


Introduction
Developing methods that accurately estimate the abundance or density of animal populations is of central importance for management of threatened species. Complete population counts are resource intensive, rarely feasible because of logistical constraints, and have poor theoretical support (Cochran, Reference Cochran1977; Lancia et al., Reference Lancia, Nichols, Pollock and Bookhout1994). The preferred approach is to sample the population. Typically, sampling is cheaper and provides measures of precision for estimated parameters (Williams et al., Reference Williams, Nichols and Conroy2002).
When using a sampling approach one must first distinguish between the observed or detected number of animals and the true population abundance. The central concept is of detection probability; i.e. only a fraction of all animals present are typically detected during a survey. If we estimate this fraction we can estimate the true population abundance. Estimating detection probability usually requires collecting ancillary data during the course of a survey (Lancia et al., Reference Lancia, Nichols, Pollock and Bookhout1994; MacKenzie et al., Reference MacKenzie, Nichols, Lachman, Droege, Andrew Royle and Langtimm2002). Several sampling methods have been developed that estimate detection probability and population abundance (e.g. line transect sampling, point count surveys, variable circular plots, and capture–recapture).
Given the several sampling techniques available it can be a challenge to determine the most appropriate method. The choice typically depends on field logistics, the ecology of the study species and available resources (Lancia et al., Reference Lancia, Nichols, Pollock and Bookhout1994). For example, species that are difficult to observe directly may require camera trapping or track surveys (Silveira et al., Reference Silveira, Jacomo and Diniz-Filho2003; Lyra-Jorge et al., Reference Lyra-Jorge, Ciocheti, Pivello and Meirelles2008). For highly visible animals in relatively open habitat, direct count-based methods such as distance sampling are often appropriate. When animals are not readily observable, capture–recapture techniques are preferable. These require that animals be individually recognizable (e.g. by tags, natural markings, or DNA identification). For animals without individual markings, distance sampling (Buckland et al., Reference Buckland, Anderson and Burnham2001) or a random encounter model method (REM; Rowcliffe et al., Reference Rowcliffe, Field, Turvey and Carbone2008) may be appropriate to estimate density.
Distance sampling is widely used for estimating abundances of wild animals (Buckland et al., Reference Buckland, Anderson and Burnham2001). For large-bodied, group-living mammals line transects are commonly used (Calambokidis & Barlow, Reference Calambokidis and Barlow2004; Ogutu et al., Reference Ogutu, Bhola, Piepho and Reid2006; Buckland et al., Reference Buckland, Plumptre, Thomas and Rexstad2010), and are suitable for large, highly visible ungulates in African savannahs (Ogutu et al., Reference Ogutu, Bhola, Piepho and Reid2006). Observers move along one or more lines and count groups of animals detected and calculate perpendicular distances to groups (Buckland et al., Reference Buckland, Anderson and Burnham2001). Because not all groups may be detected count data are adjusted according to the probability of detection (Thomas et al., Reference Thomas, Buckland, Burnham, Anderson, Laake, Borchers, Strindberg, El Shaarawi and Piegorsch2002), which is directly related to the observed distribution of distances of groups from the line. This distribution may be modelled using various functions. These functions, and consequently the precision of the resulting density estimates, may be improved through a number of key functions and adjustment terms (Thomas et al., Reference Thomas, Buckland, Burnham, Anderson, Laake, Borchers, Strindberg, El Shaarawi and Piegorsch2002) from which the best-fitting model can be chosen (Burnham & Anderson, Reference Burnham and Anderson2002). For a review, see Buckland et al. (Reference Buckland, Anderson and Burnham2001).
When individuals can be recognized or marked, capture–recapture methods can be used to model population parameters (Otis et al., Reference Otis, Burnham, White and Anderson1978; Amstrup et al., Reference Amstrup, McDonald and Manly2005). For species with natural markings, individuals may be identified from photographs (taken by an observer or automated cameras). Photographic identification is commonly used for capture–recapture modelling of individually identifiable marine species (Forcada & Aguilar, Reference Forcada and Aguilar2000; Calambokidis & Barlow, Reference Calambokidis and Barlow2004). Camera traps are typically employed for cryptic or nocturnal species that are not easily observed (O'Brien et al., Reference O'Brien, Kinnaird and Wibisono2003), such as tigers Panthera tigris and leopards Panthera pardus (Karanth, Reference Karanth1995; O'Brien, Reference O'Brien, O'Connell, Nichols and Karanth2011). For photographic sampling techniques animals are identified on the first sighting and then again by re-sighting (Karanth & Nichols, Reference Karanth and Nichols1998; Smith et al., Reference Smith, Allen, Clapham, Hammond, Katona and Larsen1999), and full capture histories are developed for all identified individuals. Most recent capture–recapture models use a maximum likelihood framework, allowing allocation of model weights, and can incorporate model averaging to obtain a final estimate (Chao & Huggins, Reference Chao, Huggins, Amstrup, McDonald and Manly2005).
When individuals are not uniquely identifiable different approaches are needed to derive population estimates from camera-trap data. Under certain circumstances camera-trap encounter indices may give accurate estimates of relative abundance (O'Brien et al., Reference O'Brien, Kinnaird and Wibisono2003) although there is poor theoretical support for a general encounter rate–abundance relationship (Jennelle et al., Reference Jennelle, Runge and MacKenzie2002). Camera-trap methods require additional assumptions to relate relative abundance to density (Royle & Nichols, Reference Royle and Nichols2003). In some cases a calibrated relative abundance can be a reliable indicator of density (O'Brien et al., Reference O'Brien, Kinnaird and Wibisono2003). To eliminate the need for calibration a new method for reliably estimating densities using camera traps, without the need for uniquely marked individuals, has been proposed. The REM (Rowcliffe et al., Reference Rowcliffe, Field, Turvey and Carbone2008) estimates density by modelling the underlying process by which animals encounter camera traps (Hutchinson & Waser, Reference Hutchinson and Waser2007). By incorporating mean group size and speed of movement, encounter rates can be modelled and unbiased density estimates derived. This method has not yet been widely tested (but see Rovero & Marshall, Reference Rovero and Marshall2009; Manzo et al., Reference Manzo, Bartolommei, Rowcliffe and Cozzolino2011).
Here we examine the precision and cost of distance sampling, REM and photographic capture–recapture surveys for estimating population density and abundance of an ungulate, Grevy's zebra Equus grevyi, in a savannah habitat. The first two methods were used as part of a broader study on wildlife–livestock interactions (TGO and MFK); the latter was focused on estimating abundance of Grevy's zebra (VHZ and SRS).
Grevy's zebra is categorized as Endangered on the IUCN Red List, with < 3,000 remaining (Moehlman et al., Reference Moehlman, Rubenstein and Kebede2008). The species occurs mainly in Kenya, with a small number of isolated populations in Ethiopia. It is a territorial species that associates in unstable herds (Sundaresan et al., Reference Sundaresan, Fischhoff, Dushoff and Rubenstein2007). Reproductively active adult males defend territories (usually 6–12 km2) whereas female herds and bachelor males range more widely (Ginsberg, Reference Ginsberg1987). Individuals are easily recognized, by sight or computer identification software, from their unique stripe patterns (Hiby, Reference Hiby2010).
Currently, most monitoring of Grevy's zebra is by raw counts from the ground or air. Typically, aerial total counts are employed to estimate their populations (Parker et al., Reference Parker, Sundaresan, Chege and Brien2011) but these counts lack estimates of precision and detectability. Some researchers have begun to use mark–recapture methods to generate population estimates of Grevy's zebra (Nelson & Williams, Reference Nelson and Williams2000). Given the low overall numbers of the species and its limited range, accurate abundance estimates that incorporate precision measures are important for management and conservation.
We use Grevy's zebra as a model to explore how the three sampling methods would perform for a diversity of large mammal species in savannah habitats. Grevy's zebra may serve as a good model for three reasons. Firstly, they have home ranges similar in area to many other savannah ungulates. Secondly, they are easily observed during the day (for distance sampling and photographic transect approaches) but are also active at night (for camera trapping). Thirdly, they possess unique stripe patterns, enabling identifications of individuals.
Study area
Surveys were carried out at the 200 km2 Mpala Ranch and Conservancy, which is characterized by semi-arid Acacia bushland/grassland, in the Laikipia District of central Kenya; this area supports one of the greatest concentrations of Grevy's zebra in the country. The property, like others in the region, is unfenced and animals are free to travel across property boundaries. Mpala hosts a variety of wild ungulate species, including common zebra Equus quagga, as well as limited numbers of domestic cattle, sheep, camels and donkeys (c. 12 livestock units km−2).
Mean annual rainfall at Mpala is 594 mm in the south, where the majority of Grevy's zebra are found, but is highly variable, with droughts becoming more frequent (Franz, Reference Franz2007). The Conservancy is broadly divided into two habitats based on soil type: red sandy loam soils and clay-rich, volcanic-derived soils (‘black cotton’). Grevy's zebra are found in much lower densities across black cotton soils, so we restricted our studies to red soil habitats and the transition zones between these soil types.
Methods
Distance sampling
We conducted line transect surveys for Grevy's zebras in June 2008, January and June 2009, and January and June 2010 (Fig. 1). We used 53 transects in June 2008 totalling 98.7 km and 54 transects totalling 100.2 km thereafter. Transects were laid out systematically along roads and surveys were conducted from vehicles during 06.30–10.00 and 16.30–18.30. Two observers positioned on the roof of the vehicle at c. 3 m height recorded all groups of Grevy's zebra as part of a multi-species study. We measured observer-to-animal distances using laser rangefinders and transect bearing and angle between animal and observer with digital compasses. Perpendicular distance to the transect was calculated using the angle between the transect and the animal and radial distance from observer to animal.

Fig. 1 Mpala Ranch and Conservancy showing the areas covered for each of the three survey methods. Points and transects in the black cotton soil habitat that were not used in the analyses are not shown. The shaded rectangle on the inset shows the location of Mpala in Kenya.
It is usually inadvisable to use roads or trails for line transect surveys (Anderson, Reference Anderson2001; Buckland et al., Reference Buckland, Anderson and Burnham2001) because animals may use roads preferentially or because roads may constitute unrepresentative habitat. This is not the case for our line transect surveys because (1) animal trails traverse the habitat in a dense web so that roads do not represent special access that is restricted elsewhere, (2) vehicle use of these roads is minimal, reducing the likelihood that animals will avoid the roads, (3) animals are generally habituated to vehicles, and (4) upon examination of detection curves, we did not find any evidence of heaping (attraction) or gaps (avoidance) in the vicinity of roads.
We analysed the June 2008 and June 2010 line transect surveys (43 and 44 transects, respectively) on red soils and transition zones to estimate Grevy's zebra density using Distance v. 6.0 (Thomas et al., Reference Thomas, Buckland, Rexstad, Laake, Strindberg and Hedley2010). We used data from all five surveys to augment the estimation of the detection function and post-stratified by the June 2008 and 2010 Grevy's zebra survey vs all other surveys of Grevy's zebra combined. We analysed the data as exact perpendicular distances to zebra groups (Table 1). We evaluated half-normal, hazard, and uniform models with cosine and simple polynomial adjustments and chose the final model based on a minimum Akaike information criterion (Burnham & Anderson, Reference Burnham and Anderson2002). The June 2008 density estimate was compared to the REM estimate based on 2008 camera-trap survey data, and the June 2010 estimate was compared to the June 2010 photographic capture–recapture surveys.
Table 1 Results of the June 2008 and 2010 line transect surveys for Grevy's zebra Equus grevyi. Density estimates are presented as the number of individuals km−2 ± SE, with the coefficient of variation (CV). Abundance estimates are presented with 95% confidence intervals (CI).

1 Detection probability
2 Estimated population size
Photographic capture–recapture surveys
We used standardized photographic surveys along roads to estimate abundance. To reduce the survey area to a manageable size we divided the study area into three routes: southern, central and northern. Each section was surveyed five times on consecutive days. The south was surveyed during 8–12 June, central during 14–18 June, and north during 6–10 July 2010. A fixed route along roads and tracks, designed to minimize retracing paths on a given day, was driven during 07.00–13.00 while searching for Grevy's zebra. On each of 5 survey days roads were driven in a different order. When Grevy's zebras were encountered, as many individuals as possible were photographed using a digital SLR camera. We also recorded sex, age and reproductive status. Individuals were later identified by their stripe patterns using automated photo-identification software (Hiby, Reference Hiby2010). Individual capture histories were generated from these data and capture histories for all individuals were pooled in a single capture history file. Only three individuals were sighted in multiple survey areas and their capture histories in the two areas were combined additively into a single history (e.g. 10110 and 00001 combine to become 10111). We assume these combined histories have little effect on the overall population modelling given our large sample size. Abundance estimates from photographic surveys were generated using standard closed population models (Otis et al., Reference Otis, Burnham, White and Anderson1978). We tested for population closure using the standard Otis test (1978). Population size estimates were obtained with MARK v. 6.1 (White & Burnham, Reference White and Burnham1999). We estimated the area surveyed using a minimum convex polygon around the combined survey routes, subtracting the area of an adjacent property that cuts into the Mpala Conservancy and that was not surveyed. An estimated area of 156 km2 was sampled and used to calculate densities.
Random encounter model
The REM is a method for estimating densities with photographs from camera traps, without the need for uniquely marked animals (Rowcliffe et al., Reference Rowcliffe, Field, Turvey and Carbone2008). The REM estimates density by modelling the underlying process by which animals encounter camera traps (Hutchinson & Waser, Reference Hutchinson and Waser2007). By incorporating average group size (g) and average speed of movement (v), encounter rates can be modelled and unbiased density estimates (D) can be derived (Equation 1). Camera-related parameters include trapping rate (y/t) and trap detection zone, for which we measure detection distance (r) and angle (θ).
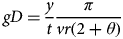 $$gD = \displaystyle{y \over t}\displaystyle{\pi \over {vr(2 + \theta )}}$$
$$gD = \displaystyle{y \over t}\displaystyle{\pi \over {vr(2 + \theta )}}$$Camera traps were set between 8 January and 12 April 2008. We divided the ranch into 2 km2 blocks, located the centroid of each block (n = 97 blocks) using ArcView v. 3.2 (ESRI, Redlands, USA) and designated these points as potential trap sites (Fig. 1). Actual trap points were located at an ‘optimal’ location (the UTM coordinates of which were determined with a global positioning system, GPS), within a 50 m radius of the centre point. Optimal camera-trap placement was subjective and typically included a road or active game trail. Two Deercam film cameras (Non-Typical Inc., Park Falls, USA) were mounted on posts at each point to photograph both sides of passing animals. Sampling took place sequentially in three blocks of 25 points, and one block of 22 points for a total of 97 points. Ten of these points were in black cotton soils and were excluded from analyses. Each point was active for 21–23 days and cameras were checked on days 5, 10 and 15, with films and batteries being changed as needed. Each photograph was stamped with a time and date that facilitated assigning photographic events to sampling intervals. Cameras were set with a 30 second delay between photographs to avoid repeated triggering by the same animals lingering in front of the camera.
We then used REM to derive density, which was appropriate in this case because we did not have a sufficient number of photographs suitable for identification of individuals. To calculate encounter rates between Grevy's zebras and camera traps we combined sequential photographs into a single encounter when it was clear that a single group was moving through the field of view (usually a series in < 5 minute span, maximum 19 minutes). Grevy's zebra speed and clustering patterns were estimated independently. Speed (v) was derived from hourly GPS collar data from seven adult zebras collared on the Mpala Conservancy, collected for 1–9 months beginning in June 2007 (D.I. Rubenstein & S.R. Sundaresan, unpubl. data). Mean group size (g) was calculated from driving censuses conducted on Mpala during 2007–2008. Camera-related parameters r (12 m) and θ (0.175 radians) were taken from Rowcliffe et al. (Reference Rowcliffe, Field, Turvey and Carbone2008) because we used the same model and placement of Deercam camera traps. We estimated variance in encounter rates (y/t) by bootstrapping, resampling camera locations with replacement 10,000 times, as recommended by Rowcliffe et al. (Reference Rowcliffe, Field, Turvey and Carbone2008). Overall variance of density estimates was computed using the delta method (Seber, Reference Seber1982; J.M. Rowcliffe, pers. comm.).
Cost comparison
We evaluated the cost effectiveness and efficiency of each method using start-up costs (equipment purchases) and the cost of transportation and human resources.
Results
Distance sampling
We observed 51 Grevy's zebras in nine groups during the June 2008 line transect surveys on red and transition soils. In June 2010 we detected 33 Grevy's zebras in three groups. We used data from all five surveys from June 2008 to June 2010 (n = 40 groups) to improve the detection function. We then post-stratified by survey period to estimate densities for each period. We only report results for 2008 and 2010 surveys for comparison with the REM and photographic capture–recapture methods, respectively. The best-fitting model was a hazard rate model with two cosine terms and the arithmetic mean for group size. We estimated densities of 1.82 ± SE 1.12 individuals km−2 in 2008 and 1.21 ± SE 0.768 individuals km−2 in 2010, and thus population estimates of 313 and 207 Grevy's zebras in the study area in 2008 and 2010, respectively (Table 1).
Photographic capture–recapture surveys
During 15 days of photographic capture–recapture surveys we were able to photograph the majority of Grevy's zebras sighted. In 36 of 40 sightings (90%) we photographed at least one Grevy's zebra in each group. The average proportion of individuals in a group photographed and successfully identified within a sighting was high (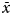 $\bar x$ = 98.5%). Foals were excluded from our analyses because their movements are not independent of their mothers. In total 103 adult individuals were identified.
$\bar x$ = 98.5%). Foals were excluded from our analyses because their movements are not independent of their mothers. In total 103 adult individuals were identified.
Otis' test (1978) supported the closure assumption (Z = 2.75, P = 0.997). For both the standard closed capture and the Huggins linear logit estimates a time-based/heterogeneity model was the best fit, yielding similar results (M th,Otis: 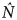 $\hat N$ = 145 ± SE 15.1; M th,Huggins:
$\hat N$ = 145 ± SE 15.1; M th,Huggins: 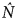 $\hat N$ = 147 ± SE 15.5). Most of the models produced comparable estimates with similar confidence intervals (Table 2) suggesting that the set of closed models is a good fit for our photographic sighting data. Density estimates were based on a sampling area of 156 km2 (
$\hat N$ = 147 ± SE 15.5). Most of the models produced comparable estimates with similar confidence intervals (Table 2) suggesting that the set of closed models is a good fit for our photographic sighting data. Density estimates were based on a sampling area of 156 km2 (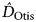 $\hat D_{{\rm Otis}} $ = 0.93 ± SE 0.10 and
$\hat D_{{\rm Otis}} $ = 0.93 ± SE 0.10 and  $\hat D_{{\rm Huggins}} $ = 0.94 ± SE 0.10). These results are lower than the line transect estimates for 2010 but had overlapping confidence intervals, and are not significantly different (Fig. 2).
$\hat D_{{\rm Huggins}} $ = 0.94 ± SE 0.10). These results are lower than the line transect estimates for 2010 but had overlapping confidence intervals, and are not significantly different (Fig. 2).

Fig. 2 Comparison of the 2008 and 2010 density estimates for Grevy's zebra Equus grevyi for the three sampling techniques: line transects, random encounter model (REM) and photographic capture–recapture (Photo C–R). Figures are presented as individuals per km with SE bars.
Table 2 Estimated abundance and capture probabilities of adult Grevy's zebras from the June and July 2010 photographic capture–recapture surveys under different closed population models generated with MARK. Time varying models with heterogeneity, shown in bold, were chosen based on Akaike information criterion (AIC) values. All models use a logit link function. We ran a basic behavioural model but no combinations thereof.

1 Estimated population size ± SE with lower and upper 95% confidence limits
2 Detection probability, averaged in the case of multiple detection probabilities
3 Only nine were estimated, two were singular
Random encounter model
From camera-trapping data we encountered 40 separate Grevy's zebra groups at 87 camera points over an average of 19.4 trap days, yielding an encounter rate of 0.0236 encounters per day (Table 3). Bootstrapping the encounter rate gave a standard error of 0.0076. The velocity calculated from hourly zebra GPS locations using collar data was 0.34 km h−1 (n = 12,141 hourly locations, SE = 0.004) or 8.16 km day−1. Group size, as determined from independent counts during road surveys in Mpala, was 4.58 individuals per group (n = 224, SE = 0.316). Camera-related parameters (Rowcliffe et al., Reference Rowcliffe, Field, Turvey and Carbone2008) were assumed to have no variance. The resulting density estimate was 0.346 groups km−2 (SE = 0.190) or 1.58 Grevy's zebra km−2 (SE = 0.42). The REM density estimate is slightly lower than the line transect estimate for 2008 but has overlapping confidence intervals and is not significantly different (Fig. 2).
Table 3 Summary of the variables required to calculate density of Grevy's zebra from camera-trapping rates using Rowcliffe's random encounter model (Rowcliffe et al., Reference Rowcliffe, Field, Turvey and Carbone2008). The mean value for each variable is presented ± SE, with the coefficient of variation (CV).

Cost comparison
Because all three methods produce comparable results, choosing the most cost-efficient method will depend on the implementation cost relative to the cost of improving precision. Basic equipment for line transect surveys includes two sets of binoculars, laser rangefinders, and digital compasses (USD 1,750; Table 4). Photographic capture–recapture simply requires a digital camera and lens (USD 700). Camera-trap projects have the highest initial investment in camera traps (30 traps at USD 200 each), batteries and memory cards (USD 6,860 total). For density estimates using camera traps we also required GPS collars (USD 3,000 each) to collect movement data and additional field costs to collect group size data. Person days in the field were greatest for photographic capture–recapture and less for camera trapping and line transects, which were similar. Data processing was most intensive for photographic capture–recapture and least intensive for distance sampling (Table 4). The running cost is similar for line transect surveys (USD 1,160) and camera-trap surveys (USD 712 using 30 camera units) and lower compared to photographic capture–recapture (USD 2,045). Most of the difference in running cost is because of the time required for post-processing, which may be substantial for camera traps and photographic capture–recapture. Both methods require processing photographs of animals prior to data analysis and the time taken for this is related to the number of photographs taken.
Table 4 Resource comparison for the three sampling methods. The cost of vehicles and GPS units were common to all projects and are not presented here. Note that two vehicles were used in distance sampling whereas single vehicles were used for the other techniques. All methods covered a survey area of c. 170 km2.

Discussion
Our results show that line transect surveys, photographic capture–recapture surveys and the REM methods provide congruent estimates of the density and abundance of Grevy's zebra. Any one of these methods is preferable to the common practice of attempting to conduct complete censuses of populations. If we had used only sighting data (i.e. untransformed counts from transects or counts of individuals identified from photographs) from line transect and photographic capture–recapture surveys we would have captured only c. 16 and 71% of the estimated Mpala population, respectively. Our results indicate that census data may lead to spurious conclusions about Grevy's zebra populations, and that any sampling methods that incorporate measures of detection probability and precision are preferable (Williams et al., Reference Williams, Nichols and Conroy2002).
For short-term surveys over small areas we recommend using photographic capture–recapture techniques to yield the most precise abundance estimates of easily observable, individually identifiable species. Distance sampling, although somewhat imprecise in this particular case, is an affordable method for assessing densities of species that are directly observable, and may be desirable when monitoring multiple species simultaneously or comparing across sites. Our study suggests that REM may be an accurate and appropriate method for difficult-to-observe and unmarked species being monitored over time.
In general, the photographic capture–recapture method is highly precise when abundance is used as the monitoring metric in a relatively small area and when recapture rates are high. This method has been used for closed population estimates of Grevy's zebra in northern Kenya (Nelson, Reference Nelson2003). In a study of Arabian oryx Oryx leucoryx, an ungulate of similar size, detection cues and densities, mark–resighting was more precise than distance sampling (Seddon et al., Reference Seddon, Ismail, Shobrak, Ostrowski and Magin2003). Photographic capture–recapture also proved to be the most precise (CV 10.4%) of the three survey techniques in our study but is the most expensive method because of the substantial costs of data collection and processing. One advantage of this technique is that field methods are simple and start-up costs are low.
Although the field component of photographic surveys is rapid and cost-effective for short-term surveys, the identification of animals is time-consuming and requires either comprehensive knowledge of individual markings or the use of an impartial, automated identification system. If appropriate software is not available, identification by eye may be expensive in terms of both time and cost, and may be unreliable. This can be controlled through careful training of technicians involved in data processing but is much easier when automated identification software is used (Hiby et al., Reference Hiby, Lovell, Patil, Kumar, Gopalaswamy and Karanth2009).
Photographic capture–recapture requires that individuals are photographed at fairly close range, which may prove difficult in bush habitat or in areas where animals have become shy of humans. In this case, bias can arise because some individuals in the population are uncatchable. We do not believe there is any such bias in our population of Grevy's zebra as we photographed the majority of animals encountered. More generally, it is possible that using heterogeneity models (M h) may be able to account for unequal catchability in other species.
Using the photographic capture–recapture method to monitor density, as opposed to abundance, may be problematic because we do not have an objective method for estimating the sampling area. In our study it is impossible to determine if the small difference between the 2010 density estimates from line transects (1.2 km−2) vs the capture–recapture method (0.93 km−2) is best explained by true variation or by our ad hoc estimation of survey area for our photographic surveys. The photographic method does not generate movement data that can be used in a number of ad hoc density estimation procedures (Wilson & Anderson, Reference Wilson and Anderson1985; Jett & Nichols, Reference Jett and Nichols1987; Parmenter et al., Reference Parmenter, Yates, Anderson, Burnham, Dunnum and Franklin2003). We estimated the area of coverage based on a minimum convex polygon but it is possible that we overestimated the coverage, resulting in an underestimation of density. Given the uncertainty in estimating the area sampled we recommend that the photographic method be used with abundance as the state variable of interest.
The utility of photographic capture–recapture surveys can be extended by targeted, long-term monitoring, which can provide estimates of population vital rates. This method would also facilitate estimation of survival when using open population models and Pollock's robust design (Pollock, Reference Pollock1982) applied to data collected over several seasons (Karanth et al., Reference Karanth, Nichols, Kumar and Hines2006).
Distance-based sampling surveys yielded the least precise (CV 61–64%) density estimates, even with the use of ancillary data to augment the detection function. This is a common problem when distance-based sampling methods are applied to rare species that occur in widely dispersed groups (Otto & Pollock, Reference Otto and Pollock1990). Low encounter rates, with many transects having zero encounters, can dominate the variance in density; in our study > 70% of overall variance was because of variance in encounter rate. Large variation in group sizes (1–16 in this study) can also inflate the variance of density. In this study variation in group size has less of an effect than the variance in encounter rate. Low encounter rates are difficult to improve through changes in sampling design as using more transects does not guarantee an increase in the encounter rate.
Distance sampling is based on the idealized scenario of animals being distributed in space according to a stochastic process with rate parameter D (density). Transects are placed at random or systematically to ensure that objects in the survey strip are uniformly distributed in relation to distance from the transect. Although we were restricted to using roads we believe that our methods meet the qualification of a systematic sample with a randomized starting point and that the open landscape allowed us to sample almost all potential habitats. In many areas occupied by Grevy's zebra the development of systematic survey designs may be feasible. Distance sampling rests on four basic assumptions: (1) animals on the transect are always detected, (2) animal locations are always measured to the point where the animal was first detected, (3) distances to the animals and angles between the animal and the transect are measured exactly, and (4) groups are counted accurately (Buckland et al., Reference Buckland, Anderson and Burnham2001). Grevy's zebras are large conspicuous animals, so the first assumption is met. Accurate measurements are possible using laser rangefinders and digital compasses, and this helps to minimize heaping of observations on the transect line and also improves estimation of perpendicular distances. Recording group sizes accurately can be problematic in some bush habitats and care must be exercised to ensure that group counts are as complete as possible.
However, there are several benefits of using density estimates from distance sampling as a monitoring metric. Firstly, start-up costs are low and it is relatively inexpensive to process data because recognition of individuals is not required. Secondly, line transect surveys may be used to estimate density for several large ungulates simultaneously. In this case, the lack of precision for rare species may be outweighed if there is a need to derive estimates for more common species. Thirdly, density estimates are comparable between sites whereas abundance estimates cannot be compared directly between sites. We recommend the use of distance-based density estimates as a state variable when between-site comparisons are important and when multi-species monitoring methods are needed.
Our density estimate from REM was consistent with the estimates from distance-based sampling and was more precise (CV 26.3%). In contrast, when testing REM for forest ungulates Rovero & Marshall (Reference Rovero and Marshall2009) found that it yielded higher estimates than line transect counts. Although the REM estimate was more precise than for distance sampling it may be untenable to generate accurate speed-of-movement data in wide-ranging species; GPS collar data for this study were obtained from a separate study. Additionally, group size of Grevy's zebra is highly variable because of the fluid nature of their social structure, and required an assessment from previous surveys. The high start-up cost of REM may make it impractical for short-term studies or projects with limited funding. Camera malfunction and theft may also pose significant obstacles. A large number of cameras are required for ungulates occurring at low to medium densities but fewer cameras or trapping days may be feasible for common species. In this study, however, cameras were not placed specifically to capture Grevy's zebra, as this was part of a multi-species study. Variation in trapping rate and precision could be reduced with a more targeted trapping scheme. This method is best for shy animals, provided that unbiased, independent estimates of speed and group size can be generated.
Cost may be a limiting factor in the choice of a field survey technique. The REM has the highest equipment start-up costs but because it requires little field time it is cheapest in the long term. The photographic capture–recapture method had the lowest start-up costs, but high operation costs because of the amount of time in the field and the time to process photographs. The line transect method is intermediate for both start-up costs and field time, and data analysis is quickest.
In conclusion, all three methods are suitable for estimating Grevy's zebra densities and are preferable to complete census counts. Based on our study we can make some wider recommendations for the monitoring of other similar ungulate species. For small or enclosed properties, photographic capture–recapture surveys may be the best choice for estimating wildlife abundances provided the substantial operating costs are affordable. High-precision methods may be required when monitoring is part of a management action that is likely to result in subtle abundance changes over time. For multi-species programmes distance sampling or REM are preferred as they may provide sufficient precision and data can easily be gathered for several species simultaneously. Distance sampling lacks the precision of the other two methods especially when encounter rates are low but it is appropriate for easily observable or common species, for animals without individual markings or for long-term studies. The REM is an efficient and precise method of estimating densities but has high initial costs. We believe REM, although relatively new, holds promise for estimating density of many wildlife species.
Acknowledgements
We thank J. Marcus Rowcliffe for discussion and advice on estimating variance associated with REM. For assistance in the field we thank Francis Lomojo, Hussein Mohamed, Frankline Otiende and Allison Williams. This work was funded by Panthera, Wildlife Conservation Society, Leslie Scott, Denver Zoological Foundation and Princeton University. For supporting the authors during this study we thank Princeton University (VHZ), Denver Zoological Foundation (SRS), Wildlife Conservation Society (TGO) and the Mpala Wildlife Foundation (MFK).
Biographical sketches
Victoria H. Zero's research interests focus on the ecology and conservation of terrestrial vertebrates. Siva R. Sundaresan conducts research on large mammalian ungulates and carnivores. He directs Denver Zoological Foundation's conservation programme in Kenya, which includes research on Grevy's zebra. Timothy G. O'Brien has conducted research on primates, birds and terrestrial mammals, and is especially interested in the development of accurate and precise monitoring metrics. He is currently a senior scientist and statistician at the Wildlife Conservation Society. Margaret F. Kinnaird has conducted research on a wide variety of topics, including human–elephant conflict, parrot trade, forest dynamics, non-timber forest product use, and hornbill and primate conservation. She currently serves as the Executive Director of the Mpala Research Centre where her research interests include how different livestock management regimes influence wildlife abundance and diversity.






