



Non-technical Summary
In southern Guizhou, South China, there are well-exposed shallow-water deposits of the Jiusi and Shangsi formations, which consist of platform limestones mixed with shales and sandstones. Conodonts, ancient tooth-like fossils, were found in the limestones and were dominated by two groups: the Cavusgnathus aff. Cav. unicornis and Clydagnathus windsorensis in the upper Jiusi Formation and the Cavusgnathus unicornis in the lowest Shangsi Formation. Vogelgnathus campbelli with small lateral denticles was found for the first time in Eastern Paleotethys. The Cavusgnathus unicornis is used as a marker for the regional Shangsian stage and correlated to other conodont zones marked by similar species in North America and Europe of middle Visean.
Introduction
Carboniferous shallow-water conodont assemblages from open marine shallow-water environments (e.g., carbonate platform and outer shelf) are typically diverse and accompanied by other benthic taxa such as corals, brachiopods, and foraminifers (e.g., the late Tournaisian conodonts with short ranges; Tian and Coen, Reference Tian and Coen2005). More-restricted environments (e.g., inner shelf, back reef, and lagoon) are highly influenced by sea level, temperature, and salinity, which makes these conodont faunas different from their open-marine counterparts. Genera belonging to Cavusgnathidae (mentioned in the following) are the typical representatives of the restricted shallow-water-environment faunas (Austin and Davies, Reference Austin and Davies1984). Although they may be found in deeper-water environments, such as slopes and outer shelves, they are uncommon and make up a small portion of these assemblages. In these cases, reworking of these fossils due to downslope transport should be considered (Austin and Davies, Reference Austin and Davies1984). The globally recognized Cavusgnathus biofacies (Mississippian) and Adetognathus biofacies (Pennsylvanian) are based on these dominant euryhaline genera (Merrill and Martin, Reference Merrill and Martin1976; Davis and Webster, Reference Davis and Webster1985). Their early homomorphic relatives Clydagnathus and Patrognathus represent similar facies during Late Devonian (Sandberg and Dreesen, Reference Sandberg and Dreesen1984). Taphrognathus is also very useful in correlation of Visean sequences (Purnell et al., Reference Purnell, von Bitter and Groessens2002).
China was composed of several independent tectonic blocks during the Carboniferous. Most of these tectonic units were scattered around the eastern rim of the Paleotethys Ocean in low- and middle-latitude belts (e.g., Am, An, NC, SC, Si, T, and Ti in Fig. 1). Diverse conodont assemblages are documented from different depositional settings along a dip profile. Salinity appears to be one of the most important ecological factors for the Carboniferous conodonts in China because high salinity levels are associated with low-diversity faunas (Wang and Clark, Reference Wang and Clark1990). Most of the Carboniferous shallow-water conodont faunas in China are reported from the Mississippian. A conodont zonation based on index species of Siphonodella has been recognized in Guizhou, Hunan, Guangxi, and Guangdong and is a useful tool for regional correlation of the early Tournaisian successions (Ji, Reference Ji1987a; Ji and Ziegler, Reference Ji and Ziegler1992; Qie et al., Reference Qie, Wang, Zhang, Ji, Grossman, Huang, Liu and Luo2016). In Yunnan and Hunan, Finognathodus and Dollymae are shallow-water benthonektonic representatives of the late Tournaisian and probably early Visean (Ji, Reference Ji1987b; Coen et al., Reference Coen, Groessens and Sevastopulo2004; Tian and Coen, Reference Tian and Coen2005). There are some scattered records of Clydagnathus, Bispathodus, and Rhachistognathus from Tarim and Junggar areas in Xinjiang (Zhao et al., Reference Zhao, Zhang and Xiao2000), but most of them are from borehole data without reliable stratigraphic control. Species of Neognathodus from Xainza, Tibet, may be shallow-water representatives of Bashkirian and Moscovian strata in the Lhasa block (Ji et al., Reference Ji, Yao, Wu, Liu, Jiang and Fu2007).

Figure 1. Paleogeographic reconstructions of the globe (Am = Amuria; An = Annamia; NC = North China; SC = South China; Si = Sibumasu; T = Tarim; Ti = Tibet; modified from Torsvik and Cocks, Reference Torsvik and Cocks2017) and South China (DQGX Platform = Yunnan–Guizhou–Guangxi–Hunan Platform; QG Basin = Guizhou–Guangxi Basin; C1 = Mississippian; C2 = Pennsylvanian; P = Permian; modified from Wu, Reference Wu1987; Feng et al., Reference Feng, Yang, Bao, Jin, Zhang, Wu and Qi1998) during Visean to Serpukhovian.
The reconstructed temperature shows that the Visean may have been a relatively warm period before the main stage of the late Paleozoic ice age (Yao et al., Reference Yao, Aretz, Wignall, Chen, Vachard, Qi, Shen and Wang2020). This was inferred mainly from the compiled oxygen isotope curve based on calcitic brachiopod shells (Veizer and Prokoph, Reference Veizer and Prokoph2015). Statistics on the distributions and dimensions of the glaciers and reef-like bioconstructions also support such deduction (for more references, see Isbell et al., Reference Isbell, Henry, Gulbranson, Limarino, Fraiser, Koch, Ciccioli and Dineen2012; Yao et al., Reference Yao, Aretz, Wignall, Chen, Vachard, Qi, Shen and Wang2020). Shallow-water environments are widely distributed in South China with abundant macrofaunas. To clarify the coevolution of life and environments, it is important to develop techniques that allow precise stratigraphic correlation. The foraminifer-based stratigraphy in shallow-water deposits is conventionally used, but the scarcity of shallow-water taxa in deep-water deposits makes the correlation between the two realms difficult. The nektobenthic conodont species are highly cosmopolitan, less diverse, and provide clearer correlation across different lithological facies. The conodont assemblages described here are from the restricted shallow-water environment at Yashui, Guizhou, and are dominated by cavusgnathids. Similar assemblages in South China have been reported previously only from the boreholes in Shaoguan, Guangdong (Ding and Wan, Reference Ding and Wan1989). These new conodont assemblages elucidate the evolution of Cavusgnathus unicornis Youngquist and Miller, Reference Youngquist and Miller1949, which is an important species for biostratigraphic correlation, from Cavusgnathus aff. Cav. Unicornis Youngquist and Miller, Reference Youngquist and Miller1949, allowing for a proposed global correlation based on the reported taxa.
Geological setting
The South China Block is located in the northeastern Paleotethys Ocean at low latitude, and the old lands of Yangtze and Cathaysia were connected during the Visean to Serpukhovian (Fig. 1). Marine-to-terrestrial alternations are widely recognized in the present northeastern regions such as Jiangsu, Anhui, and Hubei provinces (e.g., the Gaolishan Formation, mainly shales and siliciclastics with brachiopods and plants). In the southwestern regions, carbonates vary between nearshore (e.g., the Jiusi and Shangsi formations, intercalated with shales and abundant macrofossils) and basinal (e.g., the Nandan Formation, intercalated with siliceous cherts and few macrofossils) settings. Classic separation of the carbonates in Guizhou was based partly on their distinct colors, with the “white” and “black” carbonates indicating the shallow-water and deep-water deposits, each with mainly benthic and planktic organisms, respectively (Xiong and Zhai, Reference Xiong and Zhai1985). The regional subdivisions of the Carboniferous in China are based mainly on the shallow-water sections from the white carbonate regions (Shen and Wang, Reference Shen and Wang2015).
The section at Yashui is well exposed along the road near the town of Yashui and consists of the typical white carbonate and some shaly strata (Fig. 2). The composite stratigraphic section is composed of three outcrops: A, B, and C, in ascending order, and consists of the Jiusi, Shangsi, Baizuo, and Huashiban formations (Wu, Reference Wu1987). The continuous sections B and C are the reference sections of the regional Dewuan Stage, characterized by the thick limestone with abundant corals and brachiopods (Wu, Reference Wu, Wang and Huang2008; Lin et al., Reference Lin, Wang, Poty and Aretz2012; Lin, Reference Lin2013). The beds with paleokarsts or storm-sorted brachiopods may indicate intervals of disturbed deposition (Chen et al., Reference Chen, Montanez, Qi, Wang, Wang and Lin2016; Yao et al., Reference Yao, Aretz, Li and Wang2016a). Conodonts are rare in this section, and foraminifers are the primary biochronostratigraphic indexes. The Visean–Serpukhovian boundary was tentatively put within a paleokarst bed in the upper part of Shangsi Formation, about 5 m below the regional Shangsian–Dewuan boundary (Wu, Reference Wu, Wang and Huang2008; Groves et al., Reference Groves, Wang, Qi, Richards, Ueno and Wang2012; Chen et al., Reference Chen, Montanez, Qi, Wang, Wang and Lin2016).

Figure 2. Outcrop pictures of the Yashui-A section. (1) Distant view of the section. (2) Detail of the section showing the boundary between the Jiusi Formation and the Shangsi Formation. (3) Typical lithostratigraphic succession of the Jiusi Formation.
The studied interval consists of section A (Yashui-A; 25.9889°N, 106.7501°E). The top of section A is correlated with the bottom of section B by a marker bed with a fossiculate coral biostrome. It consists of the upper part of Jiusi Formation and the lowermost part of Shangsi Formation (Wu, Reference Wu1987) (Fig. 2). The upper Jiusi Formation is composed mainly of gray to dark gray, thick-bedded limestones and shaly marls intercalated with a few sandstones. Brachiopods, corals, gastropods, and crinoids are abundant in pure limestones and sometimes also in laminated marls. Foraminifer assemblages with Eoparastaffella, Dainella, and Archaediscus can be roughly correlated with the MFZ11 to MFZ12 foraminiferal zones in Western Europe (Wu, Reference Wu1987; Poty et al., Reference Poty, Devuyst and Hance2006). The lower Shangsi Formation is composed of a higher number of limestone beds with lesser shales. The boundary between the Jiusi and Shangsi formations (roughly correlated with the regional Jiusian stage and Shangsian boundary) at this site is marked by a distinct coral biostrome bed (Wu, Reference Wu1987; Yao et al., Reference Yao, Wang, Lin, Li, Kershaw and Qie2016b, and our bed 100 in Fig. 3). New foraminifera data from this biostrome bed and the overlying strata include Pojarkovella nibelis (Durkina, Reference Durkina1959), Koskinobigenerina breviseptata Eickhoff, Reference Eickhoff1968, and Koskinotextularia cribriformis Eickhoff, Reference Eickhoff1968, referring to the MFZ12 foraminiferal zone of the middle Visean (Yao et al., Reference Yao, Wang, Lin, Li, Kershaw and Qie2016b).

Figure 3. Lithostratigraphic column and conodont distributions of the Yashui-A section. F = formation; S = shaly marls; M = mud limestone; W = wackestone; P = packstone; B = biostrome.
Methods and materials
We sampled every limestone bed within the 65 m outcrop, spanning the boundary of the Jiusi and Shangsi formations. In total, 58 conodont samples (4–5 kg each) were collected and processed by acetic acid dissolution (~10% vol) and heavy-liquid separation (ρ ~2.8 g/mL). Conodonts are rare, and the diagnostic specimens were extracted from only 18 samples (Fig. 3). All of the specimens are black in color (CAI 4~5), and most of them are Pa pectiniform elements, which are the most robust elements in the conodont apparatus. Discrete fragments of the ramiform elements are common, and this may indicate a high-energy environment where less-robust elements were crushed and washed away. The specimens were photographed by a Hitachi SU3500 scanning electron microscope (SEM) after gold coating.
Repository and institutional abbreviation
The illustrated specimens are stored in Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (NIGPAS), Nanjing, China, under the catalog numbers NIGP 201189–201236.
Systematic paleontology
In this paper, the systematic study is based mainly on the Pa elements because other elements are scarce and not well preserved. We follow Sweet (Reference Sweet1988) and Purnell (Reference Purnell1992) for the classification and detailed synonyms.
Phylum Chordata Bateson, Reference Bateson1886
Class Conodonta Pander, Reference Pander1856
Subclass Conodonti Branson, Reference Branson1938
Order Ozarkodinida Dzik, Reference Dzik1976
Family Cavusgnathidae Austin and Rhodes in Robison, Reference Robison1981
Remarks
Cavusgnathidae was erected by Austin and Rhodes (Robison, Reference Robison1981) and originally included seven genera: Cavusgnathus Harris and Hollingsworth, Reference Harris and Hollingsworth1933, Adetognathus Lane, Reference Lane1967, Clydagnathus Rhodes, Austin, and Druce, Reference Rhodes, Austin and Druce1969, Patrognathus Rhodes, Austin, and Druce, Reference Rhodes, Austin and Druce1969, Taphrognathus Branson and Mehl, Reference Branson and Mehl1941, Capricornognathus Austin in Austin and Mitchell, Reference Austin and Mitchell1975, and Cloghergnathus Austin in Austin and Mitchell, Reference Austin and Mitchell1975. According to the current study, this family includes eight genera. Cloghergnathus and Capricornognathus are considered junior synonyms of Patrognathus and Taphrognathus, respectively (Purnell, Reference Purnell1992), and the endemic genus Montognathus Crane in Jenkins et al., Reference Jenkins, Crane and Mory1993 that was recognized from eastern Australia is included. Dzik (Reference Dzik2006) noted that this family may be polyphyletic and includes three early genera: Pseudopolygnathus Branson and Mehl, Reference Branson and Mehl1934, Alternognathus Ziegler and Sandberg, Reference Ziegler and Sandberg1984 and Scaphignathus Helms, Reference Helms1959. This extends the family to the mid-Famennian of the Late Devonian. The conventional terms of “left” and “right” used here refer to the relative locations of certain parts when the cavusgnathid Pa element is situated venter (anterior) up and dorsal (posterior) down in oral view.
Genus Cavusgnathus Harris and Hollingsworth, Reference Harris and Hollingsworth1933
Type species
Cavusgnathus alta Harris and Hollingworth, Reference Harris and Hollingsworth1933.
Remarks
As the first described cavusgnathid, early interpretations of Cavusgnathus had a wide range of variation and a nearly global distribution. Since the erection of new morphological groups, many species originally included in this genus have been reassigned (e.g., Lane, Reference Lane1967 considered all representatives of the Pennsylvanian Cavusgnathus to belong to his new genera Adetognathus). Although the phylogeny of these similar genera is far from being resolved (Lane, Reference Lane1967; Rhodes et al., Reference Rhodes, Austin and Druce1969; von Bitter and Austin, Reference von Bitter and Austin1984; Brown et al., Reference Brown, Rexroad, Beard and Williams1990; Jenkins et al., Reference Jenkins, Crane and Mory1993), the morphological differences in Pa elements are relatively distinct. The Pa element of Cavusgnathus has a shorter blade and shows Class IV or Class IIIa symmetry while Adetognathus shows Class II or IIIb and Taphrognathus shows Class II or III symmetry (Lane, Reference Lane1967; Purnell, Reference Purnell1992). Cavusgnathus differs from Clydagnathus in having platform ornament with ridges rather than discrete nodes. The reassignment of Cly. hudsoni Metcalfe, Reference Metcalfe1981 to Cavusgnathus extended the range of this genus into the late Tournaisian.
Cavusgnathus unicornis Youngquist and Miller, Reference Youngquist and Miller1949
Figures 4.27–4.35, 5.1–5.8

Figure 4. Conodonts from Yashui-A section (1, 10–22, 25, 27–30, 33, 34 are sinistral Pa elements; 2–9, 23, 24, 26, 31, 32, 35 are dextral Pa elements). (1–26) Cavusgnathus aff. Cav. unicornis Youngquist and Miller, Reference Youngquist and Miller1949: (1, 2, 7, 8, 10–14) 201189, 201190, 201195, 201196, 201198, 201198, 201198, 201199, 201199, bed 63; (3) 201191, bed 1; (4, 26) 201192, 201192, bed 55; (5, 6) 201193, 201194, bed 57; (9, 15) 201197, 201200, bed 67; (16–18) 201201, 201202, 201202, bed 69; (19, 20) 201203, 201203, bed 91; (21–24) 201204, 201204, 201205, 201206, bed 99; (25) 201207, bed 53. (27–35) Cavusgnathus unicornis Youngquist and Miller, Reference Youngquist and Miller1949: (27, 28) 201208, 201208, bed 100b; (29, 33–35) 201209, 201213– 201215, bed 104; (30–32) 201210–201212, bed 103. (1–11, 13–35) Scale bars = 300 μm; (12) scale bar = 50 μm.

Figure 5. Conodonts from Yashui-A section (1, 2, 5, 6, 14–17, 26, 29–31 are sinistral Pa elements; 3, 4, 7–13, 18–20, 24, 25, 27, 28, 32, 33 are dextral Pa elements; 21, 22 are Pb elements; 23 is S element). (1–8) Cavusgnathus unicornis Youngquist and Miller, Reference Youngquist and Miller1949: (1, 2, 4–6, 8) 201216, 201217, 201219, 201216, 201217, 201219, bed 105; (3, 7) 201218, 201218, bed 104. (9) Hindeodus cristulus (Youngquist and Miller, Reference Youngquist and Miller1949), 201220, bed 57. (10–20, 24, 25) Clydagnathus windsorensis (Globensky, Reference Globensky1967): (10, 12) 201221, 201221, bed 57; (11, 13, 14) 201222, 201222, 201223, bed 51; (15–18, 24) 201224–201227, 201227, bed 63; (19, 20, 25) 201228, 201229, 201228, bed 69. (21) Ozarkodina sp., 201230, bed 69. (22) Ozarkodina sp., 201231, bed 63. (23) Apatognathus sp., 201232, bed 105. (26–33) Vogelgnathus campbelli (Rexroad, Reference Rexroad1957): (26–28, 30–33) 201233–201235, 201233, 201233–201235, bed 104; (29) 201236, bed 103. (1–24, 26–29) Scale bars = 300 μm; (25, 31–33) scale bars = 30 μm; (30) scale bar = 100 μm.
- Reference Youngquist and Miller1949
Cavusgnathus unicornis Youngquist and Miller, Reference Youngquist and Miller1949, p. 619, pl. 101, figs. 18–23 (Pa element, α morphotype). [Iowa]
- Reference Youngquist and Miller1949
Cavusgnathus regularis Youngquist and Miller, p. 619, pl. 101, figs. 24, 25 (Pa element, β morphotype). [Iowa]
- Reference Rexroad1957
Cavusgnathus convexa Rexroad, p. 17, pl. 1, figs. 3–6 (Pa element, γ morphotype). [Illinois]
- Reference Rexroad1957
Ozarkodina compressa Rexroad, p. 36, pl. 2, figs. 1, 2 (Pb element). [Illinois]
Holotype
The University of Iowa 4174 from Pella beds at Pella South West, Marrion County, Iowa, USA (Youngquist and Miller, Reference Youngquist and Miller1949, pl. 101, figs. 18, 19).
Occurrence
Lower Shangsi Formation in the Yashui-A section, Guizhou; mid-Visean.
Description
Both dextral and sinistral Pa elements of Cavusgnathus unicornis are recognized at Yashui. There is a difference between them, thus a Class IIIa symmetry is supported. The ventral (anterior) blade is about 35%–40% of the whole element length and occupied mostly by the fixed part. Three to eight denticles are on the blade, laterally compressed and normally with lower part fused. The dextral element has fewer denticles (usually three to five) on the relatively shorter blade. In lateral view, the blade height is greater than length, and we accept the typical α (e.g., Fig. 4.32), β (e.g., Fig. 4.27, 4.28), γ (e.g., Fig. 5.1–5.8), or other intergradational morphotypes of C. unicornis summarized by Rexroad (Reference Rexroad1981). The blade is attached to the caudal side for sinistral elements and to the rostral side for dextral elements. Thus, it always appears that the blade connects to the “right” side. The junction between the blade and the parapet is smooth for dextral elements but has a slight notch in most sinistral elements. Both parapets are ornamented with transverse ridges and usually expanded most at the dorsal two-thirds of the whole length. The flat platform in the dextral element can be clearly seen from the “left”-lateral view (Fig. 5.7, 5.8) while in the sinistral element, the “left” parapet is uplifted with less lateral expansion and upper outline of the platform seems gradually arched (Fig. 5.5, 5.6). The medial trough is very shallow ventrally where the “left” parapet decreases in height. Several linear-situated nodes may fill in the dorsal trough and extend as a dorsal blade. The parapets may occasionally taper at the end (Fig. 4.30). The basal cavity is fully open and widest near the middle length of the element. The basal cavity is slightly asymmetric with the caudal (inner) half extending farther ventrally, and this may help to recognize the chirality of the elements.
Materials
Thirteen Pa elements, including some broken specimens (201208–201219).
Remarks
Cavusgnathus unicornis from Yashui contains dextral and sinistral elements of various blade morphotypes. The comparable species Cav. hudsoni was commonly found in older strata of the Tournaisian and was originally assigned to Clydagnathus (Metcalfe, Reference Metcalfe1981). Cav. hudsoni has a relatively longer free blade and a trough that may close ventrally, which is different from all other Cavusgnathus species. Cav. naviculus (Hinde, Reference Hinde1900) is also a cosmopolitan species but was found in younger strata, mostly Serpukhovian, with a nearly filled medial trough. The platform of Cav. altus (i.e., Cav. alta) has a rather flat upper outline in lateral view and a basal cavity not extended to the dorsal end. Cav. charactus Rexroad, Reference Rexroad1957 is diagnosed by the significant notch between the blade and parapet, which is not prominent in Cav. unicornis.
Cavusgnathus aff. Cav. unicornis Youngquist and Miller, Reference Youngquist and Miller1949
Figure 4.1–4.26
- ? Reference von Bitter and Plint-Geberl1982
Cavusgnathus regularis type Youngquist and Miller, Reference Youngquist and Miller1949; von Bitter and Plint-Geberl, Reference von Bitter and Plint-Geberl1982, pl. 3, figs. 14, 17, 18 only (Pa elements).
- ? Reference von Bitter and Austin1984
Cavusgnathus unicornis Youngquist and Miller, Reference Youngquist and Miller1949; von Bitter and Austin, Reference von Bitter and Austin1984, pl. 19, figs. 12–14, 16–28 only (Pa elements).
Occurrence
Upper Jiusi Formation in the Yashui-A section, Guizhou; mid-Visean.
Description
Cavusgnathus aff. Cav. unicornis from Yashui shows Class IIIa symmetry. The ventral blade is about 25%–35% of the whole element length with very small free part. One to five laterally compressed denticles lie on the blade with their most basal part fused. The sinistral element usually has fewer denticles as one or two large denticles. The blades of the specimens show similarity of the α (e.g., Fig. 4.6–4.8, 4.13, 4.14, 4.19, 4.20, 4.24), β (e.g., Fig. 4.9–4.12), and intermediate morphotypes of the Cav. unicornis. The ventral blade is always close to or connected with the “right” parapet but usually more medially situated. Thus, a notch commonly occurs between the blade and “right” parapet because of this misalignment, and the dorsal end of the blade ends abruptly (e.g., Fig. 4.10–4.14, 4.21–4.25). The element is slim in general, ornamented with transverse ridges on both parapets, and a trough almost straight in the sinistral and slightly twisted in the dextral. The parapets in the dextral element constrict at the middle length and expand at the dorsal part when the specimens are small (e.g., Fig. 4.6, 4.7); in larger specimens, these parts are more ventrally situated (e.g., Fig. 4.23). In the sinistral element, the “left” parapet is uplifted and the “right” parapet is laterally expanded. The widest place of the platform is at the middle length. Dorsal blades in some specimens are significantly extended beyond the platform (Fig. 4.24, 4.25).
Materials
Twenty-three Pa elements, including some broken specimens (201189–201207).
Remarks
In all aspects, Cavusgnathus aff. Cav. Unicornis is similar to Cav. unicornis, but the relatively low number of specimens makes it difficult to understand what morphological changes should be considered interspecific. Because the diagnostic characteristics of Cavusgnathus aff. Cav. unicornis are restricted stratigraphically, we maintain the distinction between the two and interpret them as closely related species. The Pa element of Cavusgnathus aff. Cav. unicornis has a slimmer platform with relatively shorter blade and more significant parapet notch. These characters also show similarity with Cav. hudsoni, but the latter has a relatively longer free blade (more information in Purnell, Reference Purnell1992, table 2). The morphometric changes from small specimens to large specimens may be explained by allometric ontogeny, in which the dorsal part of platform grows faster than the ventral in Cavusgnathus. Thus, the length proportion of the ventral blade may decrease during growth as shown in Figure 6.

Figure 6. Plot of the dimensional ratios of Cavusgnathus unicornis and Cavusgnathus aff. Cav. unicornis
The lineage from Taphrognathus to Cavusgnathus (i.e., Cav. unicornis) has long been suggested on the basis of specimens from the lower and upper St. Louis formations (Rexroad and Collinson, Reference Rexroad and Collinson1963) in the United States and from Avon Gorge (von Bitter and Austin, Reference von Bitter and Austin1984) in the United Kingdom. However, after the reassignment of Cly. hudsoni as the earliest Cavusgnathus, the former lineage seems unreliable stratigraphically, thus a lineage from Clydagnathus to Cavusgnathus early forms was suggested (Purnell, Reference Purnell1992). Because no small specimens were collected, the ontogenetic patterns proposed by Austin (Reference Austin1973) and Purnell (Reference Purnell1992) could not be tested. Jenkins et al. (Reference Jenkins, Crane and Mory1993) considered the right attachment of the blade and the openness of the ventral trough as two successive phylogenetic modifications in Cavusgnathus.
A new species, Cav. altifrons Jenkins in Jenkins et al., Reference Jenkins, Crane and Mory1993, has been recognized as the intermediate between Cav. hudsoni and other Chesterian Cavusgnathus species (e.g., Cav. unicornis). Cav. altifrons has a similar platform shape and parapet notch. The dorsal blade in Cav. Altifrons is also quite like those of some of our specimens (e.g., Fig. 4.24, 4.25). Cav. altifrons differs from Cavusgnathus aff. Cav. unicornis by having a relatively longer free blade, smaller basal cavity, parapets ornamented with nodes rather than transverse ridges, and in lateral view the “left” parapet joints with the ventral blade at about half the height of the blade. Meanwhile, Cav. altifrons seems to lack a clear chirality.
Considering the dimensional ratios plot by Jenkins et al. (Reference Jenkins, Crane and Mory1993), Cavusgnathus aff. Cav. unicornis may be morphologically similar to Cav. unicornis and Cav. charactus. Since there are few illustrations of Cav. charactus, it is difficult to make a comparison. The Pb element of Cavusgnathus aff. Cav. unicorns may be represented by two Ozarkodina sp. found within the same stratigraphic range. Our Ozarkodina sp. is similar to the holotype of Ozarkodina compressa (thought as the Pb element of Cav. unicornis) but with less-fused denticles and more arched profile in lateral view (Fig. 5.21, 5.22). Thus, it is more similar to the older Pb element of Cav. hudsoni illustrated by Purnell (Reference Purnell1992; pl. 1, fig. 9).
The early Cavusgnathus record in South China is from the upper Jinling Formation, accompanied with the late Tournaisian species Polygnathus communis communis (Ni, Reference Ni, Feng, Xu, Lin and Yang1984, identified as Cly. unicornis Rhodes, Austin, and Druce, Reference Rhodes, Austin and Druce1969, but the single illustrated specimen is more like Cav. altifrons or Cav. hudsoni and reassigned to Cavusgnathus). Recently, several similar specimens have been found from Guizhou and Guangxi, with Polygnathus (probably Polygnathus dujieensis Qie et al., Reference Qie, Zhang, Du, Yang, Ji and Luo2014 spanning the Tournaisian– Visean boundary) (unpublished). These specimens are ornamented with elongated nodes or transverse ridges on both parapets. The difference between Cav. altifrons and Cav. hudsoni is blurred, if only by these ornaments on parapets, and the low-resolution SEM figures may also cause such confusion. Because of the preceding comparison, Cavusgnathus aff. Cav. unicornis is considered as an intermediate between Cav. hudsoni and Cav. unicornis.
Genus Clydagnathus Rhodes, Austin, and Druce, Reference Rhodes, Austin and Druce1969
Type species
Clydagnathus cavusformis Rhodes, Austin, and Druce, Reference Rhodes, Austin and Druce1969.
Remarks
When Rhodes et al. (Reference Rhodes, Austin and Druce1969) recognized this genus from early Tournaisian strata, they distinguished it from Cavusgnathus by the merging of the parapet and ventral blade, the closure of the medial trough ventrally, and a laterally, rather than longitudinally, expanded basal cavity. Later conodont workers found these characters not practical and proposed using parapet ornaments with nodes rather than ridges to distinguish them from Cavusgnathus instead (Nicoll and Druce, Reference Nicoll and Druce1979; von Bitter and Plint, Reference von Bitter and Plint1987). The long-ranging Clydagnathus may be polyphyletic (Rhodes et al., Reference Rhodes, Austin and Druce1969; Sandberg and Ziegler, Reference Sandberg and Ziegler1979; Purnell, Reference Purnell1992). The reassignment of Cav. windsorensis Globensky, Reference Globensky1967 to Clydagnathus extends the range of this genus into Visean.
Clydagnathus windsorensis (Globensky, Reference Globensky1967)
Figure 5.10–5.20, 5.24, 5.25
- Reference Globensky1967
Cavusgnathus windsorensis Globensky, p. 439, pl. 57, figs. 3, 4, 7, 9, 11, 19; pl. 58, fig. 1 (Pa element).
- Reference Globensky1967
Cavusgnathus cf. windsorensis Globensky, Reference Globensky1967, p. 439, pl. 57, figs. 2, 6, 10, 12; pl. 58, fig. 8 (Pa element).
- ? Reference Briggs, Clarkson and Aldridge1983
Clydagnathus? cf. cavusformis Rhodes, Austin, and Druce, Reference Rhodes, Austin and Druce1969; Briggs, Clarkson, and Aldridge, p. 3, figs. 1–3 (whole animal).
- Reference von Bitter and Plint1987
Clydagnathus windsorensis (Globensky, Reference Globensky1967); von Bitter and Plint, p. 350, pl. 57, figs. 2.1–2.7, 2.10, 2.11, 2.14–2.17 (Pa element).
Holotype
University of New Brunswick 64-F-235 from Windsor Limestone at Skir Dhu, Cape Breton, Nova Scotia, Canada (Globensky, Reference Globensky1967, pl. 57, figs. 3, 4, 7).
Occurrence
Upper Jiusi Formation in the Yashui-A section, Guizhou; mid-Visean of the Mississippian.
Description
The Pa element has a lanceolate platform with parapets ornamented by discretely spaced nodes. The nodes are laterally elongated in oral view and pointed in lateral view. The ventral blade is a little less than one-third of the whole element, with the height greater than the length. Two to six laterally compressed blade denticles are present, almost fully fused at the base. Only the last two denticles are easily recognizable. The ventral blade is almost free and medially situated. The ventral blades connect with the “right” parapet for both sinistral and dextral elements, and this Class IIIa symmetry is distinguishable in oral view (e.g., Fig. 5.16, 5.18). The medial trough is closed ventrally by the merging of the “left” parapet with the blade and dorsally by the carina and dorsal blade. Occasionally, the elongated nodes on the parapets may connect with the carina and cross at the trough (Fig. 5.19, 5.25). The basal cavity is mainly restricted at the middle length of the element. The stoutest and slimmest specimens studied here are considered as two ends of the morphometric range of this species (Fig. 5.10–5.13), and the similar situation of Cly. unicornis Rhodes, Austin, and Druce, Reference Rhodes, Austin and Druce1969 has been illustrated by Varker and Higgins (Reference Varker and Higgins1979, pl. 16, figs. 8, 9).
Materials
Eighteen Pa elements, including some broken specimens (201221–201229).
Remarks
Clydagnathus windsorensis has limited records outside of North America. There is a long temporal gap between this species and its early counterparts. Purnell (Reference Purnell1992) considered it as a possible progenetic offshoot of Cavusgnathus. The known conodont animal Clydagnathus? cf. Cly. cavusformis might belong to this species (Briggs et al., Reference Briggs, Clarkson and Aldridge1983). The possible Pb element is similar to that of Lochriea rather than Cavusgnathus (Ozarkodina sp. A. in Globensky, Reference Globensky1967, pl. 55, figs. 1, 5, 12), and a cladistic study based on the complete apparatus may yield similar results (Donoghue et al., Reference Donoghue, Purnell, Aldridge and Zhang2008). Unfortunately, no similar Pb element was recovered. Compared with the published materials, our Pa element shows intermediate characters of the parapet denticles between the typical Clydagnathus and Cavusgnathus, which probably adds more confusion on the phylogeny of this genus.
Family Uncertain
Genus Vogelgnathus Norby and Rexroad, Reference Norby and Rexroad1985
Type species
Spathognathodus campbelli Rexroad, Reference Rexroad1957.
Remarks
The apparatus of this genus is based on bedding plane materials (Norby and Rexroad, Reference Norby and Rexroad1985; Purnell and Donoghue, Reference Purnell and Donoghue1997; Sanz-Lopez et al., Reference Sanz-Lopez, Blanco-Ferrera and Miller2019). Vogelgnathus is similar to several genera (e.g., Lochriea, Gnathodus, and Hindeodus) but is easily differentiated by the Pa elements (Norby and Rexroad, Reference Norby and Rexroad1985; Sweet, Reference Sweet1988; Donoghue et al., Reference Donoghue, Purnell, Aldridge and Zhang2008). Dzik (Reference Dzik2006) included several Devonian species within this genus and under the family Francodinidae. The evolution of this genus is not clear. As for the conventional Mississippian representatives, the phylogeny pattern is probably complicated by geographical differentiation (Purnell and von Bitter, Reference Purnell and von Bitter1992a; Sanz-Lopez et al., Reference Sanz-Lopez, Blanco-Ferrera and Miller2019).
Vogelgnathus campbelli (Rexroad, Reference Rexroad1957)
Figure 5.26–5.33
- Reference Rexroad1957
Spathognathodus windsorensis Rexroad, p. 37, pl. 3, figs. 13–15 (Pa element).
- Reference Norby and Rexroad1985
Vogelgnathus campbelli (Rexroad, Reference Rexroad1957); Norby and Rexroad, p. 3, pl. 1, figs. 1, 2 (assemblage), pl. 2, figs. 3–10 (Pa element), pl. 2, figs. 1, 2, pl. 3, figs. 5, 9, 10 (Pb element).
Holotype
Illinois State Geological Survey 2P75 from Ridenhower Shale of the Paint Creek Formation at Floraville, St. Clair County, Illinois, USA (Rexroad, Reference Rexroad1957, pl. 3, fig. 15).
Occurrence
Lower Shangsi Formation in the Yashui-A section; mid-Visean.
Description
The paired dextral and sinistral Pa elements are mirrored in general, except for locations of the lateral nodes on the blade. The free blade occupies less than half of the whole element in length, with ~15 denticles of varying size inclined ventrally. The basal cavity is longitudinally elongated and extends to the dorsal end. There is a constriction at the ventral one-third, which separates the platform into two parts. The ventral part is slightly less expanded in the smaller specimen but more in the larger specimens (Fig. 5.26, 5.29). Between four and seven denticles are on the carina, with the highest ones upright at the middle length and inclined dorsally. The dorsal part occupies two-thirds of the platform and with around seven to 10 denticles, slightly inclined dorsally. The denticles on the blade and ventral carina are laterally compressed in cross section with a flatter outline on the “left” side and usually fused or with germ denticles inserted between larger denticles (Fig. 5.31). The denticles on the dorsal carina are round in cross section, narrowly spaced ventrally, and widely spaced dorsally. In both dextral and sinistral Pa elements, small nodes occur on the “left” side of the blade above the lateral ridge, more ventrally inclined than the blade denticles (Fig. 5.33). The microtexture of coarse fibrous crystals is exhibited around the denticles but is not apparent on the small lateral nodes (Fig. 5.32, 5.33). In lateral view, the lower outline of the basal cavity is flat.
Materials
Four Pa elements (201233–201236)
Remarks
Vogelgnathus campbelli is cosmopolitan and has been reported from many regions of middle and low latitudes (e.g., North America: Rexroad, Reference Rexroad1957; Globensky, Reference Globensky1967; Purnell and von Bitter, Reference Purnell and von Bitter1992a; western Europe: Rhodes et al., Reference Rhodes, Austin and Druce1969; Park, Reference Park1983; van den Boogaard, Reference van den Boogaard1992; Purnell and von Bitter, Reference Purnell and von Bitter1992a; Nemyrovska and Samankassou, Reference Nemyrovska and Samankassou2005; eastern Europe: Kabanov et al., Reference Kabanov, Gibshman, Barskov, Alekseev, Goreva, Alekseev and Goreva2009; East Asia: Igo, Reference Igo1973; Qi et al., Reference Qi, Nemyrovska, Wang, Chen, Wang, Lane, Richards, Hu and Wang2014; north coast of Gondwana: Jenkins et al., Reference Jenkins, Crane and Mory1993; Nemyrovska et al., Reference Nemyrovska, Perret-Mirouse and Weyant2006; Medina-Varea, Reference Medina-Varea2018; Atakul-Ozdemir et al., Reference Atakul-Ozdemir, Altiner and Ozkan-Altiner2019; south Urals of central Asia: unpublished materials of the authors). We accept the recently revised phylogeny (Purnell and von Bitter, Reference Purnell and von Bitter1992a; Nemyrovska and Samankassou, Reference Nemyrovska and Samankassou2005; Sanz-Lopez et al., Reference Sanz-Lopez, Blanco-Ferrera and Miller2019), and the profiles of our specimens are more like the possible ancestor V. simplicatus (Rhodes, Austin, and Druce, Reference Rhodes, Austin and Druce1969) rather than the possible descendant V. postcampbelli (Austin and Husri, Reference Austin and Husri1974). Our specimens closely resemble the holotype and other specimens from the type area in Illinois and also in Atlantic Canada, possessing distinct lateral nodes (Norby and Rexroad, Reference Norby and Rexroad1985; Purnell and von Bitter, Reference Purnell and von Bitter1992a, Reference Purnell and von Bitterb). Almost all previous records of V. campbelli outside of North America do not have such characters, except for some specimens from Morocco (Medina-Varea, Reference Medina-Varea2018). Purnell and von Bitter (Reference Purnell and von Bitter1992b) studied these unusual asymmetrical-placed nodes and thought they were functional. The paring pattern for ozarkodinid Pa elements might be that the sinistral lies behind the dextral element (Purnell, Reference Purnell1995). More evidence is still needed to reveal why only some specimens of this species have such lateral nodes, excluding the interference of age and sedimentary setting. The asymmetry of the denticles in cross section is also found in many other ozarkodinids without lateral nodes (especially for the blade denticles). As the microtexture is well developed on both sides of the denticles, it is difficult to determine whether it is functional or just a wear structure.
Stratigraphic correlations
Conodonts are most abundant in the middle (bed 63; ~34.5 m) and upper (bed 104; ~63.5 m) parts of the Yashui-A section, including Pa elements of Cavusgnathus aff. Cav. unicornis, Cavusgnathus unicornis, Clydagnathus windsorensis, Hindeodus cristulus (Youngquist and Miller, Reference Youngquist and Miller1949), and Vogelgnathus campbelli. Two assemblages, described in the following, are recognized: the Cavusgnathus aff. Cav. unicornis–Cly. windsorensis assemblage from the upper Jiusi Formation and the Cav. unicornis assemblage from the lowermost Shangsi Formation (Figs. 3, 7).
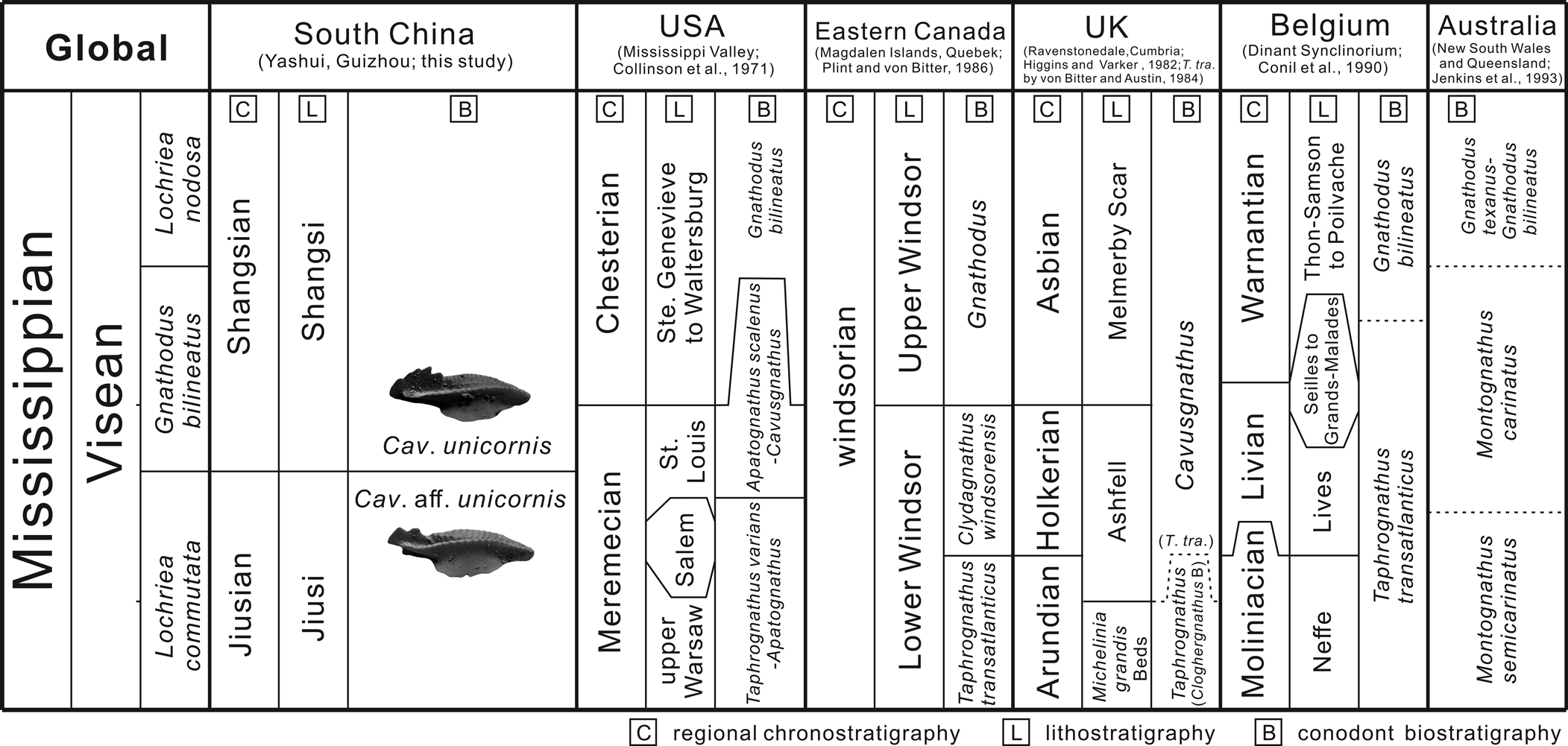
Figure 7. Correlation of the biozonations based on the shallow-water conodonts during the Visean of different regions.
In the Yashui-A section, common Visean conodont representatives of deep-water facies (e.g., Gnathodus and Lochriea) are absent. The Cavusgnathus aff. Cav. unicornis assemblage contains the nominate species through the lower part of the studied section, but its real first occurrence is not clear. Cavusgnathus aff. Cav. unicornis occurs with Clydagnathus windsorensis or Hindeodus cristulus in the relatively productive samples of beds 43–69. The Cav. unicornis assemblage is present in the uppermost part of the section. It is marked by the phylogenetic first occurrence of Cav. unicornis and also includes Vogelgnathus campbelli. Hindeodus cristulus is in both assemblages.
The aforementioned foraminifers of the MFZ11 to MFZ12 foraminiferal zone suggest a possible early to middle Visean age for the studied interval at the Yashui-A section. As for the conodonts, Hindeodus cristulus and Vogelgnathus campbelli are common in the late Visean and early Serpukhovian, but their first occurrences are much lower, from late Tournaisian (Metcalfe, Reference Metcalfe1981; Zhuravlev, Reference Zhuravlev2017; Sanz-Lopez et al., Reference Sanz-Lopez, Blanco-Ferrera and Miller2019). Invasions of V. campbelli were considered indicators of rapid sea-level-rise events in the Cantabrian Mountains, Spain (van den Boogaard, Reference van den Boogaard1992; Nemyrovska and Samankassou, Reference Nemyrovska and Samankassou2005); however, their occurrence in shallow-water facies is sparse and therefore not useful to correlate sea-level change. The shallow-water conodonts are particularly prone to facies control; thus, correlation between restricted and endemic faunas is complicated (Austin and Davies, Reference Austin and Davies1984; Barrick et al., Reference Barrick, Alekseev, Blanco-Ferrera, Goreva, Hu, Lambert, Nemyrovska, Qi, Ritter and Sanz-Lopez2022). However, conodont zonations for shallow-water facies have been recognized globally, based mainly on the cavusgnathids (Taphrognathus, Clydagnathus, Cavusgnathus, and Adetognathus). At Yashui, the only cosmopolitan species with potential for regional or long-distance correlations is Cav. unicornis, which also marks the base of the regional Shangsian stage.
In the central United States, this widespread species first occurred from the middle St. Louis breccia at the base of the Apatognathus scalenus–Cavusgnathus Zone (Collinson et al., Reference Collinson, Scott and Rexroad1962, Reference Collinson, Rexroad and Thompson1971; ~base of Faunal Unit 8 scitulus–scalensus Zone of Lane and Brenckle, Reference Lane and Brenckle2005). Near the zonal boundary, there is a short interval with co-occurrence of Cav. unicornis and Taphrognathus varians Branson and Mehl, Reference Branson and Mehl1941 (including the “Taphrognathus–Cavusgnathus transitions” of Rexroad and Collinson, Reference Rexroad and Collinson1963). We correlate the Cavusgnathus aff. Cav. unicornis assemblage at Yashui with this interval and above the main hosting horizons for T. varians of the Warsaw and Salem formations (Fig. 7).
In eastern Canada, the shallow-water conodont succession incudes the T. transatlanticus (von Bitter and Austin, Reference von Bitter and Austin1984) (the generic assignment is suspect; see Purnell et al., Reference Purnell, von Bitter and Groessens2002), Cly. Windsorensis, and Gnathodus zones in the Windsor Group, in ascending order (Plint and von Bitter, Reference Plint and von Bitter1986). The Cav. unicornis associated with Cly. windsorensis often occurred in the Gnathodus Zone (e.g., von Bitter and Plint-Geberl, Reference von Bitter and Plint-Geberl1982). The illustrated Cav. unicornis from the lower horizons within the T. transatlanticus Zone are similar to Cavusgnathus aff. Cav. unicornis in this study with blade-parapet notches (e.g., von Bitter and Plint-Geberl, Reference von Bitter and Plint-Geberl1982, pl. 3, figs. 14, 17, 18). We propose that the Cavusgnathus aff. Cav. unicornis and Cav. unicornis assemblages may be correlative with the Cly. windsorensis Zone in Canada, although the original concept of this zone includes no Cavusgnathus (Fig. 7).
The shallow-water conodont succession in the United Kingdom is similar to that of eastern Canada. In Ravenstonedale of Cumbria, Higgins and Varker (Reference Higgins and Varker1982) erected the Taphrognathus Zone marked by the co-occurrence of T. varians and T. carinatus (Higgins and Varker, Reference Higgins and Varker1982) (originally as Cloghergnathus carinatus). The overlying Cloghergnathus Zone is marked by the extinction of T. carinatus. Above this is the Cavusgnathus Zone that is marked by the first occurrence of Cav. unicornis. The lower two zones are hard to differentiate with the proposed markers (Purnell et al., Reference Purnell, von Bitter and Groessens2002), and there was a conodont-barren interval within the Ashfell Sandstone between the top of Cloghergnathus Zone and the base of the Cavusgnathus Zone. This barren interval was later filled by the T. transatlanticus Zone recognized in the Avon Gorge (von Bitter and Austin, Reference von Bitter and Austin1984). This zone is correlative with the same-name zone in eastern Canada but different in that the nominated species co-occurred with the Cly. windsorensis and probably the real first occurrence of Cav. unicornis s.l. in the United Kingdom. We think their Cav. unicornis (e.g., von Bitter and Austin, Reference von Bitter and Austin1984, pl. 19, figs. 12–14, 16–28) resembles our Cavusgnathus aff. Cav. unicornis and use this in our correlation (Fig. 7).
The T. transatlanticus Zone of Belgium is not very clear, but as Conil et al. (Reference Conil, Groessens, Laloux, Poty and Tourneur1990) stated, “a few specimens of small taphrognathid-like conodonts have been found at the top of the Moliniacian as well as some cavusgnathids at the base of the Livian.” Some of the Australian Adetognathus from the early to middle Visean (e.g., Adetognathus taphrognathoides Jenkins in Jenkins et al., Reference Jenkins, Crane and Mory1993, fig. 10A–C, H, I) might be Taphrognathus, and this is considered in the proposed correlation here (Fig. 7).
Considering all the preceding factors, the studied interval at Yashui could be tentatively correlated with Visean conodont zones from other regions that include the first occurrence of Cavusgnathus unicornis s.l. (Fig. 7). This horizon consistently occurs above the widespread Taphrognathus varians assemblage in multiple regions (e.g., the United States and the United Kingdom). The primitive type Cavusgnathus aff. Cav. unicornis commonly co-occurs with Clydagnathus windsorensis and occasionally with the suspect T. transatlanticus. The Cavusgnathus aff. Cav. unicornis to Cav. unicornis lineage is now recognized only in this single section. More work studying the lineage of Cav. unicornis will test our proposed phylogeny and may significantly improve global correlations of Visean shallow-water deposits.
Conclusion
Conodonts from the typical Visean shallow-water platform deposits in southern Guizhou have been studied. Two conodont assemblages are described, one from the upper Jiusi Formation and the other from the lowest Shangsi Formation of the Yashui-A section. The conodont faunas of this interval are dominated by cavusgnathids, and the yield is low. The first occurrences of Pa elements of Cavusgnathus aff. Cav. unicornis, Hindeodus cristulus, Clydagnathus windsorensis, Cav. unicornis, and Vogelgnathus campbelli occur successively. The Cavusgnathus aff. Cav. unicornis assemblage and the Cav. unicornis assemblage are associated with the lithological boundary between the two formations. Judging from the intermediate features of the Pa and possible Pb elements, Cavusgnathus aff. Cav. unicornis is likely a direct ancestor of Cav. unicornis and a possible descendant of Cav. hudsoni. The first occurrence datum of Cav. unicornis near the base of the Shangsi Formation is constrained by the most abundant data here compared with neighboring regions and is suggested to mark the beginning of the middle Visean and regional Shangsian. Although the underlying zone with Taphrognathus varians has not been reported from South China, this work shows potential in finding biostratigraphically significant conodonts in other shallow-water sections. More work is needed in this region to refine the temporal scheme.
Acknowledgments
We greatly acknowledge X.H. Wu (Guiyang) for the assists in the fieldwork and D. Korn (Berlin) for his help in language polishing. We thank N. Hogancamp (Houston), S.A. Leslie (Harrisonburg), and S. Zamora (editor) for their constructive advice. This work was financially supported by the National Natural Science Foundation of China (41902030, 41630101) and the Strategic Priority Research Programs of Chinese Academy of Sciences (XDB26000000).
Declaration of competing interests
The authors declare none.







