


Introduction
In temperate shrublands and grasslands extensive livestock ranching affects wild herbivore populations through interspecific competition for forage and, at the same time, overgrazing can result in increased desertification (Montgomery, Reference Montgomery2007; Brown, Reference Brown2008). As domestic livestock ranching practices in South America were mostly imported from Europe native species are not seen as a resource to improve local economies. Instead, wild mammals are perceived as an obstacle to the development of domestic livestock production (du Toit, 2010) and hence livestock ranching leads to the decline and local extirpation of wild herbivores and carnivores (Mishra et al., Reference Mishra, Allen, McCarthy, Madhusudan, Bayarjargal and Prins2003).
In arid Patagonia, Argentina, the guanaco Lama guanicoe has been the only large native mammalian herbivore since the end of the Pleistocene. The total population was 7–10 million at the end of the 19th century (Franklin, Reference Franklin, Mares and Genoways1982) when European settlers introduced the domestic sheep Ovis aries. Since then sheep ranching has been the dominant economic activity and the abundance and distribution of the guanaco has declined drastically. Sheep numbers peaked at 22 million within 50 years of their introduction (Soriano & Movia, Reference Soriano and Movia1986), while guanacos declined to < 1 million by the end of the 20th century (Baldi et al., Reference Baldi, Novaro, Funes, Walker, Ferrando, Failla, Carmanchahi, du Toit, Kock and Deutsch2010) and now occupy only 40% of their original range (Puig, Reference Puig and Torres1992; Franklin et al., Reference Franklin, Bas, Bonacic, Cunazza and Soto1997). The abundance of the guanaco is negatively associated with both sheep abundance and the availability of preferred forage plants (Baldi, Reference Baldi1999; Baldi et al., Reference Baldi, Albon and Elston2001). In addition, uncontrolled hunting, poaching and chasing by landowners who consider the guanaco a threat to sheep production are still widespread across arid Patagonia (Puig, Reference Puig and Torres1992; Cunazza et al., Reference Cunazza, Puig, Villalba and Puig1995; Baldi et al., Reference Baldi, Novaro, Funes, Walker, Ferrando, Failla, Carmanchahi, du Toit, Kock and Deutsch2010). Interspecific competition with sheep also occurs and, as a consequence, guanacos often occupy marginal areas of lower productivity (Baldi et al., Reference Baldi, Albon and Elston2001). Evidence of habitat degradation across Patagonian rangelands because of overgrazing by sheep comprises changes in vegetation structure and composition and subsequent intensification of soil erosion (Adler et al., Reference Adler, Milchunas, Sala, Burke and Lauenroth2005; Chartier & Rostagno, Reference Chartier and Rostagno2006; Bisigato et al., Reference Bisigato, Laphitz and Carrera2008). The impact of sheep grazing on vegetation is greater than that of native herbivores such as guanacos, which have co-evolved with the native plant communities (Lauenroth, Reference Lauenroth1998), and severe desertification has now affected nearly 30% of the 700,000 km2 Patagonian steppe (del Valle, Reference del Valle1998).
Thus the introduction of sheep, the rapid development of the ranching industry (Baldi et al., Reference Baldi, Novaro, Funes, Walker, Ferrando, Failla, Carmanchahi, du Toit, Kock and Deutsch2010), uncontrolled commercial hunting (Ojeda & Mares, Reference Ojeda, Mares, Mares and Genoways1982) and poaching were the main causes of the drastic decline of the guanaco in Patagonia. As sheep ranching is still a major activity in this region and effective protected areas encompass only 1% of the land, the opportunities for guanacos to establish large, functional populations are scarce.
Península Valdés is a 4,000 km2 protected area in eastern Patagonia (Fig. 1), declared a UNESCO Natural World Heritage Site in 1999. Although sheep ranching is still widespread some private landowners initiated ecotourism management as a complementary activity to wool production. In addition, in 2005, the local NGO Fundación Vida Silvestre Argentina purchased a 7,360 ha ranch formerly dedicated to sheep production, for conversion to a private wildlife reserve; all the c. 3,500 sheep were removed and a permanent warden appointed.

Fig. 1 (a, b) Location of Reserva San Pablo de Valdés in Península Valdés, Argentine Patagonia. (c) Distribution of transects inside and outside the Reserve, sampling sites in each vegetation community inside and outside the Reserve, and location of water points.
The drastic changes in management at the new Reserva San Pablo de Valdés resulted in a unique opportunity to study both the response of guanacos and the vegetation following the removal of the main source of disturbance. Our objectives in the study reported here were to examine any changes in guanaco abundance at the Reserve following removal of the sheep, to survey guanaco distribution among the vegetation communities within the Reserve as an indicator of habitat preferences in the absence of sheep, and to compare vegetation cover and composition between the Reserve and neighbouring sites where sheep ranching continues.
Study area
Reserva San Pablo de Valdés is located in Península Valdés, Chubut, Argentine Patagonia (Fig. 1). The most distinctive climatic factor across this semi-arid peninsula is the low average annual rainfall (280 mm), which falls mostly in the autumn and winter. Four vegetation communities in the Reserve have been described (Codesido et al., Reference Codesido, Beeskow, Blanco and Johnson2005): (1) Shrub steppe dominated by Chuquiraga avellanedae, Lycium chilense, Schinus johnstonii, Menodora robusta, Acantholippia seriphioides, Ephedra ochreata and Prosopis denudans. (2) Grass steppe dominated by Sporobolus rigens and Nassella tenuis, with patches of Baccharis divaricata. (3) Shrub–grass steppe dominated by Chuquiraga erinacea var. hystrix, Chuquiraga avellanedae, A. seriphioides, with N. tenuis, S. rigens, Piptochaetium napostaense and Pappostipa speciosa the dominant grass species. (4) Dwarf-shrub steppe dominated by Hyalis argentea, isolated shrubs of Suaeda divaricata, C. avellanedae and Lycium spp., and patches dominated by Cyclolepis genistoides or Baccharis divaricata. Sheep density was 47 km-2 in 2005 before the ranch was converted to a Reserve. On the neighbouring ranches mean sheep density per paddock was 52 km-2 during 2006–2008 (M. Nabte, pers. comm.).
Methods
We conducted line-transect surveys of guanacos both at Reserva San Pablo de Valdés and across several ranches outside the Reserve during January–March 2006 and November 2008, along dirt roads and tracks (Fig. 1). Total transect length per survey was 22 and 71 km, inside and outside the Reserve, respectively. Surveys were conducted from an open pick-up vehicle with two observers standing in the back, using the distance sampling method (Burnham et al., Reference Burnham, Anderson and Laake1980, 1993). For every guanaco group encountered we stopped the vehicle, recorded the number of animals, and the perpendicular distance (measured using a laser rangefinder) from the transect line to the location where the group was standing at the time it was detected.
We estimated guanaco densities using Distance v. 5.0 (Buckland et al., Reference Buckland, Anderson, Burnham and Laake1993). We also calculated encounter rate of individuals (guanacos per km travelled) per transect, to compare trends in guanaco abundance inside and outside the Reserve. We applied a linear model to account for the effects of site (inside vs outside the Reserve) and year (2006 vs 2008) on the log of encounter rate. We compared encounter rates instead of densities because reliable estimates of density derived from distance sampling require a minimum number of observations, and guanaco densities outside the Reserve were so low that the detection function would have had to be pooled across sites to achieve the minimum sample size required. Density estimates inside and outside the Reserve are not therefore independent, precluding statistical comparison.
Three additional surveys were conducted in April and November 2008 and January 2009 to investigate differences in the distribution of guanacos across vegetation communities in Reserva San Pablo de Valdés. We travelled along all roads available inside the Reserve, recording the location and size of each guanaco group observed. We calculated group encounter rates (groups per km travelled) in each vegetation community and compared log of group encounter rate among vegetation communities using a linear model.
To account for possible group size-effects associated with different vegetation communities or season (southern autumn, spring and summer), we fitted a mixed linear model to the log of group size (Pinheiro & Bates, Reference Pinheiro and Bates2000) using vegetation community as fixed and season as a random factor. We tested the statistical significance of differences in group size among vegetation communities using a Wald test (Payne et al., Reference Payne, Murray, Harding, Baird, Soutar and Lane2003). GenStat v. 7.2 (Lawes Agricultural Trust–VSN International, Rothamstead, UK) was used for all statistical analyses.
During 2009 we conducted plant surveys across the four vegetation communities, both in Reserva San Pablo de Valdés and on contiguous ranches outside the Reserve. Inside the Reserve one site was surveyed within each vegetation community and outside the Reserve two sites per vegetation community (Fig. 1). At each sampling site we randomly placed two linear 50-m transects, along which we estimated total plant cover, cover per functional group and floristic composition using the point interception line method at 1-m intervals (Herrick et al., Reference Herrick, Van Zee, Havstad, Burkett and Whitford2005). We compared the number of interceptions of the total canopy and per functional group among sampling sites (inside and outside the Reserve) within each vegetation community using a χ 2 test. We compared floristic composition and abundance of plant species among sampling sites within each vegetation community using rank-abundance curves (Feinsinger, Reference Feinsinger2003). We applied this latter analysis only to grass species because it is the functional group preferred by herbivores.
Results
The density of guanacos was higher in Reserva San Pablo de Valdés than on the surrounding ranches, and increased from a mean of 3.95 ± SE 1.14 in 2006 to 12.95 ± SE 4.14 km-2 in 2008, whereas it remained relatively constant across the ranches surrounding the Reserve (mean 1.10 ± SE 0.53 km-2 in 2006 and 1.29 ± SE 0.46 in 2008).
We assessed year and site effects on the encounter rate of individuals and found that these factors explained almost 80% of the observed variation in the data (R2 = 0.798, df = 28, P < 0.001). Encounter rate was significantly higher inside the Reserve than outside, irrespective of the year, and significantly higher in 2008 than in 2006 inside the Reserve (Table 1), in accordance with the three-fold increase in guanaco density.
Table 1 Analysis of the effects of year and site on the log of encounter rate of guanacos Lama guanicoe in Reserva San Pablo de Valdés and neighbouring sites, with the estimated effect and the statistical significance of year, site and year*site interaction. The estimated effects are expressed as differences compared with reference levels, which are 2006, Outside Reserve.

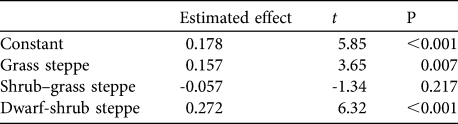
Although mean group size of guanacos was similar among vegetation communities (Wald = 6.61, df = 3, P = 0.085) the group encounter rates varied significantly (R 2 = 0.86, df = 8, P < 0.001). Encounter rates of groups were higher in grass steppe and dwarf-shrub steppe communities (1.7–2.3 groups km-1) than in shrub steppe and shrub–grass steppe communities (0.8–1 groups km-1; Table 2).
Table 2 Analysis of the effects of vegetation communities on the log of encounter rate of guanacos, with the estimated effect of the different vegetation communities. The estimated effects are expressed as differences compared with a reference level, which is the encounter rate in the shrub steppe vegetation community.
Total plant cover tended to be higher inside than outside the Reserve in all four vegetation communities (Fig. 2). Significant effects were found only for the dwarf-shrub and grass steppe communities (χ 2 = 10.53, df = 2, P < 0.01; χ 2 = 74.25, df = 2, P < 0.001, respectively). These differences were accounted for by the cover of perennial grasses in both communities inside the Reserve (χ 2 = 13.33, df = 2, P < 0.01; χ 2 = 80.02, df = 2, P < 0.001, for dwarf-shrub and grass steppe, respectively). For the shrub–grass steppe community, although grass cover was higher inside than outside the Reserve (χ 2 = 20.17, df = 2, P < 0.001), the cover of woody plants was significantly lower inside the Reserve than in one of the sampling sites outside the Reserve (χ 2 = 29.15, df = 2, P < 0.001) resulting in similar total plant cover.

Fig. 2 Number of intercepts of the total canopy, shrubs/dwarf shrubs and grasses in two 50-m transects (at 1-m point intervals) in the sampling site inside the Reserve and in the two sampling sites outside the Reserve, in each of the four vegetation communities. Asterisks indicate significant differences among sites within each community (* P < 0.05, ** P < 0.001 and *** P < 0.0001).
In all four vegetation communities the richness of perennial grass species was higher inside the Reserve than outside (Fig. 3). However, for all four communities the two most abundant grass species (representing > 70% of total grass cover) were the same both inside and outside the Reserve. Outside the Reserve the differences in relative abundances among the dominant and non-dominant grasses were higher than inside the Reserve.

Fig. 3 Rank-abundance curves for grass species in the four vegetation communities, with log of the proportion of each grass species (log pi) in the sampling site inside the Reserve and in the two sampling sites outside the Reserve. Nt, Nassella tenuis; Pli, Poa ligularis; Pla, Poa lanuginosa; Ps, Pappostipa speciosa; Pn, Piptochaetum napostaense; Ely, Elymus sp.; Sr, Sporobolus rigens; Pa, Panicum sp..
Discussion
Guanaco abundance
The differing densities of guanacos and sheep between sites and spatial shifts in habitat use support predictions derived from interspecific competition hypothesis (de Boer & Prins, Reference de Boer and Prins1990). Previous studies have shown that guanacos and sheep compete for forage because they eat similar proportions of the same plant species, and that guanaco abundance is negatively correlated with sheep abundance and with the abundance of preferred forage plants (Baldi et al., Reference Baldi, Albon and Elston2001, Reference Baldi, Pelliza-Sbriller, Elston and Albon2004) Accordingly, sheep removal at Reserva San Pablo de Valdés resulted in a habitat free of competitors that was rapidly occupied by guanacos. Habitat shifts by guanacos have been previously reported for the arid lands of central Chubut (Baldi et al., Reference Baldi, Albon and Elston2001). Similar habitat shifts of wild ungulate species following changes in the density of domestic livestock have been reported for other wild ungulate species, such as the impala Aepyceros melampus (Fritz et al., Reference Fritz, De Garine-Wichatitsky and Letessier1996) and the mule deer Odocoileus hemionus (Loft et al., Reference Loft, Menke and Kie1991).
In addition to the removal of sheep, changes in management at Reserva San Pablo de Valdés included the presence of a permanent warden who prevented poaching or chasing of the guanacos. Elsewhere, guanacos are either killed or chased because they are perceived as an impediment to sheep production (Baldi et al., Reference Baldi, Novaro, Funes, Walker, Ferrando, Failla, Carmanchahi, du Toit, Kock and Deutsch2010). Although the entire Península Valdés is a protected area and the presence of Government officers has increased since it was declared a World Heritage Site, the limited number of personnel and the persistence of large sheep stocks are probably still limiting the recovery of guanacos and their habitat.
Guanaco distribution
Guanaco groups inside Reserva San Pablo de Valdés were mostly observed in grass-dominated communities, which have a higher total plant cover than shrub-dominated areas. Although guanacos are generalist herbivores, grass species make up most of their diet in Chubut (Baldi et al., Reference Baldi, Pelliza-Sbriller, Elston and Albon2004). Ongoing studies on guanaco diet in the Reserve indicate that 70% comprises the grasses N. tenuis and Poa spp. (A. Marino, unpubl. data). Thus, guanacos in Reserva San Pablo de Valdés are selecting vegetation communities with a high foraging value, in contrast to most other sites with sheep in which guanacos occupy marginal habitats. Baldi et al. (Reference Baldi, Albon and Elston2001) found that guanaco densities were negatively related to the abundance of preferred plant species (Nassella and Poa) in Chubut, including Península Valdés, where sheep were present.
Vegetation cover and composition
Grass and total plant cover, and grass species richness and evenness inside Reserva San Pablo de Valdés were higher than at surrounding neighbouring sites. Although overgrazing can result in severe desertification (Adler et al., Reference Adler, Milchunas, Sala, Burke and Lauenroth2005; Chartier & Rostagno, Reference Chartier and Rostagno2006) it appears that vegetation inside the Reserve recovered to some extent after removal of sheep even though guanaco abundance increased by three-fold. The estimate of c. 13 guanacos km-1 in the Reserve is among the highest densities reported for the species in Patagonia (Baldi et al., Reference Baldi, Novaro, Funes, Walker, Ferrando, Failla, Carmanchahi, du Toit, Kock and Deutsch2010). In terms of sheep-equivalent biomass one adult guanaco is approximately equal to two adult sheep (Elissalde et al., Reference Elissalde, Escobar and Nakamatsu2002). Therefore the net stocking rate at Reserva San Pablo de Valdés decreased from 47 sheep km-1 in 2005 to 26 sheep-equivalents km-1 in 2008. Patagonia has a long history of grazing by guanacos and this is evident in the anti-herbivore defences of various plant species (Lauenroth, Reference Lauenroth1998), whereas sheep grazing in the region is more recent and drives vegetation deterioration by reducing both total plant cover and the availability of highly palatable, perennial grasses, and decreasing plant species richness (Beeskow et al., Reference Beeskow, Elissalde and Rostagno1995; Bisigato & Bertiller, Reference Bisigato and Bertiller1997).
Guanacos recycle nitrogen efficiently (Schmidt-Nielsen et al., Reference Schmidt-Nielsen, Schmidt-Nielsen, Houpt and Jarnum1957; Livingston et al., Reference Livingston, Payne and Friend1962) and are larger than sheep and thus they are expected to tolerate lower-quality forage (Demment & Van Soest, Reference Demment and Van Soest1985; Illius & Gordon, Reference Illius and Gordon1991; Fraser & Gordon, Reference Fraser and Gordon1997). Trampling by guanacos is less damaging to soil and vegetation than sheep because guanacos posses nail-covered digital pads rather than hooves (Wheeler, Reference Wheeler1995). Finally, guanacos are not confined to paddocks, as are sheep, and move across different foraging areas, thus allowing vegetation to recover (Raedeke, Reference Raedeke1979). It is likely, therefore, that guanacos have a lower impact on vegetation compared to the known effects of sheep. However, as vegetation recovery also depends on previous vegetation condition, grazing history and primary production (Soriano et al., Reference Soriano, Sala and León1980), vegetation recovery at Reserva San Pablo de Valdés and other sites requires monitoring.
Conclusions and conservation implications
Our results show that a drastic change in land management and the appointment of a warden resulted in a rapid response by guanacos: their abundance increased three-fold within Reserva San Pablo de Valdés and their density was an order of magnitude greater than that in neighbouring sites. It also appears that the vegetation recovered to some extent after the removal of sheep despite increased grazing pressure by guanacos. A monitoring programme inside the Reserve has now been developed and the most recent survey conducted, in 2010, showed that the density of guanacos is still increasing inside the Reserve and that vegetation is recovering, with an increase in total plant and grass cover. However, the 7,360 ha Reserve is too small to harbour a population of large herbivores such as guanacos, which need to be managed across Península Valdés as a whole to avoid land-bridge island effects (Newmark, Reference Newmark1995).
An ongoing review of the management plan for Península Valdés, developed in 2000, is showing that most of the objectives have not been attained because of the lack of implementation of appropriate actions. At the same time, pressure by landowners on wildlife authorities to reduce guanaco densities in particular areas of the peninsula has increased since 2010 because of an ongoing drought in the area. As Península Valdés is a World Heritage Site in IUCN category VI (IUCN 1994), implementation of the management plan is required, including programmes for the coexistence of sustainable livestock production and healthy wildlife populations. We believe the plan should consider an array of strict reserves, such as Reserva San Pablo de Valdés, in a matrix of sustainable use in which the guanaco could contribute to improving the local economy as an iconic species of Patagonia.
Acknowledgements
This study was funded and supported by Consejo Nacional de Investigaciones Científicas y Técnicas, the Ministerio de Educación de la Provincia de Chubut, the Rufford Small Grant Foundation for Nature Conservation, the Wildlife Conservation Society, and Fundación Vida Silvestre Argentina. We thank A. Johnson, M. Nabte, F. Siri, M. Primost, V. Alonso Roldán, M. Zamero and Lucas Andreani for help during the fieldwork.
Biographical sketches
María Virginia Burgi is interested in the study of the ecological and anthropogenic factors affecting the distribution of the guanaco in Patagonia. Andrea Marino focuses on the behavioural ecology and population dynamics of ungulates. María Victoria Rodríguez investigates plant interactions and water use by plant functional groups in grazed ecosystems. Gustavo Pazos studies spatial vegetation dynamics in arid ecosystems and the effect of grazing disturbance on the population ecology of grasses. Ricardo Baldi’s research focuses on the ecological and anthropogenic processes affecting guanacos and other wildlife species in Patagonia.





