
116 results
New integrated molecular approaches for investigating lake settlements in north-western Europe
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
309 MYC Inhibition Overcomes IMiD Resistance in Heterogeneous Multiple Myeloma Populations
-
- Journal:
- Journal of Clinical and Translational Science / Volume 6 / Issue s1 / April 2022
- Published online by Cambridge University Press:
- 19 April 2022, p. 54
-
- Article
-
- You have access
- Open access
- Export citation
Addressing personal protective equipment (PPE) decontamination: Methylene blue and light inactivates severe acute respiratory coronavirus virus 2 (SARS-CoV-2) on N95 respirators and medical masks with maintenance of integrity and fit
- Part of
-
- Journal:
- Infection Control & Hospital Epidemiology / Volume 43 / Issue 7 / July 2022
- Published online by Cambridge University Press:
- 21 May 2021, pp. 876-885
- Print publication:
- July 2022
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
A conceptual framework to address administrative and infection control barriers for animal-assisted intervention programs in healthcare facilities: Perspectives from a qualitative study
- Part of
-
- Journal:
- Infection Control & Hospital Epidemiology / Volume 43 / Issue 4 / April 2022
- Published online by Cambridge University Press:
- 25 January 2021, pp. 531-533
- Print publication:
- April 2022
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
Validation and calibration of the Eating Assessment in Toddlers FFQ (EAT FFQ) for children, used in the Growing Up Milk – Lite (GUMLi) randomised controlled trial
-
- Journal:
- British Journal of Nutrition / Volume 125 / Issue 2 / 28 January 2021
- Published online by Cambridge University Press:
- 17 August 2020, pp. 183-193
- Print publication:
- 28 January 2021
-
- Article
-
- You have access
- HTML
- Export citation
Part I - Identifying priorities and collating the evidence
-
- Book:
- Conservation Research, Policy and Practice
- Published online:
- 18 April 2020
- Print publication:
- 16 April 2020, pp 27-142
-
- Chapter
-
- You have access
- Open access
- HTML
- Export citation
Chapter One - Making a difference in conservation: linking science and policy
- from Introduction and scene setting
-
-
- Book:
- Conservation Research, Policy and Practice
- Published online:
- 18 April 2020
- Print publication:
- 16 April 2020, pp 3-8
-
- Chapter
-
- You have access
- Open access
- HTML
- Export citation
Chapter Twenty - Successfully translating conservation research into practice and policy: concluding thoughts
- from Conclusion
-
-
- Book:
- Conservation Research, Policy and Practice
- Published online:
- 18 April 2020
- Print publication:
- 16 April 2020, pp 325-328
-
- Chapter
-
- You have access
- Open access
- HTML
- Export citation
Part II - Influencing and making decisions
-
- Book:
- Conservation Research, Policy and Practice
- Published online:
- 18 April 2020
- Print publication:
- 16 April 2020, pp 143-262
-
- Chapter
-
- You have access
- Open access
- HTML
- Export citation
Contents
-
- Book:
- Conservation Research, Policy and Practice
- Published online:
- 18 April 2020
- Print publication:
- 16 April 2020, pp vii-ix
-
- Chapter
-
- You have access
- Open access
- HTML
- Export citation
Contributors
-
- Book:
- Conservation Research, Policy and Practice
- Published online:
- 18 April 2020
- Print publication:
- 16 April 2020, pp x-xvi
-
- Chapter
-
- You have access
- Open access
- HTML
- Export citation
Part III - Communicating the message
-
- Book:
- Conservation Research, Policy and Practice
- Published online:
- 18 April 2020
- Print publication:
- 16 April 2020, pp 263-322
-
- Chapter
-
- You have access
- Open access
- HTML
- Export citation
Introduction and scene setting
-
- Book:
- Conservation Research, Policy and Practice
- Published online:
- 18 April 2020
- Print publication:
- 16 April 2020, pp 1-26
-
- Chapter
-
- You have access
- Open access
- HTML
- Export citation
Plate Section (PDF Only)
-
- Book:
- Conservation Research, Policy and Practice
- Published online:
- 18 April 2020
- Print publication:
- 16 April 2020, pp 337-356
-
- Chapter
-
- You have access
- Open access
- HTML
- Export citation

Conservation Research, Policy and Practice
-
- Published online:
- 18 April 2020
- Print publication:
- 16 April 2020
-
- Book
-
- You have access
- Open access
- Export citation
Copyright page
-
- Book:
- Conservation Research, Policy and Practice
- Published online:
- 18 April 2020
- Print publication:
- 16 April 2020, pp vi-vi
-
- Chapter
-
- You have access
- Open access
- HTML
- Export citation
Conclusion
-
- Book:
- Conservation Research, Policy and Practice
- Published online:
- 18 April 2020
- Print publication:
- 16 April 2020, pp 323-328
-
- Chapter
-
- You have access
- Open access
- HTML
- Export citation
Index
-
- Book:
- Conservation Research, Policy and Practice
- Published online:
- 18 April 2020
- Print publication:
- 16 April 2020, pp 329-336
-
- Chapter
-
- You have access
- Open access
- HTML
- Export citation
Deformation of ambient chemical gradients by sinking spheres
-
- Journal:
- Journal of Fluid Mechanics / Volume 892 / 10 June 2020
- Published online by Cambridge University Press:
- 08 April 2020, A33
-
- Article
-
- You have access
- Open access
- HTML
- Export citation
-
A sphere sinking through a chemical gradient drags fluid with it, deforming the gradient. The sphere leaves a trail of gradient enhancement that persists longer than the velocity disturbance in the Reynolds
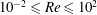 $10^{-2}\leqslant Re\leqslant 10^{2}$, Froude
$10^{-2}\leqslant Re\leqslant 10^{2}$, Froude 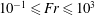 $10^{-1}\leqslant Fr\leqslant 10^{3}$ and Péclet
$10^{-1}\leqslant Fr\leqslant 10^{3}$ and Péclet 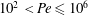 $10^{2}<Pe\leqslant 10^{6}$ regime considered here. We quantify the enhancement of the gradient and the diffusive flux in the trail of disturbed chemical left by the passing sphere using a combination of numerical simulations and scaling analyses. When
$10^{2}<Pe\leqslant 10^{6}$ regime considered here. We quantify the enhancement of the gradient and the diffusive flux in the trail of disturbed chemical left by the passing sphere using a combination of numerical simulations and scaling analyses. When 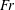 $Fr$ is large and buoyancy forces are negligible, dragged isosurfaces of chemical form a boundary layer of thickness
$Fr$ is large and buoyancy forces are negligible, dragged isosurfaces of chemical form a boundary layer of thickness 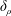 $\unicode[STIX]{x1D6FF}_{\unicode[STIX]{x1D70C}}$ around the sphere with diameter
$\unicode[STIX]{x1D6FF}_{\unicode[STIX]{x1D70C}}$ around the sphere with diameter 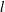 $l$. We derive the scaling
$l$. We derive the scaling 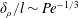 $\unicode[STIX]{x1D6FF}_{\unicode[STIX]{x1D70C}}/l\sim \mathit{Pe}^{-1/3}$ from the balance of advection and diffusion in the chemical boundary layer. The sphere displaces a single isosurface of chemical a maximum distance
$\unicode[STIX]{x1D6FF}_{\unicode[STIX]{x1D70C}}/l\sim \mathit{Pe}^{-1/3}$ from the balance of advection and diffusion in the chemical boundary layer. The sphere displaces a single isosurface of chemical a maximum distance 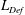 $\mathit{L}_{Def}$ that increases as
$\mathit{L}_{Def}$ that increases as 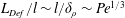 $\mathit{L}_{Def}/l\sim l/\unicode[STIX]{x1D6FF}_{\unicode[STIX]{x1D70C}}\sim \mathit{Pe}^{1/3}$. Increased flux through the chemical boundary layer moving with the sphere is described by a Sherwood number,
$\mathit{L}_{Def}/l\sim l/\unicode[STIX]{x1D6FF}_{\unicode[STIX]{x1D70C}}\sim \mathit{Pe}^{1/3}$. Increased flux through the chemical boundary layer moving with the sphere is described by a Sherwood number, 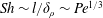 $Sh\sim l/\unicode[STIX]{x1D6FF}_{\unicode[STIX]{x1D70C}}\sim \mathit{Pe}^{1/3}$. The gradient enhancement trail extends much farther than
$Sh\sim l/\unicode[STIX]{x1D6FF}_{\unicode[STIX]{x1D70C}}\sim \mathit{Pe}^{1/3}$. The gradient enhancement trail extends much farther than 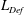 $\mathit{L}_{Def}$ as displaced isosurfaces slowly return to their original positions through diffusion. In the reference frame of a chemical isosurface moving past the sphere, a new quantity describing the Lagrangian flux is found to scale as
$\mathit{L}_{Def}$ as displaced isosurfaces slowly return to their original positions through diffusion. In the reference frame of a chemical isosurface moving past the sphere, a new quantity describing the Lagrangian flux is found to scale as 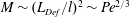 $\mathit{M}\sim (\mathit{L}_{Def}/l)^{2}\sim \mathit{Pe}^{2/3}$. The greater
$\mathit{M}\sim (\mathit{L}_{Def}/l)^{2}\sim \mathit{Pe}^{2/3}$. The greater 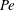 $\mathit{Pe}$ dependence of
$\mathit{Pe}$ dependence of 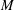 $\mathit{M}$ versus
$\mathit{M}$ versus 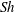 $Sh$ demonstrates the importance of the deformation trail for determining the total flux of chemical in the system. For
$Sh$ demonstrates the importance of the deformation trail for determining the total flux of chemical in the system. For 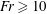 $\mathit{Fr}\geqslant 10$, buoyancy forces are weak compared to the motion of the sphere and the preceding results are retained. Below
$\mathit{Fr}\geqslant 10$, buoyancy forces are weak compared to the motion of the sphere and the preceding results are retained. Below 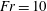 $\mathit{Fr}=10$, an additional Froude dependence is found and
$\mathit{Fr}=10$, an additional Froude dependence is found and 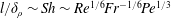 $l/\unicode[STIX]{x1D6FF}_{\unicode[STIX]{x1D70C}}\sim Sh\sim Re^{1/6}Fr^{-1/6}Pe^{1/3}$. Buoyancy forces suppress gradient deformation downstream, resulting in
$l/\unicode[STIX]{x1D6FF}_{\unicode[STIX]{x1D70C}}\sim Sh\sim Re^{1/6}Fr^{-1/6}Pe^{1/3}$. Buoyancy forces suppress gradient deformation downstream, resulting in 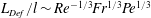 $\mathit{L}_{Def}/l\sim Re^{-1/3}Fr^{1/3}Pe^{1/3}$ and
$\mathit{L}_{Def}/l\sim Re^{-1/3}Fr^{1/3}Pe^{1/3}$ and 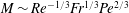 $\mathit{M}\sim Re^{-1/3}Fr^{1/3}Pe^{2/3}$. The productivity of marine plankton – and therefore global carbon and oxygen cycles – depends on the availability of microscale gradients of chemicals. Because most plankton exist in the fluids regime under consideration, this work describes a new mechanism by which sinking particles and plankton can stir weak ambient chemical gradients a distance
$\mathit{M}\sim Re^{-1/3}Fr^{1/3}Pe^{2/3}$. The productivity of marine plankton – and therefore global carbon and oxygen cycles – depends on the availability of microscale gradients of chemicals. Because most plankton exist in the fluids regime under consideration, this work describes a new mechanism by which sinking particles and plankton can stir weak ambient chemical gradients a distance 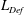 $\mathit{L}_{Def}$ and increase chemical flux in the trail by a factor of
$\mathit{L}_{Def}$ and increase chemical flux in the trail by a factor of 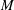 $\mathit{M}$.
$\mathit{M}$.
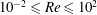
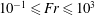
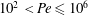
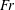
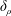
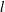
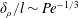
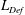
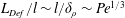
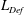
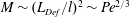
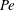
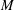
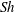
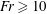
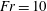
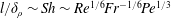
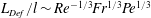
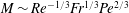
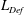
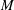
Physiological responses to maximal eating in men
-
- Journal:
- British Journal of Nutrition / Volume 124 / Issue 4 / 28 August 2020
- Published online by Cambridge University Press:
- 06 April 2020, pp. 407-417
- Print publication:
- 28 August 2020
-
- Article
-
- You have access
- HTML
- Export citation
