



Introduction
Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae), or cotton bollworm, is one of the important pests of many plants that are important in agriculture. This insect larva is a polyphagous pest and feeds on more than 200 types of agricultural products, including soybean, cotton, chickpea, peas, tobacco, corn, and a number of other crops and vegetables. The larvae show a high tendency to feed on the reproductive organs of these plants, which has a significant effect on reducing their production. The use of chemical pesticides is the simplest and least costly method of control in cotton bollworm, but especially in recent years, the use of pesticides has caused many problems, including environmental pollution, human health threats, and pesticide resistance. Alternative bollworm-control methods that reduce the use of chemical pesticides in agricultural lands and protect natural enemies are of interest to researchers (Lawo et al. Reference Lawo, Mahon, Milner, Sarmah, Higgins and Romeis2008).
Hormones play an important role in controlling insect reproduction. Juvenile hormone and ecdysteroids play an important role in their physiology (Franssens et al. Reference Franssens, Smagghe, Simonet, Claeys, Breugelmans, De Loof and Broeck2006). These hormones are involved in regulating the physiological processes throughout the life cycle of insects, especially the reproductive system, which produces vitellogenin and oocyte maturation. Ecdysteroid regulates the reproductive system, including oocyte maturation and vitellogenesis. (Khalid et al. Reference Khalid, Ahmad, Ngegba and Zhong2021). Insect growth regulators that mimic the activity of the juvenile hormone increase larval age. In addition, Sorge et al. (Reference Sorge, Nauen, Range and Hoffmann2000) found that pseudo-hormones sterilise insects and disrupt reproductive behaviour. The application of pyriproxyfen, a juvenile hormone analogue, on adult female Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae) could decrease reproduction. It reduced oviposition, shortened life span, and affected the expression of vitellogenin and ecdystroied receptors in S. litura (Xu et al. Reference Xu, Tang, Zou, Zheng, Liu and Wang2015). Based on this evidence, the use of hormone analogues can interfere with the reproduction of insects (Xu et al. Reference Xu, Tang, Zou, Zheng, Liu and Wang2015). The insect growth regulators, pyriproxyfen, flufenoxuron, and chlorfluazuron, were evaluated on various reproductive aspects of Spodoptera littoralis (Boisduval) (Lepidoptera: Noctuidae) (Xu et al. Reference Xu, Tang, Zou, Zheng, Liu and Wang2015). Toxicity and morphological deformities induced by the insect growth regulators were seen. These insect growth regulators have potential for controlling S. littoralis (Shaurub et al. Reference Shaurub, Abdel Aal and Emara2020). Moreover, in a study by Martínez et al. (Reference Martínez, Plata-Rueda and Serrão2021), fenoxycarb, methoxyfenozide, and tebufenozide were found to be effective against Euprosterna elaeasa (Dyar) (Lepidoptera: Limacodidae), inhibiting feeding in larvae, with less leaf area consumed, thereby indicating potential for controlling this pest.
Mahmoudvand and Moharramipour (Reference Mahmoudvand and Moharramipour2015) showed that fenoxycarb, a juvenile hormone analogue, decreased fertility of Plutella xylostella (Linnaeus) (Lepidoptera: Plutellidae).
In the present study, the effects of fenoxycarb on fecundity, fertility, ecdysteroid level, oocyte size, and expression levels of the USP, EcR, HR3, and Vg genes in adult female H. armigera were examined.
Materials and methods
Insect rearing
Cotton bollworm larvae were collected in cotton field in Gorgan, Iran (36° 50′ 19″ N,54° 26′ 05″ E) and were reared in the laboratory of the entomology department of Tarbiat Modares University, Tehran, Iran, using artificial diet (800 mL distilled water, 40 g yeast powder, 260 g chickpea powder, 4 g ascorbic acid, 1.5 g sorbic acid, 1 mL formaldehyde (40%), 2.5 g Nipagin, 0.115 g cholesterol, and 12 g agar), per Shorey and Hale (Reference Shorey and Hale1965). The larvae were maintained at 25 ± 2 °C, 16:8-hour light:dark photoperiod, and 65 ± 5% relative humidity. Forty newly emerges adult female and male (1:1) were released in a cage (20 × 20 × 30 cm) to mate. Cotton soaked in a 10% (vol/vol) honey solution was provided for the adult moths as food.
Fenoxycarb treatment
Fenoxycarb, a juvenile hormone analogue insecticide, was supplied by Sigma-Aldrich (St. Louis, Missouri, United States of America; catalogue number 34343) and dissolved in a water and acetone (1:1) solution. Third-instar larvae were fed an artificial diet (1 mg) that contained 100 µL of fenoxycarb at doses of LC10 (3.42 mg/L), LC25 (5.53 mg/L), and LC50 (9.2 mg/L; lethal dose). Control larvae were allowed to feed on artificial diet containing water and acetone (1:1) in the same ratio as that provided in the fenoxycarb treatments. The experiment was repeated three times, and each replication contained 10 third-instar larvae, each of which was kept in a separate plastic box (7 cm3). Mortality was recorded after 10 days of exposure to the insecticide-laced food, as noted in the treatments above.
Determining fenoxycarb effects on fecundity, fertility, and longevity parameters
One hundred young third-instar larvae were selected randomly and fed individually on diet containing fenoxycarb (doses: LC10 and LC25) for 10 days. Surviving larvae were then collected and reared on artificial diet and monitored daily, and larval mortality and pupal weight were determined. After adult emergence, one male and one female were transferred to a cylindrical container, and their fecundity, fertility, and longevity parameters were determined. The adults were fed on a 10% (v:v) honey solution in water. The experiments to determine the reproductive parameters were carried out with 15 replications for each treatment. The number of eggs laid by each female was counted daily until the female died. The longevity of each male and female was also recorded.
Measuring oocyte length
To determine the effect of fenoxycarb on the size of the insect’s oocytes, terminal oocytes (the largest oocyte is located at the base of the ovarioles) were measured according to the method of Van Wielendaele et al. (Reference Van Wielendaele, Wynant, Dillen, Badisco, Marchal and Vanden Broeck2013) in 10 females. A piece of millimetre-squared paper was used to calculate the average oocyte size.
Ecdysteroid extraction and quantification
To determine the concentration of ecdysteroid in ovaries, the enzyme immune assay method (Cayman Chemicals, Inc., Ann Arbor, Michigan, United States of America) was used (Ahmadi et al. Reference Ahmadi, Mikani and Moharramipour2021; Hackney et al. Reference Hackney, Zolali-Meybodi and Cherbas2012).
Ecdysteroid extraction
The assay is based on competition between a 20-hydroxyecdysone-acetylcholinesterase (AChE) conjugate (20E Tracer) and 20-hydroxyecdysone for a limited number of 20E-specific rabbit antiserum–binding sites.
The ovaries extracted from each female H. armigera were homogenised with 100 µL methanol, followed by centrifugation at 15 000 × g, 4 °C for 15 minutes. The supernatant was removed and poured into a separate tube. This step was repeated once. The supernatant was then placed in a Speed Vac machine to evaporate the solvent, then 1000 µL enzyme immunoassay buffer (1 mM ethylenediaminetetraacetic acid (EDTA), 0.4 M NaCl, and 0.1% bovine serum albumin in 0.1 M phosphate buffer) was added to the samples and kept at –20 °C for further experiment.
Ecdysteroid quantification
To determine the ecdysteroid concentration in each well, 50 µL of 20E-acetylcholinesterase (AchE) tracer, 100 µL of enzyme immune assay buffer, 50 µL of 20E enzyme immune assay antiserum (Cayman Chemicals, Inc.), and 50 µL of sample solution extracted from ovaries, per the above, were poured and placed on a shaker at 4 °C for 18 hours. The plate was then emptied and rinsed six times with a washing buffer to remove any unbound reagents. Then, 200 μL of Ellman’s reagent (which contains the substrate to AChE) was added to each well for development. Then, 5 µL of tracer (acetylcholinesterase linked to 20E) was added, and finally, the plate was covered and placed on the shaker for 90–120 minutes.
The product of this enzymatic reaction has a distinct yellow colour, determined spectrophotometrically, that is proportional to the amount of 20E tracer bound to the well, which is inversely proportional to the amount of free 20E present in the well during incubation. Using a microplate reader spectrometer (BioTek, Winooski, Vermont, United States of America), the absorbance was read at 412 nm.
Real-time quantitative reverse transcription–polymerase chain reaction
Ribonucleic acid samples were extracted from the fat bodies of female H. armigera using TRIzol solution, according to the manufacturer’s instructions (Molecular Research Center, Cincinnati, Ohio, United States of America), then 0.2 mL of chloroform per 1 mL of TRIzol Reagent (Molecular Research Center Inc., Cincinnati, Ohio, United States of America) was added. The tubes were shaken vigorously by hand for 15 seconds and then incubated at 15–30 °C for 2 to 3 minutes. The samples were centrifuged at 12 000 × g for 15 minutes at 2–8 °C, which separated the mixture into a lower red, phenol-chloroform phase, an interphase, and a colourless upper-aqueous phase, with the RNA remaining exclusively in the aqueous phase. The aqueous phase was transferred to a fresh tube. The RNA from the aqueous phase was precipitated by mixing with 0.5 mL of isopropyl alcohol per 1 mL of TRIzol Reagent for the initial homogenisation. Samples were incubated at 15–30 °C for 10 minutes, followed by centrifugation at 12 000 × g for 10 minutes at 2–8 °C. The supernatant was removed, and the RNA pellet was washed once with 75% ethanol, adding at least 1 mL of 75% ethanol per 1 mL of the TRIzol Reagent used for the initial homogenisation. The sample was mixed by vortexing and centrifuged at 7500 × g for 5 minutes at 2–8 °C. At the end of the procedure, the RNA pellet was dried.
The concentration of the purified RNA was measured using a nanodrop spectrophotometer. One microlitre total RNA (containing 1 ng–5 µg) and 1 μL oligo (dT) were added to sterile nuclease-free water for 5 minutes to create a total volume of 13.41 μL on ice. This mixture was incubated for 5 minutes at 65 °C and chilled on ice. Then 4 μL M-MLV, 1 μL M-MLV, and 1 μL deoxyribonucleoside 5′-triphosphates (dNTPs) were added to the solution and incubated for 60 minutes at 55 °C, followed by incubation for 5 minutes at 85 °C. The primers used for real-time quantitative reverse transcription–polymerase chain reaction were designed using the Primer3 software. The real-time quantitative reverse transcription–polymerase chain reaction was performed to determine transcription levels of the vitellogenin gene (Vg; GenBank accession number: JQ723600), the nuclear hormone receptor, ultraspiracle (USP; GenBank accession number: EU526832.1), the ecdysone receptor (EcR; GenBank accession number: EU526831.1), and hormone receptor 3 (HR3; GenBank accession number: AF337637.3), using specific primers and Actin as reference genes (Table 1). Twenty nanograms of total genomic DNA were subjected to quantitative polymerase chain reaction as a template, using SYBR Green (Ampliqon, Odese, Denmark) with a Mic-qPCR cycler (Bio Molecular Systems, Upper Coomera, Queensland, Australia). Polymerase chain reaction conditions were as follows: 95 °C for 10 minutes, followed by 40 cycles of 95 °C for 15 seconds and 60 °C for 60 seconds. All the experiments repeated three times, with three biological replicates.
Table 1. Primers used in the present study.

Statistical analysis
One-way analysis of variance, followed by Tukey’s test, was used to compare the data (Tables 2, 3, 4, and 5; Figs. 1, 2, 3, and 4). Quantitative polymerase chain reaction data were analysed using the 2−ΔΔCT method (Livak and Schmittgen Reference Livak and Schmittgen2001). Student’s t-test was applied. Graphs were created with PRISM, version 7 (GraphPad, San Diego, California, United States of America).
Table 2. Effects of sublethal doses of fenoxycarb (doses: LC10 and LC25) on the larval survival rate, pupation, and adult emergence of Helicoverpa armigera. Means followed by the same letters within a column are not significantly different (Tukey’s test; P < 0.05). The values with different superscript letters in a column are significantly different (P < 0.05).

Table 3. Effects of fenoxycarb (doses: LC10 and LC25) on the pupal weight, fecundity (eggs/female), and fertility (fertile eggs/female) of Helicoverpa armigera. Means followed by the same letters within a column are not significantly different (Tukey’s test; P < 0.05). The values with different superscript letters in a column are significantly different (P < 0.05).

Table 4. Effects of sublethal fenoxycarb (doses: LC10 and LC25) on the preoviposition period, oviposition period (day), postoviposition period (day), and peak oviposition period (day) of Helicoverpa armigera. Means followed by the same letters within a column are not significantly different (Tukey’s test; P < 0.05). The values with different superscript letters in a column are significantly different (P < 0.05).

Table 5. Effects of sublethal fenoxycarb (doses: LC10 and LC25) on pupa longevity and female longevity of Helicoverpa armigera. Means followed by the same letters within a column are not significantly different (Tukey’s test; P < 0.05). The values with different superscript letters in a column are significantly different (P < 0.05).


Figure 1. Effects of sublethal doses, LC10 (3.42 mg/L) and LC25 (5.53 mg/L), of fenoxycarb on oocyte size in adult female Helicoverpa armigera. One-way analysis of variance and Tukey’s test were used to compare data (** P < 0.01).

Figure 2. Effects of sublethal doses, LC10 (3.42 mg/L) and LC25 (5.53 mg/L), of fenoxycarb on ecdysteroid level in the ovaries of adult female Helicoverpa armigera. Ecdysteroids concentration was measured using enzyme immune assays. One-way analysis of variance and Tukey’s test were used to compare data. Asterisks indicate significant differences (** P < 0.01), and “n.s.” indicates no significant difference.

Figure 3. Effects of sublethal doses, LC10 (3.42 mg/L) and LC25 (5.53 mg/L), of fenoxycarb on EcR, HR3, USP, and Vg gene expression levels in pupal Helicoverpa armigera. Actin was used as reference gene. Each experiment was replicated three times with three biological replicates. The levels of EcR, HR3, USP, and Vg gene expression were analysed. One-way analysis of variance and Tukey’s test were used to compare data. Asterisks indicate significant differences (***P < 0.001, ** P < 0.01, * P < 0.05), and “n.s.” indicates no significant difference.

Figure 4. Effects of sublethal doses, LC10 (3.42 mg/L) and LC25 (5.53 mg/L), of fenoxycarb on EcR, HR3, USP, and Vg gene expression levels in adult female Helicoverpa armigera. Actin was used as reference gene. Each experiment was replicated three times with three biological replicates. The levels of EcR, HR3, USP, and Vg gene expression were analysed. One-way analysis of variance and Tukey’s test were used to compare data. Asterisks indicate significant differences (***P < 0.001, **P < 0.01, *P < 0.05), and “n.s.” indicates no significant difference.
Mortality data from the larval toxicity bioassays were analysed to calculate LC10, LC25, and LC50 values, with 95% confidence limits, slope, and intercept, using the Probit procedure of the SPSS programme, version 23, developed by IBM (Armonk, New York, United States of America).
Results
Toxicity of fenoxycarb
The data presented in Table 6 indicate that the fenoxycarb clearly had high toxicity against H. armigera. The estimated LC90 and LC50 doses were 25.99 and 9.12 mg/L after 10 days, respectively. Also, the LC10 and LC25 values were 3.42 and 5.53 mg/L after 10 days, respectively (Table 6).
Table 6. Toxicity of fenoxycarb on Helicoverpa armigera, 10 days after feeding on artificial diet containing the insecticide. LCL, lower confidence limit, at 95%; UCL, upper confidence limit, at 95%.
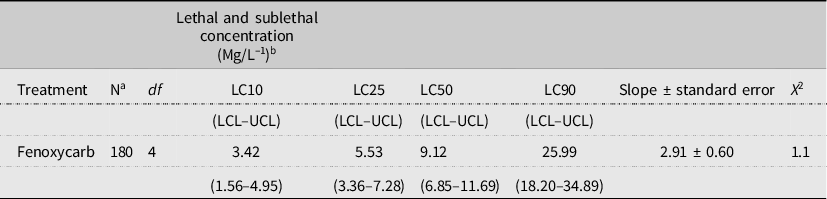
a Number of larvae.
b 95% confidence limits in parenthesis.
Effects of sublethal dose of fenoxycarb on larval survival rate, pupation, and adult emergence
Feeding on artificial diet that contained fenoxycarb (doses: LC10 and LC25) significantly decreased the percentage of larval survival rate (F = 9.95; df = 2, 297; P < 0.0001). The sublethal doses (at LC10 and LC25) of fenoxycarb caused losses of almost 15.8 and 23.1% of the larval survival rate, respectively. In addition, the pupation rate (F = 14.80; df = 2, 297; P < 0.0001) significantly decreased, as did the adult emergence rate (F = 13.33; df = 2, 297; P < 0.0001). Fenoxycarb at the LC10 and LC25 doses elicited a decrease of about 20.5 and 32.6% in pupation rate, respectively. Moreover, losses of 18.7 and 35.9% were observed in adult emergence when the insecticide was applied at LC10 and LC25 concentrations, respectively (Table 2).
Effects of sublethal dose of fenoxycarb on pupal weight, fecundity (eggs/female), and fertility (fertile eggs/female)
Pupal weight decreased by 7.7 and 8.7% with the fenoxycarb LC10 and LC25 treatments, respectively, compared to the control (F = 10.44; df = 2, 87; P < 0.0001). Fecundity (F = 4.87; df = 2, 42; P < 0.012) and fertility (F = 24.51; df = 2, 42; P < 0.0001) were also affected by the insecticide. Fenoxycarb at the LC25-dose treatment clearly reduced fecundity and fertility by about 23.2 and 46.3%, respectively (Table 3).
Effects of sublethal dose of fenoxycarb on preoviposition period (day), oviposition period (day), postoviposition period (day), and peak oviposition period (day)
The preoviposition period and oviposition period of H. armigera were significantly affected by fenoxycarb. The LC25 treatment significantly increased the preoviposition period by about 39% (F = 6.29; df = 2, 42; P < 0.004), but it clearly decreased oviposition period by about 31.3% (F = 30.60; df = 2, 42; P < 0.0001). The postoviposition period (day) and the peak oviposition period (day) were unaffected by the insecticide (Table 4).
Effects of sublethal dose of fenoxycarb on pupa longevity (day) and female longevity (day)
Feeding on artificial diet that contained fenoxycarb (doses: LC10 and LC25) significantly increased pupal longevity in H. armigera (F = 20.004; df = 2, 87; P < 0.0001) but shortened female longevity (F = 15.33; df = 2, 87; P < 0.0001). The LC25 treatment increased pupa longevity by about 15.81% but decreased female longevity by about 23.02% (Table 2).
Effects of sublethal dose of fenoxycarb on oocyte size
To investigate the effect of the sublethal doses of fenoxycarb on oocyte size, third-instar larvae were fed individually on artificial diet containing fenoxycarb (doses: LC10 and LC25) for 10 days. After adult emergence, the oocyte size was measured. The results showed that feeding on diet containing fenoxycarb clearly decreased oocyte size compared to the control. The LC25 fenoxycarb treatment decreased oocyte size by about 28.1% (Fig. 1).
Effects of sublethal dose of fenoxycarb on ecdysteroid concentration in the ovaries
Third-instar larvae of H. armigera were fed on diet containing sublethal doses of fenoxycarb for 10 days. The ecdysteroid concentration in the ovaries of the resulting adult females was measured using enzyme immune assay. The results show that larvae fed diet containing 5.53 mg/L (the LC25 treatment) of fenoxycarb lead to adult female ovaries having a significantly lower ecdysteroid concentration than did ovaries of adult female H. armigera raised as the control. The ecdysteroid concentration in ovaries of the control was approximately 1.5 times that found in the ovaries of the insects fed the fenoxycarb-containing (LC25) diet (Fig. 2).
Sublethal effects of fenoxycarb on Vg, USP, EcR, and HR3 gene expression levels in pupae and adults of H. armigera
To understand the contribution of fenoxycarb to vitellogenesis in H. armigera, the insect was fed on diet containing the insecticide. The expression of EcR, USP, HR3, and Vg genes in the fat body dissected from both fenoxycarb-treated and control insects were investigated at 24, 48, and 72 hours after treatment. As shown in Figures 3 and 4, the fenoxycarb treatment suppressed the expression of the USP, EcR, and Vg genes in both pupal and adult H. armigera. A similar expression profile was observed for EcR and USP genes in pupa, but a different expression profile was observed in the adult insects. However, relative expressions of the EcR gene were 3.22- and twofold lower in fenoxycarb-treated (LC25) adult and pupa after 72 hours, respectively, compared to those in control insects (Figs. 3 and 4). Although relative expression of the USP gene was not downregulated after 48 hours in fenoxycarb-treated (LC25) pupa, it became threefold lower after 72 hours, compared to the control. In the case of adult insects, it was 2.72-fold lower compared to that in the control after 48 hours. That is, although the level of USP expression was similar between the fenoxycarb-treated and control adult in the last 24 hours of the experiment, it significantly decreased in the fenoxycarb-treated insects during the first 48 hours of the observation period compared to the controls (Fig. 4).
Discussion
In this study, the effects of the insecticide fenoxycarb on the female reproductive system of H. armigera were determined. Many studies confirm the toxicity of fenoxycarb on some insects. Mahmoudvand and Moharramipour (Reference Mahmoudvand and Moharramipour2015) tested its effects on some biological parameters of Plutella xylostella and found the calculated LC10 and LC25, and LC50 values of the insecticide were 21.58 and 43.25 mg/L on third-instar larvae of that insect, respectively. Martínez et al. (Reference Martínez, Plata-Rueda and Serrão2021) evaluated potential of fenoxycarb to control E. elaeasa, assessing the insecticide against E. elaeasa larvae for toxicity, feeding inhibition, and mortality. Those authors found that the LC50 of fenoxycarb was 0.199 g/L−1.
In the present study, we report the sublethal effects of fenoxycarb on H. armigera. In previous studies, fenoxycarb’s effects on fecundity varied. For example, Horton and Lewis (Reference Horton and Lewis1996) found the chemical enhanced the fecundity in Cacopsylla pyricola (Foerster) (Hemiptera: Psyllidae), whereas the present study shows that fenoxycarb decreased fecundity and fertility in H. armigera. Zhang et al. (Reference Zhang, Wang, Liu, Zhao, Jin, Zhou and Guo2019) showed that silencing the Vg gene decreased fecundity and ovary development in Agasicles hygrophila (Selman) (Coleoptera: Chrysomelidae), but in the present study, we found that fenoxycarb decreases Vg expression, thereby inhibiting fertility and egg hatching rates.
Treatment of the larvae with fenoxycarb in the present study caused H. armigera pupae to be underweight compared to the controls. This result is consistent with those of studies of Mahmoudvand and Moharramipour (Reference Mahmoudvand and Moharramipour2015) and Biddinger and Hull (Reference Biddinger and Hull1999). The present study’s findings are also consistent with those of Mahmoudvand and Moharramipour (Reference Mahmoudvand and Moharramipour2015), who reported that LC10 and LC50 fenoxycarb affects longevity, fecundity, fertility, and pupa weight in P. xylostella. Moreover, those authors reported that fenoxycarb did not affect the postoviposition period in P. xylostella, and Xu et al. (Reference Xu, Tang, Zou, Zheng, Liu and Wang2015) showed no significant difference in the ovipositing peak of S. littoralis when different concentrations of pyriproxyfen were applied. The present study found similar results.
In the present study, fenoxycarb significantly increased the preoviposition period but shortened the oviposition period, compared to the control. Yusoff et al. (Reference Yusoff, Abd Ghani, Wahida Othman, Mohd Aizat and Hassan2021) observed similar effects were observed with farnesyl acetate decreasing oviposition in P. xyllostella. According to Yusoff et al. (Reference Yusoff, Abd Ghani, Wahida Othman, Mohd Aizat and Hassan2021), decreasing oviposition time may led to decreases in fecundity.
We demonstrated that fenoxycarb decreased the size of H. armigera oocytes. It also decreased the level of ecdysteroid within the ovaries. Previously, Bai et al. (Reference Bai, Gelman and Pallia2010) reported that methoprene, a juvenile hormone analogue, inhibited ecdysteroid biosynthesis in Anopheles gambiae (Giles) (Diptera: Culicidae), and Rountree and Bollenbacher (Reference Rountree and Bollenbacher1986) reported that juvenile hormone or its analogues inhibited ecdysteroid secretion in some lepidopterous insects. In the present study, relative expression levels of the EcR gene decreased after H. armigera larvae fed on diet containing fenoxycarb. It may be possible that the low level of EcR expression led to a decrease in the length of the oocytes. Previously, Gujar and Palli (Reference Gujar and Palli2016) reported that knockdown of the EcR gene decreased the oocyte size in Cimex lectularius (Linnaeus) (Hemiptera: Cimicidae).
In the present study, real-time quantitative reverse transcription–polymerase chain reaction was used to understand the effect of fenoxycarb on the relative expression levels of the EcR, USP, HR3, and Vg genes. The results showed that relative expression levels of EcR and USP were lower in fenoxycarb-treated insects than in their controls. Hossain Ahmed et al. (Reference Hossain Ahmed, Saunders, Mullins, Rahman and Zhu2020) showed that pyriproxyfen considerably decreased the expression of the EcR, HR3, and Vg genes in Aedes aegypti (Linnaeus) (Diptera: Culicidae), and Bai et al. (Reference Bai, Gelman and Pallia2010) reported that the transcription of EcR and HR3 is downregulated in methoprene-treated A. gambiae, possibly due to the inhibition of 20E action by insecticide. The EcR and USP genes combine to generate a heterodimer, which 20E can combine with to regulate the expression levels of downstream genes and thus affect the ovary development (Qian et al. Reference Qian, Zhang, Zhao, Guo, Li and Xu2020). However, in the present study, fenoxycarb was found to have no effect on HR3, which is an important downstream hormone. This result means that HR3 is not involved in vitellogenesis in H. armigera.
Our results show that fenoxycarb decreased Vg expression. Previously, Bai et al. (Reference Bai, Gelman and Pallia2010) reported that methoprene suppressed expression of Vg in A. gambiae by 30%. This may be a result of the insecticide effects on ecdysteroid levels. 20-Hydroxyecdysone-acetylcholinesterase can play an important role in regulating vitellogenesis (Sun et al. Reference Sun, Song and Barrett2003). Fenoxycarb decreased the 20E levels, which led to decreased Vg gene expression.
We showed that gene expression levels were affected, according to different concentrations of fenoxycarb. This finding is consistent with the results of Xu et al. (Reference Xu, Tang, Zou, Zheng, Liu and Wang2015) regarding the influence of pyriproxyfen on reproductive system in S. litura, which showed that the effects of pyriproxyfen on Vg and USP gene expression levels were dose dependent.
In conclusion, feeding H. armigera larvae a diet containing fenoxycarb decreased ecdysteroid titre in ovaries and the expression levels of EcR, USP, and Vg, which led to decreases in fecundity, fertility, and oocyte size. Further study is needed to clarify fenoxycarb’s effects on the reproductive system in the adult female H. armigera.
Acknowledgements
The authors thank Dr. Mohammad Mehrabadi for his assistance.
Competing interests
The authors declare they have no competing interests.










