fig. 4) are discerned on the lateral surface of the maxillary bones, which represent the maxillary nerve external terminations (Benoit et al., Reference Benoit, Ruf, Miyamae, Fernandez, Rodrigues and Rubidge2019). One of the foramina is anteriorly oriented and located at the midline of the canine root, close to the nasal openings. The other two are laterally oriented, one dorsal to the ventral margin of the maxilla above the canine root, and the other at the second postcanine alveolar level. The two anterior foramina could represent the anterior infraorbital foramina, and the posterior one the foramen for the rostral alveolar canal (according to the homology of the infraorbital foramina proposed by Benoit et al., Reference Benoit, Ruf, Miyamae, Fernandez, Rodrigues and Rubidge2019).
Nasal.—The nasal bones are elongated, extending throughout the dorsal surface of the snout. They greatly expand laterally shortly before contact with the lacrimals and prefrontals (Fig. 2). The suture with the frontals is serrated and perpendicular to the skull sagittal plane (not visible on the 3D model). The elevation observed on the sutures between the nasals appears to be a result of taphonomic processes, because the right bone is more elevated in comparison with the left one (Fig. 8.1). A similar structure was described in Chiniquodon omaruruensis by Mocke et al. (Reference Mocke, Gaetano and Abdala2020).
Frontal.—The frontal bones compose the skull roof and are situated medially between the orbits. They have an arrow shape, in which the greater lateral expansions are anteriorly located, close to the suture with the nasals (Fig. 2). The lateral projections of the ‘arrow’ contact the prefrontals. The medial suture that separates the left and right frontals form a small crest clearly visible on the 3D model (Fig. 7.1). The frontals in Chiniquodon theotonicus and Chiniquodon sanjuanensis are smaller and lack the lateral projections (Romer, Reference Romer1969b, Reference Romer1973; Abdala, Reference Abdala1996; Martínez and Forster, Reference Martínez and Forster1996). The internal surface of the frontals forms a groove, which delimits the dorsal region of the olfactory bulbs (Hoffmann et al., Reference Hoffmann, Rodrigues, Soares and Andrade2021).
Lacrimal.—The lacrimals are bordered anteriorly by the maxillae, medially by the nasals and posteromedially by the prefrontals. A small portion of the dorsal region contacts the jugal on a marked protuberance located on the anterior edge of the orbital openings (Fig. 8). This protuberance is also present in Chiniquodon omaruruensis, although the suture in that species is between the maxillary and lacrimal bones (Mocke et al., Reference Mocke, Gaetano and Abdala2020). The suture with the prefrontals extends throughout the orbital interior cavity, where the two lacrimal foramina are located (Fig. 4), as commonly seen in other nonmammalian synapsids (e.g., Benoit et al., Reference Benoit, Manger and Rubidge2016, Reference Benoit, Ruf, Miyamae, Fernandez, Rodrigues and Rubidge2019). Two small foramina are located on the rostral external surface of the lacrimals (almost in the suture between the maxillary and lacrimal bones), which could correspond to the zygomaticofacial foramen (Benoit et al., Reference Benoit, Ruf, Miyamae, Fernandez, Rodrigues and Rubidge2019).
Prefrontal.—The prefrontals contact the nasals anteromedially, the postorbitals posteriorly, and the lacrimals anteroventrally. The suture with the frontals is straight, short, and slightly oblique to the sagittal plane of the skull (Fig. 2). It contacts the ascending process of the palatine and the pterygoids in the orbital cavity (Figs. 4, 8). The CT-scan revealed two foramina laterally oriented on the prefrontal portion of the orbital cavity (Fig. 8.1), visible on both sides of the skull.
Postorbital.—The postorbitals protrude in relation to the frontals and present a V-shape. The posterior processes are confluent in direction with the suture with the parietals, forming the anterior border of the temporal fenestra (Fig. 2). Although the sutures with the jugal are not clear on the surface of the skull (Fig. 4), the CT-scan images revealed an overlapping suture between those two bones. The dorsal surface of the postorbital bar is composed of the postorbitals, whereas the ventral region is 50% formed by the jugal (Fig. 8).
Jugal.—The jugal composes the ventral and the posterior edges of the orbital openings (~50% of the circumference). The lateral ascending process connects to the postorbitals, forming the postorbital bar. Laterally, the jugal forms nearly half of the zygomatic arch, but not reaching the posterior region (Figs. 4, 8). The suture between the jugal and squamosal begins horizontally on the anterior dorsal region of the zygomatic arch and extends ventrally on the middle, almost at a right angle. The zygomatic arch is deep and arched, with its posterodorsal edge forming more than half of the orbit height (Fig. 4), in contrast to the specimens from the Chañares Formation (Romer, Reference Romer1969b, Reference Romer1973).
Squamosal.—The squamosal forms nearly half of the zygomatic arch, the posterior squamosal sulcus (external auditory meatus) is incipient, and a deep V-shape notch separates the lambdoidal crest from the zygoma (Fig. 5). On the ventral region of the squamosal, a notch to the quadrate is present, although this bone is not preserved. The lambdoidal crest is thin and strongly projected, extending posteriorly to the occipital condyles.
Pterygoid.—The pterygoid bones project anterolaterally into the secondary bony palate, contacting the palatines (Fig. 7.2). The pterygoid flanges are long and end in a thin process. They strongly project ventrally and are clearly visible on the lateral view of the skull (Figs. 4, 8.2). The posterior projections contact each other in the basicranial girder, beyond the level of the pterygoid processes (Fig. 7.2), and do not contact the vomer. An interpterygoid vacuity is absent. The quadrate process of pterygoid is reduced or absent (Fig. 8).
Vomer.—The externally visible portion of the vomer is a triangular bone, situated between the palatine posterior processes (Figs. 3, 7.2). Inside the nasal cavity (only visible on the CT-scan images), it delimits the dorsal region of the choana and extends anteriorly on a parallel-sided keel.
Orbitosphenoid.—This bone is distinguishable on the CT-scan images, because it is laterally covered by the prefrontals. It is somewhat broken and slightly displaced (Fig. 9.1). The orbitosphenoid is a delicate laminar bone that forms the ventral support of the olfactory bulbs (Benoit et al., Reference Benoit, Jasinoski, Fernandez and Abdala2017). Within the nonmammaliaform cynodonts, it was described for Procynosuchus delaharpeae Broom, Reference Broom1938, Galesaurus planiceps Owen, Reference Owen1860, Thrinaxodon liorhinus Seeley, Reference Seeley1894, Cynosaurus suppostus (Owen, Reference Owen1876), Lumkuia fuzzi Hopson and Kitching, Reference Hopson and Kitching2001, Chiniquodon theotonicus, Probainognathus jenseni Romer, Reference Romer1970, Prozostrodon brasiliensis, Brasilodon quadrangularis Bonaparte et al., Reference Bonaparte, Martinelli, Schultz and Rubert2003, Trirachodon sp., Massetognathus pascuali Romer, Reference Romer1967, Exaeretodon riograndensis Abdala, Barberena, and Dornelles, Reference Abdala, Barberena and Dornelles2002, Kayentatherium wellesi Kermack, Reference Kermack1982, Tritylodon longaevus Owen, Reference Owen1884, and Diademodon tetragonus Seeley, Reference Seeley1894 (see Hopson, Reference Hopson, Gans, Northcutt and Ulinski1979; Quiroga, Reference Quiroga1979; Sues, Reference Sues1986; Bonaparte et al., Reference Bonaparte, Martinelli and Schultz2005; Kemp, Reference Kemp2009; Benoit et al., Reference Benoit, Jasinoski, Fernandez and Abdala2017; Crompton et al., Reference Crompton, Musinsky, Rougier, Bhullar and Miyamae2017; Pavanatto et al., Reference Pavanatto, Kerber and Dias-Da-Silva2019; Stefanello et al., Reference Stefanello, Martinelli, Müller, Dias-da-Silva and Kerber2023).
Braincase.—The poor preservation that affected the pterygoids posteriorly also affected the basicranial girder, but the posterior part of the basisphenoid remains preserved. The fenestra rotunda and the jugular foramen are separated and located in a common fossa (jugular fossa) (Figs. 3, 9), differing from the condition in other early-diverging probainognathians (e.g., Probainognathus spp.) in which both foramina are completely or partially confluent (e.g., Rougier et al., Reference Rougier, Wible and Hopson1992; Luo, Reference Luo, Fraser and Sues1994; Stefanello et al., Reference Stefanello, Martinelli, Müller, Dias-da-Silva and Kerber2023). The fenestra ovalis is relatively small, facing laterally, without a rimmed margin (Fig. 9). Two carotid foramina are present on the basisphenoid (Figs. 3, 9). The sagittal crest is slightly higher than the rest of the skull (Fig. 4), in contrast to the posteriorly tall, sigmoid crest highly elevated posteriorly described for Chiniquodon omaruruensis and some specimens of Chiniquodon theotonicus (see Mocke et al., Reference Mocke, Gaetano and Abdala2020). There is no evidence of a parietal foramen. A wide interorbital vacuity is present. It is posteriorly delimited by the ascending process of the epipterygoid, dorsally by the frontals, and anteroventrally by the palatine (Fig. 9.1).
The lateral walls of the braincase are damaged (especially on the left side of the skull) and filled by matrix. Consequently, accurate visualization of the sutures between the prootic, opisthotic, and epipterygoid on the external surface of the skull is not possible. The CT-scan images show a trigeminal foramen on the right side of the skull, between the prootic and epipterygoid. However, we could not distinguish the suture between the prootic and opisthotic (cited here as prootic-opisthotic complex).
Secondary bony palate.—The secondary bony palate is well developed (7.99 cm; Table 1), extending back to the posterior region of the postcanine row (Figs. 3, 7.2). It comprises ~48% of the basal skull length, with ~37.5% formed by the maxillae (Fig. 8.2). The suture between the palatines and maxillae is serrated (Fig 3) and is situated between the sixth postcanines (Figs. 3, 7.2). The palatine contacts the maxilla anterolaterally, delimits the choanal laterally and ventrally, and contacts the pterygoid posterolaterally (Fig. 3). A small ectopterygoid is visible on the left side of the skull (Fig. 7.2), delimited by the palatine projections of the pterygoid and not contacting the maxillae. This bone has also been described in some chiniquodontid specimens from Argentina (Abdala, Reference Abdala1996).
Occipital plate.—The lambdoidal crest forms a deep V-shaped notch between the squamosal and parietal, as in most of the cynognathian cynodonts, e.g., Luangwa drysdalli Brink, Reference Brink1963, Exaeretodon riograndensis, Massetognathus ochagaviae Barberena, Reference Barberena1981), and some nonmammaliaform probainognathians (e.g., Probainognathus jenseni, Chiniquodon theotonicus; Brink, Reference Brink1963, Romer, Reference Romer1970, Abdala, Reference Abdala1996, Abdala et al., Reference Abdala, Barberena and Dornelles2002, Liu et al., Reference Liu, Soares and Reichel2008). The post-temporal foramen is completely enclosed by the tabular (Fig. 5). The condyles are double, oval, and separated by a notch on the basioccipital. The paroccipital process is located at the base of the occipital complex and is dorsally delimitated by the tabular, and medially by the exoccipital. The sutures of the postpariental are not clear, even on the CT-scan images, but their presence can be inferred. The foramen magnum is delimited dorsally by the supraoccipital, laterally by the exoccipital, and ventrally by the basioccipital (Fig. 5).
Dentition.—The dental formula is I4/C1/PC10. The incisive teeth are represented by four alveoli on the left premaxilla (Fig. 3). A small diastema separates them from the canine. The canines are well developed, with a prominent root and without denticulations. A diastema between the canine and postcanines is absent. The left canine is slightly smaller in comparison to the right canine, and a small space is present on the left canine alveolus (Fig. 3.2). This might indicate a replacement tooth. The postcanines are laterally compressed, with a single row of cusps mesiodistally aligned and without a cingulum. The main cusp is strongly recurved; there is a distal cusp but not a mesial one. Only the left postcanines 8–10, right postcanines 2–4, and both canines are preserved. The postcanine rows are slightly curved and extend laterally to the subtemporal fenestra (Fig. 3).
Remarks
The holotype (MCP 1600 PV) was first described as ‘Probelesodon’ kitchingi (Sá-Teixeira, Reference Sá-Teixeira1982).
Results
Specimen preservation
Overall, the skull MCP 1600 PV is quite well preserved (Figs. 2–5). Most sutures are easily distinguishable; bone texture lacks signs of alteration or abrasion; and edges lack chipped areas, spheroid deformation, and other signs of mechanical or chemical alteration. A certain degree of lateral deformation is evident, but quite limited, because the skull retains a most bilaterally symmetric profile. The three-dimensional preservation of orbits, nasal cavity, and choanal region provide evidence that the specimen suffered no more than the slightest dorsoventral compression. A few teeth are broken, but the preservation of delicate pterygoid laminae and other bony edges reveal damage due to transport probably preceding burial, and that burial itself probably occurred prior to the necrolysis of soft tissues that enclosed the skull. There are no signs of bite marks, or any signs that can be related to agonistic behavior or necrophagy. Despite all of that, the teeth do show mild signs of chemical degradation, which is probably postburial.
Despite the seemingly well-preserved external surface, the CT-scan images revealed the results of the taphonomic process in the internal cavities, in which the most fragile bones were especially affected. Inside the nasal and maxillary cavities, the bones that compose the secondary bony palate are fragmented and broken. Consequently, the suture between the maxillae and palatines could not be identified on the CT-scan images, and, therefore, is not visible on the 3D model (Fig. 7.2). The delicate vomer is fragmented throughout the nasal cavity, but its maxillary portion is better preserved and distinguishable. Internal endocast features, however, remained visible, allowing the evaluation of internal neurological features and calculation of the Encephalization Quotient (EQ) in this specimen (see Hoffmann et al., Reference Hoffmann, Rodrigues, Soares and Andrade2021).
General skull morphology
The basal skull length is 16.4 cm, larger than most specimens of Chiniquodon from Argentina (Chiniquodon theotonicus and Chiniquodon sanjuanensis) and Brazil (Chiniquodon theotonicus and Chiniquodon sp.) but smaller than two specimens formerly referred to ‘Belesodon’ Huene, Reference Huene1936 and some referred to A. cromptoni, from Brazil (Table 2). The snout composes ~38% of the skull length, the orbits 17%, and the temporal region 36%. These proportions are similar to the South American Chiniquodon specimens (Abdala and Giannini, Reference Abdala and Giannini2002) and Chiniquodon omaruruensis from Namibia (Mocke et al., Reference Mocke, Gaetano and Abdala2020). The orbital openings are located on the median region of the skull and are anterolaterally oriented. They present an overall circular shape, with small projections toward the lacrimals, visible in lateral view (Fig. 4). The temporal fenestrae are somewhat rectangular, and widest at midlength (Fig. 2).
Table 2. Basal skull length (BSL, in cm) of chiniquodontid species with complete or nearly complete skulls. Holotypes in bold font. * = holotype of the former species ‘Belesodon magnificus,’ synonymized with Chiniquodon theotonicus and considered an advanced ontogenetic stage of the species due to its size (Abdala and Giannini, Reference Abdala and Giannini2002).

The CT-scan images revealed the following morphological structures: (1) sutures not visible on the external surface of the skull (e.g., between the jugal and postorbital bones); and (2) the trigeminal foramen between epipterygoid and prootic. In addition, it allowed the clear visualization and confirmation of cranial foramina.
Comparative morphology with chiniquodontid species
Specimen MCP 1600 PV presents an overall cranial structure and proportions similar to other chiniquodontid species, including a well-developed secondary bony palate, a prominence formed by the root of the canines on the maxilla, sectorial postcanines with the main cusp strongly recurved, a suborbital angulation between the jugal and maxillary bones, and pterygoid processes long and ending in a thin projection (Romer, Reference Romer1969b, Reference Romer1973; Abdala, Reference Abdala1996; Abdala and Giannini, Reference Abdala and Giannini2002).
However, the external morphological analysis and the CT-scan images revealed some differences of MCP 1600 PV in relation to the other chiniquodontids: (1) absence of pointed snout tip, composed by the anterior portions of premaxillae in dorsal and lateral views; (2) extension of the posterior projections of nasals to the anterior portions of frontals, visible in dorsal view; (3) medial contact between the posterior rami of postorbitals; (4) presence of an ectopterygoid and absence of contact with the maxilla; (5) greater participation of the maxillae in the secondary bony palate, with the maxilla longer than the palatine; (6) angled suture between the jugal and the squamosal on the zygomatic arch (~90°); (7) zygomatic arch more arched and developed dorsoventrally, with the posterodorsal edge forming above half of the orbit height, also present in Chiniquodon sanjuanensis (see Martínez and Forster, Reference Martínez and Forster1996); (8) the presence of a step on the ventral margin of the maxillae between the last incisor and canine, also present in Chiniquodon omaruruensis (see Mocke et al., Reference Mocke, Gaetano and Abdala2020); (9) a straight sagittal crest in lateral view, also seen in Chiniquodon sanjuanensis; and (10) the presence of 10 postcanine teeth.
The holotype of ‘Probelesodon’ kitchingi differs from Aleodon spp. mainly in dental morphology and the slenderly built skull. Aleodon spp. have nonsectorial postcanine teeth on the posterior region of the tooth row, bearing a conspicuous, basined lingual platform (Crompton, Reference Crompton1955; Martinelli et al., Reference Martinelli, Kammerer, Melo, Neto, Ribeiro, Da-Rosa, Schultz and Soares2017). Also, the sectorial portions of the postcanines of Aleodon spp. have less-developed carnassial cusps, but they are still recurved (Martinelli et al., Reference Martinelli, Kammerer, Melo, Neto, Ribeiro, Da-Rosa, Schultz and Soares2017). The chiniquodontid Chiniquodon kalanoro from Madagascar was not used for comparison due to the lack of comparative cranial material, because the species was described only from a lower jaw (Kammerer et al., Reference Kammerer, Flynn, Ranivoharimanana and Wyss2010).
In addition, MCP 1600 PV exclusively differs from the original description of Chiniquodon theotonicus (see Abdala, Reference Abdala1996 and Romer, Reference Romer1969b) in the following features: (1) a proportionally longer and slender snout in relation to skull length and width; (2) ‘arrow’-shaped frontals versus delta-shaped in Chiniquodon theotonicus; (3) serrated suture between frontals and nasals versus ‘smooth’ and transverse contact in Chiniquodon theotonicus; and (4) lower sagittal crest, whereas in Chiniquodon theotonicus, the skull roof profile is sigmoid in lateral view.
Discussion
Cranial morphology differences of MCP 1600 PV in relation to the other chiniquodontids
The morphological differences observed between MCP 1600 PV and Chiniquodon theotonicus, Chiniquodon sanjuanensis, and Chiniquodon omaruruensis are discussed below. Notwithstanding, we note that some of the differences could be related to ontogenetic or intraspecific variation, and thus should not be used as diagnostic characteristics.
Absence of a pointed snout tip.—In Chiniquodon theotonicus, the anterior portions of the premaxillae form a prominence, visible in lateral and ventral views (Romer, Reference Romer1969a, figs. 7, 8; Reference Romer1969b, figs. 2, 5); the anterior border of the snout is rounded in MCP 1600 PV. However, it can be argued that the absence of this feature is a taphonomic bias, but there is no evidence of broken bone on the internarial bar and premaxillae of the specimen.
Posterior projections of nasals extending to the anterior portions of frontals.—The suture between these two bones is visible in dorsal view, but the projections are absent in Chiniquodon theotonicus (see Romer, Reference Romer1969a, Reference Romerb), with a serrated suture perpendicular to the sagittal plane of the skull. However, further comparisons are needed to confirm if the nasal projections are an exclusive feature of MCP 1600 PV, an ontogenetic variation, or an observational bias due to the poor preservation of most of the chiniquodontid specimens.
Medial contact between the posterior rami of postorbitals.—In Chiniquodon theotonicus, the posterior projections are well developed, but do not contact each other on the external surface of the skull. In contrast, the postorbitals are widely separated by the anterior portions of the parietals in Chiniquodon sanjuanensis (see Martínez and Forster, Reference Martínez and Forster1996). However, the CT-scan images of MCP 1600 PV confirmed that the posterior projections of the postorbitals overlap the parietal bone, contacting each other medially. Consequently, CT-scan data of other chiniquodontids are needed to confirm if this is an exclusive feature of MCP 1600 PV.
Absence of contact between the ectopterygoid and maxilla.—In Chiniquodon theotonicus, the ectopterygoid is absent (Romer, Reference Romer1969a), but this bone is present and contacts the maxillae in some Chiniquodon theotonicus specimens previously described by Romer (Reference Romer1969b) as ‘Probelesodon lewisi.’ However, the absence of an ectopterygoid is considered a synapomorphy for Probainognathia (Hopson and Kitching, Reference Hopson and Kitching2001), and the presence of this bone in MCP 1600 PV should be treated with caution until new specimens are discovered. Also, the presence of this bone in MCP 1600 PV and ‘Probelesodon lewisi’ can be useful to further explore the taxonomic status of the latter taxon, synonymized with Chiniquodon theotonicus.
Greater participation of the maxillae on the secondary bony palate.—In specimens with a well-preserved secondary bony palate, the palatines form approximately two-thirds of this structure (Romer, Reference Romer1969b; Sá-Teixeira, Reference Sá-Teixeira1982; Abdala, Reference Abdala1996). However, in MCP 1600 PV, the maxillae have a greater contribution on the secondary bony palate (~37.5%), and it seems to be associated with the elongation of the secondary bony palate in the specimen compared to the other chiniquodontids.
Greater participation of the jugal on the zygomatic arch.—In MCP 1600 PV, the suture between the jugal and squamosal forms nearly a 90° angle, increasing the participation of the jugal on the zygomatic arch. In contrast, this suture forms a more subtle curve in the other chiniquodontids.
Zygomatic arch more developed dorsoventrally.—In Chiniquodon theotonicus, Chiniquodon sanjuanensis, and Chiniquodon omaruruensis, the zygomatic arch is more gracile and slenderer dorsoventrally, whereas in MCP 1600 PV, it gradually expands in depth posteriorly. In addition, this feature is not related to the degree of curvature of the zygomatic arch, in which the dorsal region reaches the level of the middle of the orbit or higher (present in MCP 1600 PV, Chiniquodon sanjuanensis, and Chiniquodon omaruruensis).
Height of the sagittal crest.—This feature is highly variable within chiniquodontid specimens, therefore, it should not be considered diagnostic. In addition, the development of the sagittal crest can be a result of ontogenetic variation (discussed below in ‘ontogenetic considerations’) or sexual dimorphism (e.g., Flores et al., Reference Flores, Abdala and Giannini2010; Jasinoski et al., Reference Jasinoski, Abdala and Fernandez2015; Martinelli et al., Reference Martinelli, Kammerer, Melo, Neto, Ribeiro, Da-Rosa, Schultz and Soares2017).
Postcanine number.—This feature is highly variable within chiniquodontid specimens (see Abdala and Giannini, Reference Abdala and Giannini2002, table 1) and is seemingly not related to basal skull length (BSL). Specimens with approximately the same BSL present some variation in postcanine number (e.g., PULR 12 and MCZ 4100, each first described as ‘Probelesodon minor’). In contrast, specimens with different BSLs have 10 postcanines (e.g., MCP 1600 PV, UFRGS-PV-0274-T, and MCZ 3777). Hence, this feature should not be used as a diagnostic trait.
The length and width of the snout in relation to the same skull proportions.—These features were briefly discussed by Abdala (Reference Abdala1996). The author cited the similarity in the general skull proportions among the MCP 1600 PV and the specimens from the Chañares Formation: slender snout and cranial structure more gracile in comparison to Chiniquodon theotonicus specimens from Santa Maria Supersequence (Fig. 10). This feature can also be useful to further explore the taxonomic status of the specimens from the Chañares Formation, which might represent a different species or even be included in Chiniquodon kitchingi. However, an extensive morphological analysis is needed to evaluate this feature in all chiniquodontid specimens.
The shape and size of frontals.—In MCP 1600 PV, the frontals are ‘arrow’-shaped and in Chiniquodon theotonicus they are delta-shaped, with lateral expansions toward the prefrontals (Romer, Reference Romer1969a, fig. 6). An extensive comparison of the shape and size of frontals within the chiniquodontids sample is not possible due to the poor preservation of some materials (e.g., specimens from the Santa Maria Supersequence), which present a ‘crackle-ware’ bone surface (Romer, Reference Romer1969a).
Frontal and nasal bones suture.—In MCP 1600 PV and some Chiniquodon theotonicus specimens from the Chañares Formation (PULR 18 and MCZ 3781, 4002; see Romer, Reference Romer1969b), the suture is serrated, and in Chiniquodon theotonicus it is, in general, smooth and straight. Considering this, there are three possible conclusions: (1) the serrated suture is a characteristic of the genus ‘Probelesodon,’ because the specimens cited by Romer (Reference Romer1969b) were previously described as ‘Probelesodon lewisi’; (2) it could be an ontogenetic trait because similar changes have been reported in an ontogenetic series of Thrinaxodon Seeley, Reference Seeley1894 (Jasinoski et al., Reference Jasinoski, Abdala and Fernandez2015); and (3) the straight suture on the schematic reconstruction of Chiniquodon theotonicus (see Romer, Reference Romer1969a, fig. 6) could be a result of the poor preservation of the skull stated by the author and thus, a misidentification of structure.
Separation of the fenestra rotunda and the jugular foramen.—In Chiniquodon theotonicus, both foramina are completely confluent (Romer, Reference Romer1969a; Kemp, Reference Kemp2009), whereas in the specimen here described, the jugular foramen seems completely separated from the fenestra rotunda (Figs. 3, 9). In nonprobainognathian cynodonts, early diverging probainognathians (e.g., Lumkuia Hopson and Kitching, Reference Hopson and Kitching2001, Probainognathus Romer, Reference Romer1970), and nonmammaliaform prozostrodontians (e.g., Pseudotherium Wallace, Martínez, and Rowe, Reference Wallace, Martínez and Rowe2019, Brasilodon Bonaparte et al., Reference Bonaparte, Martinelli, Schultz and Rubert2003), the jugular foramen and fenestra rotunda are totally or fully confluent (e.g., Rougier et al., Reference Rougier, Wible and Hopson1992; Luo, Reference Luo, Fraser and Sues1994; Hopson and Kitching, Reference Hopson and Kitching2001; Bonaparte et al., Reference Bonaparte, Martinelli and Schultz2005; Wallace et al., Reference Wallace, Martínez and Rowe2019). The condition in MCP 1600 PV could be an autapomorphy among chiniquodontids.
Ontogenetic considerations
When is not possible to perform allometric analysis due to the lack of or a low number of specimens available, qualitative ontogenetic variation should be considered in comparative morphological analysis. Basal skull length and discrete cranial features, e.g., the shape of bone sutures, or the absence/presence of structures, can be used to determine the ontogenetic stages of fossil specimens (Jasinoski et al., Reference Jasinoski, Abdala and Fernandez2015).
Considering this, four of the 13 observed and discussed skull differences between MCP 1600 PV and other chiniquodontids can be associated with a variation in growth rates of the skull during ontogeny, considering previous studies on nonmammaliaform cynodonts and mammals (e.g., Estes, Reference Estes1961; Flores et al., Reference Flores, Abdala and Giannini2010; Jasinoski et al., Reference Jasinoski, Abdala and Fernandez2015; Jasinoski and Abdala, Reference Jasinoski and Abdala2016): (1) The nasal-frontals suture in MCP 1600 PV (serrated, with the posterior projections of nasals advancing toward the anterior portions of frontals) is commonly associated with juvenile specimens (Estes, Reference Estes1961; Jasinoski et al., Reference Jasinoski, Abdala and Fernandez2015), although, in the epicynodont Galesaurus planiceps, this feature does not change throughout ontogenetic stages (Jasinoski and Abdala, Reference Jasinoski and Abdala2016). (2) The posterior rami of postorbitals are usually longer in adults or subadults (Jasinoski et al., Reference Jasinoski, Abdala and Fernandez2015; Jasinoski and Abdala, Reference Jasinoski and Abdala2016), therefore, MCP 1600 PV presents the condition associated with adult specimens (long postorbital posterior projections). This condition also corroborates the interpretation of the Chiniquodon sanjuanensis holotype as a juvenile, with 8.0 cm of BSL and short posterior rami of postorbitals (Abdala and Giannini, Reference Abdala and Giannini2002). However, the medial contact of these projections is not yet described as an ontogenetic characteristic. (3) The degree of development of the sagittal crest is related to ontogenetic variation, with younger specimens usually presenting a lower sagittal crest, resulting in the same height between the snout and the temporal regions (Jasinoski et al., Reference Jasinoski, Abdala and Fernandez2015; Jasinoski and Abdala, Reference Jasinoski and Abdala2016). The chiniquodontid specimens from Brazil and Argentina are apparently divided into two groups, independent of size: the sample from the Chañares Formation (formerly described in ‘Probelesodon’ Romer, Reference Romer1969b) and MCP 1600 PV with a lower sagittal crest, and the remaining Chiniquodon theotonicus specimens, with a marked concave outline between the sagittal crest and the roof of the snout in lateral view (see Martinelli et al., Reference Martinelli, Kammerer, Melo, Neto, Ribeiro, Da-Rosa, Schultz and Soares2017, fig. 17). Consequently, the height of the sagittal crest should be reviewed in further comparative studies. (4) The number of postcanines can be used as an indication of ontogenetic stage (Abdala et al., Reference Abdala, Jasinoski and Fernandez2013; Norton et al., Reference Norton, Abdala, Rubidge and Botha2020). This feature has not yet been studied in chiniquodontid species, but the postcanine number is highly variable within the smaller and larger Argentinian and Brazilian specimens (Martínez and Forster, Reference Martínez and Forster1996; Abdala and Giannini, Reference Abdala and Giannini2002).
The results provided here do not disprove the allometric tests and the arrangement of chiniquodontid skulls in a continuous growth series performed by Abdala and Giannini (Reference Abdala and Giannini2002). However, the authors also discussed the additional support provided by the lack of qualitative traits useful for diagnosis in chiniquodontid specimens. Eight of the 13 cranial features discussed here are not ontogenetic variations. The specimen MCP 1600 PV, in addition, presents a combination of features related to juvenile and adults; thus, they can be used as evidence that ‘Probelesodon’ kitchingi might not be a subjective junior synonym of Chiniquodon theotonicus. As such, we consider the species valid but placed it in Chiniquodon Huene, Reference Huene1936, therefore, Chiniquodon kitchingi cannot be considered within the Chiniquodon theotonicus skull growth model. However, further specimens and morphometric and allometric analyses are needed to elucidate the taxonomy of Chiniquodon and its closest allies.
Conclusion
The differences observed in the cranial morphology of MCP 1600 PV compared to Chiniquodon theotonicus, Chiniquodon omaruruensis, and Chiniquodon sanjuanensis raised doubt on the synonymization of these species. In addition, the results highlight the importance of a reassessment of the family to elucidate if the heterogeneity in skull features within Chiniquodontidae is a result of intraspecific variation, sexual dimorphism, or ontogeny. At this point, considering the differences not related to ontogeny observed in MCP 1600 PV from features in other chiniquodontids, we maintain the species ‘Probelesodon’ kitchingi as valid within Chiniquodon.
Moreover, the CT-scan and analysis of the 3D model are relevant for the study and a better understanding of the cranial morphology of Chiniquodon kitchingi. It also allowed the accurate visualization and confirmation of fossil characteristics that were covered by matrix. Therefore, the CT-scan is a relevant tool for further analyses of new chiniquodontid specimens, as well as in the study of already described and published materials.
Acknowledgments
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Brasil (CAPES)-Finance Code 001 (process number 88887.179660/2018-00), and Paleontological Society (PalSIRP Sepkoski Grants 2016). We are indebted to J. Borges and D. Anschau, Instituto do Cérebro (InsCer/PUCRS), for providing access to the medical CT-scanner and for showing interest in paleontological research. Special thanks to P. Henrique Morais (UFRGS) for helping with software use and discussions on the morphology of cynodonts. Finally, we thank C. Kammerer and F. Abdala for their thorough and constructive reviews of this manuscript, and H.-D. Sues for editing.
Declaration of competing interests
The authors declare none.
Data availability statement
Data available from the MorphoMuseuM: http://morphomuseum.com/Specimenfiles/sendFile/1151/b9634c. See Hoffmann et al. (Reference Hoffmann, Martinelli and Brandalisem2023).












