


The gut epithelial integrity and barrier function has been a central predisposing factor in the pathogenesis of inflammatory bowel disease (IBD) and its related gastrointestinal diseases, food allergies, type 1 diabetes and other autoimmune diseases( Reference Zeissig, Burgel and Gunzel 1 – Reference Vaarala 6 ). Gut epithelium is constantly renewing, which extensively involves epithelial cell proliferation and differentiation. Disruption of this process impairs epithelial barrier function, which underlies many digestive-related diseases. In IBD, such as Crohn's disease and ulcerative colitis, the gastrointestinal tract has been found to be damaged due to extensive mucosal hyperplasia, inflammatory reactions and abnormal expression of MHC class II molecules in the intestinal epithelial cells( Reference Maloy and Powrie 4 ).
Intestinal cell proliferation and differentiation are regulated by various signalling pathways and mediators( Reference van der Flier and Clevers 7 ). Bone morphogenetic protein 2 (Bmp2) functions to promote the overall epithelial differentiation; hairy and enhancer of split 1 (Hes1) regulates the differentiation of enterocytes; atonal homolog 1 (Atoh1) is essential for the formation of secretory cells; and Krueppel-like factor 4 (Klf4) is one of the factors important for the development of goblet cells( Reference van der Flier and Clevers 7 ). Growth arrest and DNA damage (GADD45) inducible protein levels are elevated under stresses in order to arrest the cell cycle and promote cell survival( Reference Mak and Kultz 8 ), and proliferating cell nuclear antigen (PCNA) is a commonly used marker of cell proliferation( Reference Kubben, Peeters-Haesevoets and Engels 9 ). In addition to the above-mentioned pathways and mediators, the Wingless and Int (Wnt) signalling is one of the most fundamental mechanisms driving the proliferation of intestinal epithelial cells, and alters cell fate commitment and tissue homeostasis. The Wnt/β-catenin signalling regulates cell proliferation through mediating the transcription of many target genes including cyclin D1 (Ccnd1) and myelocytomatosis oncogene protein (Myc)( Reference Wang, Lisanti and Liao 10 , Reference Xue, Wang and Zhao 11 ). Dysregulation of the Wnt/β-catenin pathway has been implicated in IBD-associated carcinogenesis as well as colorectal cancer( Reference Claessen, Schipper and Oldenburg 12 , Reference Serafino, Moroni and Zonfrillo 13 ). Ccnd1 expression was up-regulated in active ulcerative colitis- and ulcerative colitis- related carcinogenesis( Reference Wong, Mayer and Anderson 14 ). The aminosalicylate mesalazine drug exerts chemopreventive effects to IBD through inhibition of the Wnt/β-catenin signalling( Reference Bos, Diks and Hardwick 15 ). Polyphenols down-regulate the Wnt/β-catenin signalling in colonic cancer cells( Reference Wang, Wise and Li 16 , Reference Taira, Uehara and Tsuchida 17 ); however, relatively less is known about their effects on the proliferation of gut epithelial cells in vivo.
Grape seed extract (GSE) is a heterogeneous mixture of polyphenols (anthocyanidins, catechins and their derivatives) extracted from grape seed and skin. A body of evidence has shown that GSE has anti-inflammatory, antioxidant, cardioprotective, hepatoprotective, neuroprotective, anti-diabetic, anti-carcinogenic and anti-ageing effects( Reference Terra, Montagut and Bustos 18 – Reference Yang, Wang and Kang 26 ). Our previous study using 1 % (w/w) GSE supplementation attenuated intestinal inflammation and ameliorated the colitis symptom in IL10-deficient mice( Reference Wang, Xue and Zhang 27 ). In a different study, 0·5 % (w/w) GSE supplementation has been shown to reduce the cell proliferation rate in the small intestine of the APC knockout mice( Reference Velmurugan, Singh and Kaul 28 ). In addition, 0·1 % GSE supplementation has been shown to increase the intestinal occludin content and reduce the faecal neutrophil protein calprotectin level in mice fed with a high-fat diet( Reference Goodrich, Fundaro and Griffin 29 ). IL10-deficient mice develop spontaneous enterocolitis similar to human Crohn's disease( Reference Kuhn, Lohler and Rennick 30 ), and have been widely used to study the mucosal structure damage and barrier function impairment( Reference Zhou, Streutker and Borojevic 31 , Reference Haub, Ritze and Bergheim 32 ). Using this mouse model, the objective of the present study was to test whether a lower dose (0·1 % GSE) compared with that of our previous study (1 % GSE) has beneficial effects, and to further explore the mechanistic changes linking GSE to improved gut epithelial function.
Materials and methods
Grape seed extract
The GSE product (Gravinol-S™) was purchased from OptiPure Chemco Industries, Inc. As per the company product specification sheet, the GSE product contains a minimum of 95 % flavonols, of which 82 % are oligomeric proanthocyanidins and 12 % being the highly active monomeric proanthocyanidins.
Animal care and experimental design
All animal procedures were approved by the Washington State University Animal Care and Use Committee. The homozygous IL10-deficient mice (B6.129P2-Il10 tm1Cgn/J) were purchased from The Jackson Laboratory and were bred in the Experimental Animal Laboratory Unit at the Washington State University. Mice were housed in a temperature-controlled room with a 12 h light–12 h dark cycle, and had free access to food (2018 Teklad Global 18 % Protein Rodent Diet) and drinking-water. Female mice aged 6 weeks were randomised into two groups (n 10 per group) and were given drinking-water containing 0 or 0·1 % GSE (w/v) for 12 weeks( Reference Goodrich, Fundaro and Griffin 29 ). Wild-type mice were not included in this study, because our previous study has shown that GSE supplementation has little effect on the wild-type mice( Reference Wang, Xue and Zhang 27 ). To avoid potential confounding effects of sex, only female mice were used in the present study. Water was changed on a daily basis to avoid the possible oxidation of functional compounds in GSE. During the feeding trial, mice were housed individually for the measurement of feed intake and water consumption. There was no difference in the amount of water drunk per mouse between treatment groups (3·5–4 ml/mouse per d). This equates to 140–160 mg/kg per d for an adult mouse (approximately 25 g), which converts to 680–780 mg of polyphenols for a 60 kg human as per the published formula( Reference Reagan-Shaw, Nihal and Ahmad 33 ). This indicates that the GSE concentration used in the present study was also the equivalent of the consumption of 180–205 ml of red wine (3·8 g/l total polyphenols)( Reference Rifler, Lorcerie and Durand 34 ) per d for a 60 kg adult.
In vivo intestinal permeability
At the 10th week of the feeding trial, in vivo intestinal permeability was measured as per the published method( Reference Cani, Bibiloni and Knauf 35 ). Briefly, mice that had been fasted for 6 h were administered with fluorescein isothiocyanate-dextran (500 mg/kg body weight; Sigma-Aldrich). After 1 h of administration, blood sample was collected from the orbital sinus and centrifuged at 12 000 g for 5 min at 4°C. The resulting serum was diluted 1:3 with PBS (pH 7·4), and the fluorescence intensity was measured using the Synergy H1 Hybrid Multi-Mode Microplate Reader (BioTek Instruments) at an excitation wavelength of 485 nm and an emission wavelength of 535 nm.
Faecal total antioxidant capacity assay and serum TNF-α measurement
Faecal samples from the 11th week of the feeding trial were powdered in liquid N2 and then homogenised in 80 % methanol solution. Scavenging activities of the faecal homogenate against the stable free radical 2, 2-diphenyl-1-picrylhydrazyl (DPPH; Sigma-Aldrich) were assayed according to the published methods( Reference Chrzczanowicz, Gawron and Zwolinska 36 , Reference Magalhaes, Barreiros and Maia 37 ) with modification. Briefly, 50 μl of diluted faecal samples or 80 % (v/v) methanol (Blank) were mixed with 150 μl of 60 μm-DPPH solution. The absorbance at 517 nm was monitored during a 2 h incubation period using the Synergy H1 Hybrid Microplate Reader (BioTek Instruments). All experiments were performed in triplicate, and the percentage of decrease in DPPH reading was calculated using the following equation:
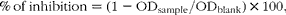 $$\begin{eqnarray} \%\,of\,inhibition = (1 - OD_{sample}/OD_{blank})\times 100, \end{eqnarray}$$
$$\begin{eqnarray} \%\,of\,inhibition = (1 - OD_{sample}/OD_{blank})\times 100, \end{eqnarray}$$
where ODsample and ODblank indicate the absorbance of faecal sample and methanol blank at a wavelength of 517 nm, respectively.
The serum collected at necropsy (at the end of the 12th week of the feeding trial) was used for analysing the level of TNF-α using the mouse TNF-α instant ELISA kit (eBiosciences) according to the manufacturer's protocol.
Tissue collection
On the day of necropsy, mice were anaesthetised intraperitoneally with tribromoethanol (250 mg/kg body weight), followed by cervical dislocation. The colon section was dissected. A 5 mm segment of colon was fixed in freshly prepared 4 % (w/v) paraformaldehyde (pH 7·0), processed and embedded in paraffin. The remaining colon tissue was cut opened longitudinally, rinsed in PBS, frozen in liquid N2 and stored at − 80°C for further biochemical analyses.
Histological examination
For pathobiological examination, paraffin-embedded colonic gut tissues were sectioned at 5 μm thickness and subjected to haematoxylin–eosin staining. Each colonic section was scored in a blinded fashion following a method published previously( Reference Burich, Hershberg and Waggie 38 ), with some modifications. Briefly, the severity of epithelial hyperplasia (scored from 0 to 3), the intensity and severity of inflammation (scored as 0–4), and the extent of mucosal hyperplasia (scored as 0–4) and extent of inflammation (scored as 0–4) were scored individually. The summation of these scores provided a total colonic inflammatory score ranging from 0 to a maximum of 15 per colonic section, with 0 being a normal tissue and 15 being the most extensive/severe disease symptoms.
For goblet cell staining, paraffin-embedded colonic tissues were sectioned at 5 μm thickness, deparaffinised and hydrated, and then stained with Alcian blue (pH 2·5)( Reference Pellegrinet, Rodilla and Liu 39 ), which stains goblet cells blue. Colonic tissue sections (five fields per section, 3–4 sections per animal, n 8) were randomly selected for the quantification of goblet cell:total area ratio using the Image J 1.30v software (split colour channels; National Institute of Health).
Immunohistochemical staining
Immunohistochemical staining was conducted as described previously( Reference Kubben, Peeters-Haesevoets and Engels 9 , Reference Zhu, Du and Hess 40 ). Briefly, colonic tissue sections were deparaffinised and hydrated, followed by antigen retrieval before incubation with anti-PCNA (Santa Cruz Biotech, Inc.), anti-β-catenin (Cell Signaling Technology) or anti-phosphorylated (phos)-β-cateninSer552 (Cell Signaling Technology) antibody. Signal was visualised using the Vectastain ABC and DAB kits (Vector Laboratories). After counterstaining with haematoxylin, images were taken using the Leica DM2000 LED light microscope (200 × ; Leica Microsystems, Inc.). The cell proliferation index was expressed as the PCNA-positive cells/total cells in the crypts. Phos-β-cateninSer552 staining was expressed as the phos-β-cateninSer552 positive crypts/100 crypts. β-Catenin staining intensity was scored semi-quantitatively from 0 (normal tissue) to 5 (intensive staining) by a trained examiner in a blinded manner, four sections per animal, eight mice per dietary group.
Quantitative RT-PCR analysis
Total RNA was extracted from powdered large intestine tissues using the TRIzol® Reagent (Sigma-Aldrich), treated with DNase I (Qiagen) and purified with RNeasy Mini kit (Qiagen). Complementary DNA was synthesised with the iScript™ complementary DNA synthesis kit (Bio-Rad). SYBR Green Master Mix (Bio-Rad) was used in all PCR. The amplification efficiency was 0·90–0·99. The quantitative RT-PCR conditions were 95°C, 3 min; 35 cycles of 95°C for 10 s, 56°C for 10 s and 72°C for 20 s. At the end of each run, dissociation melt curves were obtained to confirm the purity of PCR products. The PCR products were further confirmed by electrophoresis and sequencing. β-Tubulin (Tubb2a) was used as the housekeeping gene. The primer sequences are listed in Table 1.
Table 1 Primer sequences used in the present study

Atoh1, atonal homolog 1; Bmp2, bone morphogenetic protein 2; Myc, myelocytomatosis oncogene protein; Ccnd1, cyclin D1; Gadd45, growth arrest and DNA damage; Hes1, hairy and enhancer of split 1; Klf4, Krueppel-like factor 4; Muc, mucin; Pcna, proliferating cell nuclear antigen; Tubb2a, β-tubulin.
Immunoblotting analyses
Immunoblotting analyses were conducted as described previously( Reference Zhu, Du and Hess 40 ). The band density of target protein was normalised to the β-tubulin. Antibodies against phos-AMP-activated protein kinase (phos-AMPK), AMPK, phos-extracellular signal-regulated kinases 1 and 2 (phos-ERK1/2), ERK1/2, β-catenin were purchased from Cell Signaling Technology. Anti-β-tubulin antibody was purchased from Sigma-Aldrich. Horseradish peroxidase-conjugated anti-rabbit and anti-mouse secondary antibodies were purchased from Cell Signaling Technology.
Statistical analysis
Data were analysed as a complete randomised design using General Linear Model of Statistical Analysis System (2000). Data are presented as means with their standard errors. A significant difference was considered as P< 0·05.
Results
Effect of dietary grape seed extract on intestinal permeability, total antioxidant capacity, serum TNF-α level and histological score
There was no difference in the overall feed intake or body weight gain between the two groups after 12 weeks of feeding trial (data not shown). GSE supplementation significantly decreased the in vivo intestinal permeability (Fig. 1(a)). In addition, GSE supplementation increased the faecal total antioxidant capacity (Fig. 1(b)) and reduced the serum TNF-α level (Fig. 1(c)). However, GSE supplementation has no significant effect on the pathological score (data not shown).

Fig. 1 Intestinal permeability, faecal total antioxidant capacity and serum TNF-α level in mice fed a control (CON) or grape seed extract (GSE) supplemented drinking-water. (a) In vivo intestinal permeability measured with permeability tracer fluorescein isothiocyanate (FITC)-labelled dextran; (b) faecal total antioxidant capacity; (c) serum TNF-α level. Values are means (n 8–10), with individual values (a and b), or with standard errors represented by vertical bars (c). **Mean value was significantly different from that for CON group (P< 0·01).
Dietary grape seed extract enhanced colonic goblet cell differentiation
GSE supplementation increased the number of goblet cells (blue stained, Fig. 2(a) and (b)) associated with enhanced mRNA expression of mucin (Muc)-2, a major secretory mucin, and a tendency of increased Muc3, while had no effect on the mRNA expression of Muc1 (Fig. 2(c)). However, interestingly, no differences in the mRNA expression of Bmp2, Hes1, Atoh1 and Klf4 were observed between the control and the GSE groups (data not shown).

Fig. 2 Goblet cell density, mucin (Muc) and differentiation transcription factors mRNA expression in the colon of mice fed a control (CON; ![]() ) or grape seed extract (GSE;
) or grape seed extract (GSE; ![]() ) supplemented drinking-water. (a) Representative images of Alcian blue staining of goblet cells, original magnification at 200 × ; (b) quantification of Alcian blue staining; (c) mRNA expression of Muc1, Muc2 and Muc3. Values are means (n 8), with their standard errors represented by vertical bars. *Mean value was significantly different from that for CON group (P< 0·05). †Mean value was marginally significantly different from that for CON group (P< 0·1). A colour version of this figure can be found online at http://www.journals.cambridge.org/bjn
) supplemented drinking-water. (a) Representative images of Alcian blue staining of goblet cells, original magnification at 200 × ; (b) quantification of Alcian blue staining; (c) mRNA expression of Muc1, Muc2 and Muc3. Values are means (n 8), with their standard errors represented by vertical bars. *Mean value was significantly different from that for CON group (P< 0·05). †Mean value was marginally significantly different from that for CON group (P< 0·1). A colour version of this figure can be found online at http://www.journals.cambridge.org/bjn
Grape seed extract supplementation decreased cell proliferation
GSE supplementation reduced the number of PCNA-positive cells per crypt (Fig. 3(a) and (b)) and had a trend to decrease the mRNA expression of pcna in the control group (1·00 (sem 0·19)) compared with the GSE group (0·63 (sem 0·09)) (P <0·10). In agreement, dietary GSE significantly down-regulated mitogen-activated protein kinases/ERK growth signalling as indicated by reduced phos-ERK1/2/ERK1/2 level in the colonic tissues of GSE compared with that in control mice (Fig.3(c)). Additionally, GSE supplementation decreased the mRNA expression of Gadd45a (P< 0·05) and had a tendency to decrease Gadd45b (P< 0·1) (Fig. 3(d)).

Fig. 3 Colonic cell proliferation in the colon of mice fed a control (CON; ![]() ) or grape seed extract (GSE;
) or grape seed extract (GSE; ![]() ) supplemented drinking-water. (a) Representative images of proliferating cell nuclear antigen (PCNA) staining, original magnification at 200 × , dark brown dots indicate PCNA staining signal as pointed by red arrows; (b) ratio of PCNA-positive cells; (c) mitogen-activated protein kinases/extracellular signal-regulated kinases 1 and 2 (ERK1/2) immunoblotting (top: representative western blot images; bottom: quantification data); (d) growth arrest and DNA damage (Gadd45) mRNA expression. Values are means (n 8), with their standard errors represented by vertical bars. Mean value was significantly different from that for CON group: *P< 0·05, **P< 0·01. †Mean value was marginally significantly different from that for CON group (P< 0·1). A colour version of this figure can be found online at http://www.journals.cambridge.org/bjn
) supplemented drinking-water. (a) Representative images of proliferating cell nuclear antigen (PCNA) staining, original magnification at 200 × , dark brown dots indicate PCNA staining signal as pointed by red arrows; (b) ratio of PCNA-positive cells; (c) mitogen-activated protein kinases/extracellular signal-regulated kinases 1 and 2 (ERK1/2) immunoblotting (top: representative western blot images; bottom: quantification data); (d) growth arrest and DNA damage (Gadd45) mRNA expression. Values are means (n 8), with their standard errors represented by vertical bars. Mean value was significantly different from that for CON group: *P< 0·05, **P< 0·01. †Mean value was marginally significantly different from that for CON group (P< 0·1). A colour version of this figure can be found online at http://www.journals.cambridge.org/bjn
Dietary grape seed extract diminished Wingless and Int/β-catenin and AMP-activated protein kinase signalling
In line with the decreased epithelial cell proliferation, GSE supplementation reduced the β-catenin protein content (Fig. 4(a)), nuclear accumulation of β-catenin (Fig. 4(b) and (c)) and phos-β-cateninSer552 (Fig. 5). Consistently, β-catenin signalling target molecules, the mRNA expression of Ccnd1 and Myc, were decreased by dietary GSE supplementation (Fig. 6(a)). In agreement with decreased nuclear accumulation of phos-β-cateninSer552, the AMPK activity was reduced in the GSE-supplemented mice (Fig. 6(b)).

Fig. 4 Wingless and Int (Wnt)/β-catenin signalling in the colon of mice fed a control (CON) or grape seed extract (GSE) supplemented drinking-water. (a) Immunoblotting of β-catenin; (b) β-catenin immunohistochemical staining; (c) representative image of β-catenin staining, original magnification at 200 × . Values are means with their standard errors represented by vertical bars (n 8). *Mean value was significantly different from that for CON group (P< 0·05). A colour version of this figure can be found online at http://www.journals.cambridge.org/bjn

Fig. 5 Immunohistochemical staining of phosphorylated (phos)-β-cateninSer552 in the colon of mice fed a control (CON) or grape seed extract (GSE) supplemented drinking-water. (a) Quantification data; (b) representative images, original magnification at 200 × . Values are means with their standard errors represented by vertical bars (n 8). *Mean value was significantly different from that for CON group (P< 0·05). Yellow arrows point to the phos-β-cateninSer552-positive cells. A colour version of this figure can be found online at http://www.journals.cambridge.org/bjn

Fig. 6 AMP-activated protein kinase (AMPK) signalling, cyclin D1 (Ccnd1) and myelocytomatosis oncogene protein (Myc) mRNA expression in the colon of mice fed a control (CON; ![]() ) or grape seed extract (GSE;
) or grape seed extract (GSE; ![]() ) supplemented drinking-water. (a) mRNA expression of Ccnd1 and c-Myc (b) Immunoblotting of AMPK. Values are means with their standard errors represented by vertical bars (n 8). *Mean value was significantly different from that for CON group (P< 0·05).
) supplemented drinking-water. (a) mRNA expression of Ccnd1 and c-Myc (b) Immunoblotting of AMPK. Values are means with their standard errors represented by vertical bars (n 8). *Mean value was significantly different from that for CON group (P< 0·05).
Discussion
GSE is an extract of polyphenols, mainly catechins, proanthocyanidins and their derivatives. Accumulating evidence points to the beneficial role of GSE in intestinal inflammation and IBD, primarily mediated by proanthocyanidins( Reference Cheah, Bastian and Acott 23 , Reference Li, Cai and Qin 41 , Reference Wang, Yang and Wang 42 ). Our previous study has found that dietary 1 % GSE supplementation reduces disease indices of IBD( Reference Wang, Xue and Zhang 27 ). Intragastrical administration of GSE at a dose of 400 mg/kg reduced the qualitative histological severity score in the proximal colon of dextran sulphate sodium-induced colitis in rats( Reference Cheah, Bastian and Acott 23 ). In the present study, we tested the effectiveness of a lower dose (0·1 %) of GSE supplementation on gut inflammation, and found that GSE supplementation decreased the serum TNF-α level and increased the total antioxidant activity, which are consistent with our previous study( Reference Wang, Xue and Zhang 27 ). However, no significant differences in the pathological scores were observed between mice with/without 12 weeks of 0·1 % GSE supplementation, indicating that this low dose of GSE supplementation was insufficient to improve the gut pathological changes in the IL10-deficient mice.
A balanced rate of cell proliferation and cell differentiation is essential to maintain the intestinal epithelial homeostasis. Hyperproliferation in the intestinal tract has been observed in ulcerative colitis in both human subjects and mice( Reference Babyatsky, Rossiter and Podolsky 43 , Reference Geier, Smith and Butler 44 ). The present study found that 0·1 % of GSE supplementation reduced the colonic cell proliferation, which was consistent with the previous studies showing that GSE decreased cell proliferation in the intestinal tract of APCmin/+ mice( Reference Velmurugan, Singh and Kaul 28 ) and azoxymethane-induced Fischer 344 Rats( Reference Velmurugan, Singh and Agarwal 45 ). Inflammation is known to induce GADD45 expression( Reference Zhang, Ahsan and Zhu 46 ). Along with the decreased serum TNF-α level, GADD45 expression was down-regulated in the GSE-supplemented mice, indicating that the beneficial effects of GSE might be associated with its anti-inflammatory effect.
The goblet cell is one of the four cell lineages that are derived from the intestinal stem cells. Goblet cells secrete mucins and are one of the markers that can be used for studying cell differentiation( Reference Crosnier, Stamataki and Lewis 47 ). The depletion of goblet cells in the large intestine is a characterised feature of IBD( Reference Johansson and Hansson 48 , Reference Jadert, Phillipson and Holm 49 ). In the present study, we found that both goblet cells and Muc2 expression were enhanced in GSE-supplemented mice, indicating that GSE might result in enhanced intestinal cell differentiation, but could exert a protective effect on epithelium due to stimulation of innate immunity, given no difference was found in the expression of factors regulating lineage-specific cell differentiation including Bmp2, Hes1 Atoh1 and Klf4.
Mechanically, Wnt signalling plays a crucial role in cell proliferation and tissue homeostasis( Reference Logan and Nusse 50 ). Wnt signalling has the highest expression in the early stage of the proliferative compartment and decreases as the cells move upwards into the differentiating area( Reference Bertrand, Angus and Partis 51 ), indicating that the Wnt pathway is the dominant force behind the proliferative activity of the intestinal epithelium. In the present study, we found that GSE supplementation decreases the nuclear accumulation of β-catenin, the expression of its target genes and the ERK1/2 signalling( Reference Ding, Xia and Liu 52 ). These data are in consistent with the previous studies, showing that polyphenol compounds could attenuate aberrantly activated Wnt/β-catenin signalling( Reference Dashwood, Orner and Dashwood 53 ). Polymeric black tea polyphenols inhibit 1,2-dimethylhydrazine-induced colorectal carcinogenesis by inhibiting cell proliferation via the Wnt/β-catenin pathway( Reference Patel, Ingle and Maru 54 ). Furthermore, dietary green tea polyphenol supplementation inhibits the intestinal tumorigenesis in APC knockout mice partly through the down-regulation of β-catenin nuclear expression( Reference Hao, Sun and Yang 55 ). Recently, green tea polyphenol, (-)-epigallocatechin-3-gallate, has been shown to be toxic to human skin cancer cells by inactivation of β-catenin( Reference Singh and Katiyar 56 ). These data suggested that the inhibition of β-catenin signalling is one of the possible mechanisms underlying the beneficial effects of GSE.
AMPK is a serine/threonine protein kinase that plays an important role in maintaining cellular energy balance. In the present study, AMPK activity was suppressed in the GSE-supplemented mice associated with a decreased phosphorylation of β-catenin at Ser552. This result is in line with our previous study, showing that AMPK phosphorylates β-catenin at Ser552, which stabilises β-catenin( Reference Zhao, Yue and Zhu 57 ).
In conclusion, 0·1 % GSE supplementation reduced cell proliferation, enhanced cell differentiation and improved barrier function in IL10-deficient mice probably through inhibiting the Wnt/β-catenin pathway.
Acknowledgements
The present study was financially supported by the NIHR15HD073864 and Washington State University seed grant to M. J. Z.
The authors' contributions are as follows: G. Y. and M. J. Z. participated in the study design, data acquisition, analysis and interpretation of the data; G. Y., Y. X. and H. Z. conducted the research; G. Y., M. J. Z. and M. D. contributed to the writing and revision of the manuscript.
None of the authors has any conflicts of interest to declare.







