


Introduction
Management of threatened and endangered populations of wildlife increasingly relies upon active intervention such as predator control, habitat manipulation, and ex situ breeding or care (Cohen et al. Reference Cohen, Karpanty, Catlin, Fraser and Fischer2008, Barber et al. Reference Barber, Nowak, Tulk and Thomas2010, Escobar et al. Reference Escobar, Besier and Hayes2010). Often, however, the effect of these conservation actions is not directly quantified in terms of demographic changes to the individuals or populations of interest (Calvert and Taylor Reference Calvert and Taylor2011). Managers may therefore develop techniques that accomplish the proximate objective (e.g. successful ex situ breeding) but the ultimate goal of population enhancement or stabilisation may not be achieved. Furthermore, side-effects such as behavioural modifications, which subsequently may affect demography or site fidelity, are often not assessed (Beaulieu et al. Reference Beaulieu, Austin and Leonard2014).
One tool that has received considerable attention for the management of declining or threatened avian populations is headstarting, or the artificial incubation of eggs and subsequent placement of newly hatched chicks in original or foster nests. Also referred to as ‘hatch-and-switch’, headstarting has been used in various forms for cranes, shorebirds, passerines and other endangered birds (Powell et al. Reference Powell, Cuthbert, Wemmer, Doolittle and Feirer1997, Robertson et al. Reference Robertson, Colbourne, Nelson and Westbrooke2005, King et al. Reference King, Espenshade, Kirkpatrick-Wahl, Lapinski, Malekan and Rickett2013, Lobo and Marini Reference Lobo and Marini2013) and has also been employed for the conservation of iguanas (Perez-Buitrago et al. Reference Perez-Buitrago, Garcia, Sabat, Delgado, Alvarez, McMillan and Funk2008, Escobar et al. Reference Escobar, Besier and Hayes2010). Headstarting appears to be most effective as a tool to avoid likely nest failures when productivity is lost during incubation and when parents can be enticed to remain at nests through the placement of artificial eggs (Elliott et al. Reference Elliott, Merton and Jansen2001). A typical headstart programme would include the following steps: (1) collection of eggs from the nest during incubation, (2) replacement of collected eggs with artificial eggs that are secured to the nest site with an anchor and painted to resemble natural eggs, (3) incubation of collected eggs in an incubator in a controlled setting, and (4) release of ex situ incubated chicks into the nest immediately after hatch. If successful, nest survival and hatching success rates would improve over baseline levels and subsequently lead to an enhancement in annual productivity.
We assessed the feasibility of implementing a headstarting program for the American Oystercatcher Haematopus palliatus, a species of high conservation concern in the eastern USA (Brown et al. Reference Brown, Hickey, Harrington and Gill2001, Schulte et al. Reference Schulte, Brown and Reynolds2010). The species nests on beaches, barrier islands, salt marshes, dredge spoil islands, and waterways along the USA Atlantic and Gulf coasts (American Oystercatcher Working Group Reference Gill and Poole2012). The population size is estimated at c.11,000 individuals with c.2,500 breeding adults (American Oystercatcher Working Group Reference Gill and Poole2012). In the south-eastern USA, productivity is often low and highly variable both within and between sites and years (McGowan Reference McGowan2004, McGowan et al. Reference McGowan, Simons, Golder and Cordes2005, Jodice et al. Reference Jodice, Thibault, Collins, Spinks and Sanders2014). Nest loss is common although chick survival has been studied less often (Sabine et al. Reference Sabine, Schweitzer and Meyers2006, Wilke et al. Reference Wilke, Watts, Truitt and Boettcher2005, Denmon et al. Reference Denmon, Watts and Smith2013, Jodice et al. Reference Jodice, Thibault, Collins, Spinks and Sanders2014). Given the evidence of low and/or variable annual productivity, methods to enhance reproductive success of oystercatchers in this region are currently being considered, including headstarting of eggs to avoid nest loss.
We tested an ex situ hatching program for oystercatchers as a means to reduce nest failure and enhance annual productivity. We used a control-impact approach to assign nests as either control or headstart, measured their daily survival rate and productivity, and determined likely causes of failure. We also measured the success of hatching in the incubator, the success of parents accepting headstarted chicks, and attendance patterns and behaviours of parents attending control and headstart nests. These data provide an initial assessment of the effectiveness of a headstarting program for producing chicks and identify potential factors that contribute to reproductive failure. If productivity is lost primarily during the incubation stage due to factors such as flooding or predation, then headstarting nests may enhance productivity by improving nest success (Kuehler et al. Reference Kuehler, Lieberman, Oesterle, Powers, Kuhn, Kuhn, Nelson, Snetsinger, Herrmann, Harrity, Tweed, Fancy, Woodworth and Telfer2001, Lobo and Marini Reference Lobo and Marini2013). In contrast, if productivity is primarily or additionally lost during chick-rearing, then headstarting may not provide a means to enhance reproductive success but instead may simply increase hatching success without benefitting annual productivity.
Methods
Study area
Research was conducted in and adjacent to Cape Romain National Wildlife Refuge (CRNWR), South Carolina, USA (Figure 1; hereafter Cape Romain Region, CRR) which is comprised of barrier islands, shallow bays, tidal creeks, salt marsh (dominated by Spartina alterniflora), mudflats and eastern oyster (Crassostrea virginica) reefs. South Carolina supports c.400 pairs of American Oystercatchers and c.230 pairs nest within CRR (Sanders et al. Reference Sanders, Murphy, Spinks and Coker2008). Oystercatchers in the CRR nest primarily on elevated mounds of oyster and clam Mercenaria sp. shells formed by wind and wave energy along sections of bays, estuarine islands and waterways (Jodice et al. Reference Jodice, Thibault, Collins, Spinks and Sanders2014). We monitored oystercatcher nests on shell mounds in two study areas during the 2010 and 2011 breeding seasons, along the Atlantic Intracoastal Waterway (hereafter, AIWW), and in the south-western section of Bulls Bay (hereafter, SWBB; Figure 1). The AIWW is a navigable waterway that has seasonal migrations of large boats, creating substantial wakes that wash over shell mounds. In contrast, SWBB is shallow and surrounded by Spartina salt marsh.
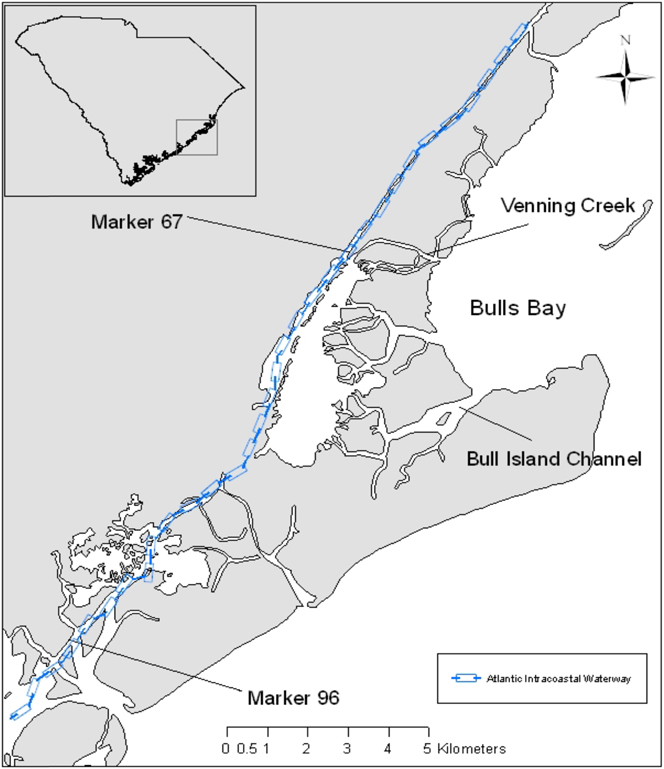
Figure 1. Study area within the Cape Romain Region, South Carolina. Study nests occurred along the Atlantic Intracoastal Waterway between markers 67 and 96, and in Bulls Bay between Venning Creek and the Bull Island Channel.
We defined four steps in our headstart programme, each with one or more measurable outcomes. For measurable outcomes where proportions were assessed for success, we set ≥ 85% as the success threshold and ≤ 15% as the failure threshold. Intermediate values were defined as partially successful. For outcomes with a single measure for each of the four site-years (i.e. DSR of nests, DSR of broods, and productivity), we defined a significant statistical test (P ≤ 0.05) differentiating headstart from control nests as fully successful, a marginally significant statistical test (0.05 < P ≤ 0.10) that suggested a slight differentiation between headstart and control nests as partially successful, and a nonsignificant statistical test (P > 0.10) that failed to distinguish between headstart and control nests as unsuccessful.
Step 1: Nest detection, egg collection, and artificial incubation
The first step was to locate nests, assign them to experimental groups, collect and transport eggs for headstarting, and artificially incubate eggs. Oystercatchers are territorial during the breeding season and are very conspicuous on their territories when protecting a nest, allowing a high detection rate for nests. Nest searches of shell rakes along the AIWW from marker 67 to 97 (c.12.6 km) and in SWBB from Venning Creek to Bulls Island Creek occurred about every three days from March to July, 2010 and 2011. We classified nests in the order they were found within each study area, alternating between headstart and control assignments. Any re-nest attempts that occurred at the same nest site retained its original classification. For each detected nest we recorded the location (± 3 m) using a handheld GPS, the number of eggs present, and the band combinations of any adults observed on the nesting territory (banding was routinely conducted by agency biologists as part of a state-wide monitoring programme). At time of detection we also installed washover cups to assess any occurrence of over-wash or localised flooding (Brooks et al. Reference Brooks, Sanders, Gerard and Jodice2013). Plastic cups (c.350 ml) were positioned adjacent to nests, secured to the ground by affixing nails to the base, had holes set along the upper circumference to allow inflow of water from horizontal movement (e.g. flooding), and lids which restricted water flow from vertical movement (e.g. rain).
For headstart nests we collected all but one egg from the clutch (clutches here are typically 1-3 eggs): we left one egg to enhance our ability to interpret predation or flooding events at the nest. Each collected egg was labelled with a non-toxic pen to identify its nest origin and measured for length (L), breadth (B) and weight. Egg volume (cm3) was calculated as 0.51 * LB2 (Hoyt Reference Hoyt1979). Wooden artificial eggs were placed in each headstart nest to encourage parents to continue to incubate. Artificial eggs were painted to resemble oystercatcher eggs and attached with string to nails (2010) or a rebar anchor (2010 and 2011). Eggs were transported to a facility located in CRNWR (9 km maximum distance from any nest) in an unheated padded bin and placed in a cabinet-style incubator (Brinsea Ova-Easy 190). In 2010, the initial temperature and humidity were set at 37.6°C and 50% following recommendations for domestic poultry eggs. In 2011, we lowered the incubator temperature to 37.4°C and increased humidity to 65% following settings suggested for artificial incubation of shorebird eggs (Powell et al. Reference Powell, Cuthbert, Wemmer, Doolittle and Feirer1997).
Measurable outcomes from step one focused on headstart nests only and included (1.1) proportion of eggs collected and transported to the incubation chamber without damage, and (1.2) hatching rate of eggs in the incubator.
Step 2: Nest survival and incubation behaviour
Step two was to monitor, measure daily survival rate (DSR), and measure parental behaviour during the incubation phase at both control and headstart nests. We visually checked the status of each control and headstart nest on average (± 1 SD) every 3.1 ± 1.1 and 3.0 ± 1.4 days, respectively, until the nest failed, was abandoned, hatched (control nests), or until chicks were returned to the nest (headstart nests). During each nest check, we recorded the date, time, number of eggs, tide phase, and number of adults present. Nest status was recorded as surviving (≥ 1 viable egg remains for control nests, parents actively attending headstart nests) or failure. If a nest failed we classified the cause as predation (signs of predation at the nest coincident with nest abandonment, e.g. teeth marks on artificial eggs, artificial eggs removed from scrape, egg shell fragments observed near nest, scat observed near nest), over-wash (over-wash cup contained salt water, over-wash cup dislodged from shell rake, recently deposited rack observed near/on nest or fake eggs buried under shells; any or all coincident with parental abandonment), abandoned (adults not observed incubating for at least three visits, or new scrape discovered later in the incubation cycle but no signs of predation or over-wash observed), undetermined, or other (one event, one adult of breeding pair appeared to be preyed upon by a raptor).
Parental attendance and behaviour were measured at headstart and control nests during incubation ± 2 hours from peak low tide when adults typically forage (Thibault et al. Reference Thibault, Sanders and Jodice2010). Surveys (53–90 mins in duration) were conducted from land or boat from a distance of at least 150 m to minimise potential impacts on behaviour (Thibault et al. Reference Thibault, Sanders and Jodice2010). We conducted continuous behavioural observations and recorded attendance and behaviour of each adult and the duration of each behaviour while present on the territory (i.e. times were noted when parents departed or arrived). We also recorded the time of day, and clutch size for each survey. Eighteen behaviours were initially identified and activities were then condensed into categories following Sabine et al. (Reference Sabine, Schweitzer and Meyers2006): reproductive (copulating, incubating eggs, maintaining nest, and brooding); self-maintenance (preening, bathing, stretching); locomotion (flying and walking); forage (using bill to open prey or probe substrate for prey and drink); rest (standing or sitting with head turned back and bill tucked under wing); vigilance (standing with no bill tuck); alarm (piping display, head bobbing, chasing, being chased, or other agnostic behaviour); and unknown.
Measurable outcomes from step two included (2.1) proportion of headstart nests for which artificial eggs remained secured during the entire incubation period, (2.2) proportion of measured behaviours for which there was no difference in time allocation between control and headstart nests during the incubation phase, and (2.3) daily survival rate of control compared to headstart nests.
Step 3: Placement of headstart chicks in nests
The third step was to place chicks hatched from the incubator into nests. Eggs within the incubator were monitored regularly for signs of hatching and returned to the original nest within 24 hours of hatching. If artificial eggs were lost, washed away, or buried, or if adults discontinued incubation before the collected eggs hatched, chicks were fostered into another headstart nest with a similar estimated hatch date (Reed et al. Reference Reed, Nilsson and Murray1993, Lobo and Marini Reference Lobo and Marini2013). After chicks were placed in the nest scrape, we observed nests to verify that adults accepted and brooded the returned chicks. The measurable outcome from step three was (3.1) proportion of headstart chicks accepted by parents (either original or foster parents).
Step 4: Brood-rearing behaviour, chick survival, annual productivity
Step four included measuring parental behaviour during chick-rearing, DSR of broods, and annual productivity. Parental attendance and behaviour were measured during chick-rearing following the same methods described above for incubation, except we recorded age and number of chicks during surveys instead of clutch size, and we also included provisioning of chicks as a behaviour within the reproductive category.
To measure DSR of broods we banded and radio-tagged chicks. All chicks (headstart and control) were banded with unique colour combinations at c.10-15 days post-hatch. We also attached 1.3 g VHF transmitters (Advanced Telemetry Systems, Isanti, MN) to the scapular region of newly hatched headstart chicks with a generic instant epoxy before they were returned to the nest. Transmitters averaged 4% of body mass for 0–2 day old chicks. Headstart chicks were then located on average every 3.1 ± 1.2 days. During each visit, chicks were examined for any evidence of physical damage from the transmitters. Glue was added to the transmitter on visits when the transmitter appeared loose. When a transmitter fell off, we attempted to relocate the chick and reattach the transmitter. If a chick died, we attempted to locate its remains and searched the area to determine the cause of death. Control chicks were not marked with radio transmitters due to funding limitations but monitored by searching shell rakes on average every 3.3 ± 1.9 days. If a chick was not found on the shell rake during two consecutive visits, pairs would be monitored at a distance until either the chicks were observed or adult behaviour indicated that chicks were no longer present (e.g. no longer actively defending a territory, vacating a shell rake and not returning when disturbed). All chicks were monitored until they were considered “fledged” at 35 days or when observed in flight.
Measurable outcomes from step four for which success or failure could be assessed included (4.1) proportion of measured behaviours for which there was no difference in time allocation between control and headstart nests during the chick-rearing phase, (4.2) DSR of control compared to headstart broods, and (4.3) annual productivity (chicks per pair) at control compared to headstart nests.
Statistical analysis
For collected eggs, we report hatching success as the percentage of collected eggs that successfully hatched in the incubator. For control nests, a nest was defined as successful if at least one egg hatched. For headstart nests which contained artificial eggs we defined nesting success as parents continuing to attend and incubate until the hatch date so that at least one chick could be returned to the nest. For both headstart and control nests we considered a brood successful if ≥ 1 chick fledged (Jodice et al. Reference Jodice, Thibault, Collins, Spinks and Sanders2014).
A mixed model (SAS, PROC GLIMMIX) was used to analyse the hatching success of headstart eggs in the incubator. Site (AIWW or SWBB), collection date, egg weight, and egg volume were included as fixed effects and year (2010 or 2011) was included as a random effect. An ANOVA was used to detect differences between the weight and volume of headstart eggs between sites and years.
Nest and brood success were analysed as daily survival rates (DSR) using logistic-exposure models (SAS, PROC GENMOD; Shaffer Reference Shaffer2004). The following nest-, local-, and time-specific explanatory variables were considered: nest type (control or headstart), year (2010 or 2011), site (AICW or SWBB), nest age, nest age2, date (represented as the day in the nest season with 1 April as day 1), date2, and tide height (maximum during interval between visits). To be consistent we followed the same approach for brood survival models although parent type (original, foster or mix parents with a combination of both real and foster chicks) was also included as an independent variable in the survival model for headstart broods. We calculated the probability of a nest surviving from egg-laying to hatch (control) or from egg-laying to return of a headstart chick (headstart) as the DSR from the final model raised to an exponent equal to the typical number of days in the incubation stage, 27. Similarly we calculated the probability of a chick surviving from hatch to fledge as the DSR from the final model raised to an exponent equal to the typical number of days to fledge, 35. We used a paired t-test to determine if there was a significant difference in productivity between control and headstart nests, with the pairs set as the productivity measures for control compared to headstart nests within each site-year.
We used general linear regression models (SAS, PROC GLM) with a manual backward-elimination process to examine the relationship between nest type (control or headstart) and adult behaviours. The percentage of time adults were present at their territory during the low-tide foraging period and the percentage of time during which parents engaged in each of the aforementioned behaviours while attending were used as response variables in these regressions. Analyses were conducted separately for incubation and chick-rearing. Percentages were transformed using the arcsine square-root transformation to standardise the variance for analyses, although untransformed values are presented throughout for ease of interpretation. During chick-rearing, analyses were limited to comparisons of attendance and behaviour for first nest attempts along the AIWW in headstart and control nests due to small and unbalanced samples among site, nest type, brood size, and nesting attempt number. We pooled data among brood sizes because there was no significant relationship between attendance and brood size in headstart or control nests along the AIWW (F1,5 ≤ 0.09, P ≥ 0.78 for each). We also assessed the relationship between nest success and attendance and each behavioural category separately for headstart and control nests using general linear regression models (SAS, PROC GLM) with nest success as the independent variable. For all analyses we report actual P-values. Means are presented ± 1 standard deviation and coefficient estimates are presented ± 1 standard error unless otherwise stated.
Results
Step 1: Nest collection, egg detection, and artificial incubation
We monitored 94 pairs of oystercatchers and 159 nest attempts during the two study years (Table 1). Of these pairs, 46 were classified as control pairs and these initiated 90 nest attempts (initial nesting effort plus replacement clutches upon failure) while 48 were classified as headstart pairs and these initiated 69 nest attempts (Table 1; Figure 2). We collected 91 eggs from 71 clutches. No eggs were lost or broken during collection or transport. Weight (44.2 ± 3.8 g) and volume (43.2 ± 3.6 cm3) of eggs collected for headstarting did not differ between years or among sites (F1,20 ≤ 2.19, P ≥ 0.15 for each). Hatching success in the incubator was affected by year (F 1,17 = 6.8, P = 0.02) but not by site, collection date, egg mass, or egg volume (F1,68 ≤ 2.45, P ≥ 0.12 for each). The odds of an egg hatching in the incubator in 2011 were 5.7 times greater than the odds of an egg hatching in the incubator in 2010. Hatching success in the incubator was 62% for all eggs and 70% for fertile eggs in 2010, and 84% for all eggs and 94% for fertile eggs in 2011. Approximately 45% of eggs that hatched in 2010 and 3% of eggs that hatched in 2011 in the incubator resulted in chicks with deformities.
Table 1. Reproductive effort of control and headstart nests of American Oystercatchers in the Cape Romain Region, South Carolina, 2010–2011. AIWW = Atlantic Intracoastal Waterway, SWBB = Southwest Bulls Bay.


Figure 2. Distribution of headstart and control nests of American Oystercatchers along (a) the Atlantic Intracoastal Waterway and (b) Southwest Bulls Bay, Cape Romain Region, South Carolina, April–July, 2010 and 2011. Only original nests (i.e. not re-nests, which occurred in the same locations) are shown to reduce clutter.
Step 2: Nest survival and incubation behaviour
Daily survival rate of control nests ranged from 0.894 to 0.954. The probability of a control nest surviving to hatching ranged from 7% to 30% among both sites and years (Table 2). Daily survival rate of headstart nests ranged from 0.987 to 0.988 and the probability of a headstart nest surviving (i.e. parents did not abandon prior to the return of chicks to the nest) ranged from 71% to 74% among both sites and years (Table 2). Predation and over-wash were the two most common causes of identifiable nest loss for control nests, while headstart nests were lost primarily to these two factors as well as abandonment (Figure 3). During 2010, the anchors that secured artificial eggs in nests failed in 26% of headstart nests while in 2011 there were no cases of anchor failure. Abandonment at headstart nests often coincided with loss of artificial eggs.
Table 2. Daily survival rate and probability of nest success and brood success from control and headstart nests of American Oystercatchers in the Cape Romain Region, South Carolina, 2010–2011. AIWW = Atlantic Intracoastal Waterway, SWBB = Southwest Bulls Bay. DSR (± SE) and probability of success calculated using parameters from final logistic exposure models.

a No SE as final model did not include any significant independent variables hence survival estimate was constant across sites and years.

Figure 3. Mean (± SE) proportion of control (n = 90, filled circles) and headstart (n = 69, filled triangles) nests of American Oystercatchers, Cape Romain Region, South Carolina, April–July, 2010 and 2011, lost to over-wash, predation, abandonment, unknown causes, and failure to hatch. Note that ‘failure to hatch’ applies only to control nests (see Methods).
DSR of nests was greater at headstart compared to control nests (χ21 = 55.3, P < 0.0001; Figure 4). The odds of a headstart nest surviving an additional day were 5.3 times the odds of a control nest surviving an additional day. The DSR of control nests was significantly related to site (χ21 = 5.1, P = 0.02), year (χ21 = 4.4, P = 0.04), and tide height (χ21 = 6.5, P = 0.01). The odds of a nest surviving each day were 1.7 times greater in nests along the AIWW compared to SWBB and also were 1.6 times greater in 2010 compared to 2011. For every metre increase in high tide, a control nest was 9.6 times more likely to fail. The DSR of headstart nests was significantly related to nest age (χ21 = 6.8, P = 0.009). The mean nest age at failure for headstart nests was 17 ± 7.6 d. The odds of a headstart nest failing increased 1.1 times for each additional day it survived.

Figure 4. Daily survival rate (DSR) of control and headstart nests and broods (mean ± SE), and productivity (chicks per pair, mean ± SE) at control and headstart nests of American Oystercatchers, Cape Romain Region, South Carolina, April–July, 2010 and 2011. Note that both metrics appear on the same scale, and that the axis label includes both metrics.
There were no differences in attendance time (F1,71 = 0.01, P = 0.9) or percentage of time allocated to any behaviour between control and headstart nests during incubation (Table 3; F1,76 ≤ 1.5, P ≥ 0.2 for each). Percentage of time spent at nest during incubation was higher (F1,22 = 8.16, P = 0.01) at control nests that hatched ≥ 1 chick (88.8 ± 10.6%) compared to control nests that failed (77.6 ± 11.9%). No other behaviours were related to nest success at either control (F1,50 ≤ 3.2, P ≥ 0.09 for each behaviour) or headstart nests (F1,50 ≤ 1.2, P ≥ 0.3 for each behaviour).
Table 3. Percent time (mean SD) attending nests or chicks and engaged in specific behaviours on nesting territory during incubation (both study areas) and chick-rearing (AIWW only) within the Cape Romain Region, South Carolina, 2010–2011. Behavioural categories defined in Methods.

Step 3: Placement of headstart chicks in nest
Healthy headstart chicks (i.e. not displaying deformities, n = 48) were returned to 36 headstart nests between both sites and years. These chicks were returned to their original parents with no foster chicks in the brood (n = 23), returned to foster parents without original chicks (n = 7), or returned to their original parents with foster chicks (i.e. mixed parents, n = 6). All healthy chicks were accepted by parents (foster or original parents).
Step 4: Brood-rearing behaviour, chick survival, annual productivity
There was no difference in attendance rates (F1,14 = 0.06, P = 0.8) or percentage of time allocated to any behaviour between control and headstart nests during chick-rearing (Table 3; F1,14 ≤ 3.8, P ≥ 0.07 for each). No behaviours were related to brood success at either control (F1,5 ≤ 2.2, P ≥ 0.2 for each behaviour) or headstart nests (F1,7 ≤ 3.3, P ≥ 0.11 for each behaviour) along the AIWW.
Fifteen chicks fledged from 30 pairs from control nests in the AIWW and one chick fledged from 16 pairs from control nests in SWBB across both years (Table 1). Eleven chicks fledged from 30 pairs from headstart nests in the AIWW and two chicks fledged from 18 pairs from headstart nests in SWBB across both years (Table 1).
Brood success was greater at control nests compared to headstart nests (χ21 = 7.3, P = 0.007; Fig. 4). The odds of a control brood surviving an additional day were 16 times the odds of a headstart brood surviving. Survival of control broods was not affected by nest age, date, or tide (χ21 ≤ 0.2, P ≥ 0.7 for each). We did not include a variable for site or year for control broods because the sample size for control broods was unbalanced for site and year. Only site and nest day were significantly related to DSR of headstart broods. A headstart brood along the AIWW was approximately five times more likely to survive an additional day than a headstart brood in SWBB (χ21 = 12.3, P = 0.0005). The age of the brood was also related to DSR (χ21 = 8.7, P = 0.003). The odds of a brood surviving an additional day increased by 1.1 times for every day it survived.
Of the headstart chicks that did not survive to fledge along the AIWW (n = 15 in 2010, n = 12 in 2011), 60% were lost 0–7 days post-release, while in SWBB, 95% of headstart chicks that did not survive to fledge (n = 20) were lost 0–7 days post-release. No chicks were lost 28 days post-release. Causes of chick loss were difficult to assess because remains and signs of loss were not often observed on nesting territories. In the few cases where field signs were evident, it appeared that predation (n = 5) and starvation (n = 2) were contributing factors.
Productivity (chick/pair) in control nests ranged from 0 to 0.59 and in headstart nests from 0 to 0.41 (Table 1). There was no significant difference in productivity between control and headstart nests (paired t3 = 0.4, P = 0.7; Figure 4).
Discussion
Our goal was to increase the productivity of American Oystercatchers by artificially incubating eggs and placing newly hatched young in the nest, thereby avoiding loss of eggs and newly-hatched chicks to predation, over-wash, or other causes. We sought to do this because nest predation and nest flooding are primary causes of lost productivity (Jodice et al. Reference Jodice, Thibault, Collins, Spinks and Sanders2014), and in such cases headstarting through artificial incubation may be an appropriate management tool (Elliott et al. Reference Elliott, Merton and Jansen2001). We identified four broad steps and nine specific measurable outcomes in our headstart programme for oystercatchers. We rated five outcomes fully successful, two outcomes partially successful, and 2 outcomes not successful (Table 4).
Table 4. Feasibility of enhancing productivity of American Oystercatchers through a headstarting programme. Steps and measurable outcomes of the headstarting process are defined and classified as fully successful, partially successful, or not successful.

Step one, collection of eggs and artificial incubation, included two measurable outcomes; collection and transportation of eggs from nests and incubating eggs ex situ. No eggs were lost during collection or transport from the nest to the incubator, despite use of small boats for transportation, and therefore this outcome was rated as fully successful. The maximum time from collection to placement in the incubator was c.1 hour and the maximum distance eggs were transported was 9 km. Transporting eggs over a greater distance or for longer periods of time may require a transportation incubator that provides a stable thermal environment (Kuehler et al. Reference Kuehler, Lieberman, Oesterle, Powers, Kuhn, Kuhn, Nelson, Snetsinger, Herrmann, Harrity, Tweed, Fancy, Woodworth and Telfer2001, Robertson et al. Reference Robertson, Colbourne, Nelson and Westbrooke2005).
Ex situ hatching success was 70% in year one but 100% during year two for fertile eggs and therefore we conservatively rated this step as partially successful. Lower hatching success in 2010 included both unhatched eggs and eggs that hatched but with deformed chicks (e.g. ectopic viscera, splay legs). Two factors appeared to contribute to this effect. First was a short-term instability in the external environment that housed the incubator (i.e. a 24 h failure in the air-conditioning unit of the facility). Several eggs failed during this time. The external environment needs to remain relatively stable to allow the incubator to function correctly and therefore the facility should be as environmentally stable as possible (Klimstra et al. Reference Klimstra, Stebbins, Heinz, Hoffman and Kondrad2009).
The second factor was less than optimum incubator settings in year one. In 2010, the initial temperature and humidity were set at 37.6°C and 50% following recommendations for domestic poultry eggs. In 2011, however, we decreased the incubator temperature to 37.4°C and increased humidity to 65% following settings suggested for artificial incubation of shorebird eggs (Powell et al. Reference Powell, Cuthbert, Wemmer, Doolittle and Feirer1997). This change resulted in increased hatch success and minimal deformities in chicks. Higher rates of hatching can be obtained if humidity levels for eggs can be decreased during development (Klimstra et al. Reference Klimstra, Stebbins, Heinz, Hoffman and Kondrad2009). Our approach may have worked reasonably well for oystercatcher eggs because the structure of the egg itself (shell thickness, pore size and structure, cuticle thickness) promotes moisture loss during development. For example, eggs of Common Terns Sterna hirundo have a thin cuticle that appears to promote moisture loss during development (Klimsta et al. Reference Klimstra, Stebbins, Heinz, Hoffman and Kondrad2009). High rates of hatching during artificial incubation appear to be common among wild birds across an array of taxa once incubation settings are optimised (Kuehler et al. Reference Kuehler, Lieberman, McIlraith, Everett, Scott, Morrison and Winchell1993, Reference Kuehler, Lieberman, Oesterle, Powers, Kuhn, Kuhn, Nelson, Snetsinger, Herrmann, Harrity, Tweed, Fancy, Woodworth and Telfer2001, Powell et al. Reference Powell, Cuthbert, Wemmer, Doolittle and Feirer1997, Lobo and Marini Reference Lobo and Marini2013). However, incubation settings for wild birds are not well understood and natural incubation temperature and humidity are often not known or difficult to measure precisely in the field (Gonzalez-Garcia et al. Reference Gonzalez-Garcia, Porras and Vargas2006, Klimstra et al. Reference Klimstra, Stebbins, Heinz, Hoffman and Kondrad2009).
Step two, nest survival and parental behaviour during incubation, included three measurable outcomes; security of artificial eggs in nests, behaviour of parents at headstart compared to control nests, and DSR of headstart compared to control nests. In general, species that have been the subject of artificial incubation experiments appear to demonstrate low levels of nest abandonment when artificial eggs are secure and provided at the appropriate time in the nest cycle (King et al. Reference King, Espenshade, Kirkpatrick-Wahl, Lapinski, Malekan and Rickett2013, Lobo and Marini Reference Lobo and Marini2013). The placement of artificial eggs in headstart nests of oystercatchers was partially successful, with results improving in our second study year. At the start of the 2010 breeding season, artificial eggs were secured via string to 10 cm nails but were often dislodged, apparently by predators or waves as evidenced by teeth marks on artificial eggs and patterns of loss that coincided with wind and wave action. It did not appear that parents systematically removed eggs, and when we returned artificial eggs to nest sites and re-secured them within 1–2 d after displacement, they tended to remain in place. We revised the attachment process mid-way through the 2010 breeding season. We secured artificial eggs in nests with a 40-cm long rebar stake and artificial eggs were not dislodged from nests. When eggs were better secured, we also observed adults at headstart nests incubating artificial eggs after over-wash or predation events appeared to occur.
Rarely is parental behaviour at the nest monitored or reported for headstart programs and therefore potential underlying effects on reproductive ecology may be missed. Our headstart programme was fully successful with respect to minimising effects on incubation behaviour. We did not detect any difference in attendance rates or other behaviours of parents between headstart and control nests during incubation.
DSR for headstart nests was significantly greater compared to control nests, and therefore outcome 2.3 was rated fully successful. Despite the loss of some artificial eggs to anchor failure, and the abandonment of some headstart nests, DSR of headstart nests were consistently higher compared to control nests (although anchor failure may have contributed to the negative relationship between DSR and nest day for headstart nests). Flooding is a primary cause of nest loss for oystercatchers in our study area (Jodice et al. Reference Jodice, Thibault, Collins, Spinks and Sanders2014). The lack of a tide effect on headstart nests during our study, however, emphasises the success of the headstart program. Lastly, a longer term comparison of DSR of headstart nests to natural nests also confirms the success of the headstart program. Jodice et al. (Reference Jodice, Thibault, Collins, Spinks and Sanders2014) report a mean DSR of 0.938 for 354 nesting attempts in this study area, while the mean DSR for headstart nests in our study was 0.988.
Step three, the placement of chicks hatched in the incubator at nest sites, included one measurable outcome, the acceptance rate of headstart chicks when placed in oystercatcher nests. We rated this outcome as fully successful; all healthy chicks were accepted in nests whether returned to original or foster parents. The primary factor that affected whether parents accepted a headstart chick was the health of the chick. Adults did not accept chicks with significant deformities (e.g. splay legs or ectopic viscera). It is, however, not uncommon for avian parents to abandon unhealthy chicks and therefore this behaviour may not be directly related to headstarting (Lobo and Marini Reference Lobo and Marini2013). Whether chicks were placed in original or foster nests did not affect acceptance by parents. Acceptance of foster chicks appears to be common among many birds for which artificial incubation and chick placement has been attempted (Reed et al. Reference Reed, Nilsson and Murray1993, Lobo and Marini Reference Lobo and Marini2013). Adults may not, however, accept foster chicks if they are returned to the nest before the anticipated hatch date of original eggs. Therefore, nests of the appropriate age need be available for fostering and anticipated hatch dates need to be known (Lobo and Marini Reference Lobo and Marini2013). Cross-fostering may not be suited for small populations with high degrees of laying asynchrony (Lobo and Marini Reference Lobo and Marini2013).
Step four, brood-rearing and productivity, included three measurable outcomes; behaviour of parents with headstart chicks compared to control chicks, DSR of headstart compared to control broods, and productivity of headstart compared to control pairs. Our headstart program was fully successful with respect to minimising effects on brood-rearing behaviour. We did not detect any difference in attendance rates or other behaviours of parents between headstart and control broods. DSR of headstart broods and productivity of headstart pairs was not, however, significantly greater compared to control pairs and hence these two steps were deemed unsuccessful. In fact, DSR of headstart broods was less than that of control broods. The reasons underlying low DSR of broods and low productivity in headstart nests are unclear. Mortality of headstart chicks appeared most frequent during the first week post-release. Nevertheless we observed no reduction in parental attendance, vigilance, or provisioning in headstart broods that may have led to reduced chick survival. Our ability to monitor survival of headstart chicks was greater than that for control chicks as the former were outfitted with radio tags. Therefore, detectability was unlikely to contribute to a failure to observe and confirm survival of headstart chicks. It is possible that headstart chicks were burdened by radio tags and that this contributed to their higher mortality rate, although prior monitoring of radio-tagged chicks of American Oystercatchers has not resulted in high rates of mortality (http://www.fws.gov/FieldNotes/regmap.cfm?arskey=30718; Simons and Shulte Reference Simons and Schulte2010). It is also possible that additional handling of headstart chicks (e.g. to reattach transmitters) may have contributed to reduced survival, although prior monitoring of radio-tagged chicks of American Oystercatchers also required additional handling to re-secure transmitters and has not resulted in high rates of mortality. In general, however, brood success and productivity were low even at control nests during this study, hence decreasing the opportunity to make clear comparisons between control and headstart nests. For example, brood success averaged 75% in AIWW and SWBB during three breeding seasons (2006–2008; Jodice et al. Reference Jodice, Thibault, Collins, Spinks and Sanders2014) while during our study brood success at control nests in AIWW and SWBB averaged only 55%.
In summary, artificial incubation of eggs of American Oystercatchers was an effective tool for enhancing nest success. Ultimately, however, productivity at headstart nests remained low suggesting that although nest survival was enhanced, chick mortality remained a limiting factor. Low DSR of broods and subsequently poor productivity at headstart nests could occur if nest sites that are vulnerable to failure during incubation (e.g. high risk for flooding, mammalian predation, or avian predation) also function as poor habitat for chick-rearing. Because we assigned nests to the headstart group using an experimental approach, we were not able to consider territory quality as an assignment factor; doing so could enhance productivity at headstart nests. For example, a conservation action plan could evaluate territory quality prior to selecting a nest for headstarting as a means to increase the probability of a chick fledging. Therefore, obtaining background data on mechanisms that may contribute to chick mortality at the study site of interest (e.g. rates and timing of flooding or predation among nest sites and distribution and home range size of predators) would benefit a headstart program. Ultimately, employing a headstart program as the sole mechanism to increase productivity for American Oystercatchers may be most appropriate if productivity is lost primarily during the incubation stage.
Acknowledgements
We thank Mark Spinks, Sarah Woodward, Billy Shaw, Charles Wahl, Adam DiNuovo, Nicholas Wallover, J. Benjamin Mongold, and the staff at the Charles Lee Morgan Poultry Center at Clemson University, particularly Carol Foster-Mosely, for their assistance and advice. Carolyn Wakefield assisted with logistics and administration. We also acknowledge the support of Cape Romain National Wildlife Refuge, in particular, Sarah Dawsey. This research was funded and supported by the National Fish and Wildlife Foundation, US Fish and Wildlife Service, South Carolina Department of Natural Resources, USGS South Carolina Cooperative Fish and Wildlife Research Unit and Clemson University. The manuscript benefitted from comments by Patrick Gerard and David Jachowski. Permits for this research were provided by The Clemson University Institutional Animal Care and Use Committee (protocol # 2010-028). The South Carolina Cooperative Fish and Wildlife Research Unit is supported by the South Carolina Department of Natural Resources, Clemson University, the U.S. Fish and Wildlife Service, and the U.S. Geological Survey. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.








