



Introduction
According to data published by the World Health Organization, infertility is a condition that affects ∼48 million couples and 186 million individuals worldwide (WHO, 2020). Female infertility is responsible for approximately half of the cases, with ovarian ageing being a current societal dilemma (Walker and Tobler, Reference Walker and Tobler2022).
Oocyte quality is a critical component in overall fertility, as oocyte competence is critical for normal fertilization, embryonic development, implantation and full pregnancy (Conti and Franciosi, Reference Conti and Franciosi2018; Sirait et al., Reference Sirait, Wiweko, Jusuf, Iftitah and Muharam2021). Oocyte competence is acquired during oogenesis, where the oocyte passes through a sequence of maturation changes involving both nuclear and cytoplasmic domains (Arroyo et al., Reference Arroyo, Kim and Yeh2020). Nuclear maturation includes a cascade of events culminated by the pre-ovulatory peak of luteinizing hormone (LH), which allows the oocyte to resume meiosis and ovulate (Sánchez and Smitz, Reference Sánchez and Smitz2012). This process is characterized by nuclear envelope breakdown (GVBD: germinal vesicle breakdown) in prophase I oocytes (GV oocytes), followed by the condensation and alignment of chromosomes in a metaphase plate-I (MI oocytes). MI oocytes then enter the first meiotic division, with extrusion of the first polar body (PB1), containing half of the chromosomes, and condensation and alignment of the other half of the chromosomes in a metaphase plate-II (MII oocytes), with oocytes remaining in an MII arrest until fertilization (Arroyo et al., Reference Arroyo, Kim and Yeh2020). Nuclear maturation must occur in coordination with cytoplasmic maturation (Eppig, Reference Eppig1996). Cytoplasmic maturation is characterized by the accumulation of mRNA, proteins, substrates and nutrients (Watson, Reference Watson2007; Richani and Gilchrist, Reference Richani and Gilchrist2018; Cornet-Bartolomé et al., Reference Cornet-Bartolomé, Barragán, Zambelli, Ferrer-Vaquer, Tiscornia, Balcells, Rodriguez, Grinberg and Vassena2021), as well as by the remodelling of organelles and the cytoskeleton (Mao et al., Reference Mao, Lou, Lou, Wang and Jin2014; Coticchio et al., Reference Coticchio, Dal Canto, Mignini Renzini, Guglielmo, Brambillasca, Turchi, Novara and Fadini2015: Reader et al., Reference Reader, Stanton and Juengel2017), in an intimate cross-talk with the surrounding follicular cells (Dumesic et al., Reference Dumesic, Meldrum, Katz-Jaffe, Krisher and Schoolcraft2015; May-Panloup et al., Reference May-Panloup, Boucret, Chao de la Barca, Desquiret-Dumas, Ferré-L’Hotellier, Morinière, Descamps, Procaccio and Reynier2016).
Although the objective in assisted reproductive treatments (ART), using controlled hormonal ovarian hyperstimulation, is to obtain a maximum number of MII oocytes, in ∼10–15% of the cycles immature oocytes, GV and MI, are also retrieved (Ebner et al., Reference Ebner, Moser, Sommergruber and Tews2003; Pinto et al., Reference Pinto, Oliveira, Cardoso, Teixeira-da-Silva, Silva, Sousa and Barros2009; Braga et al., Reference Braga, Zanetti, Setti, Iaconelli and Borges2020), and, in a lower percentage, oocytes may display cytoplasmic and extracytoplasmic dimorphisms (Rienzi et al., Reference Rienzi, Vajta and Ubaldi2011; Sá et al., Reference Sá, Cunha, Silva, Luís, Oliveira, Teixeira da Silva, Barros and Sousa2011; Braga et al., Reference Braga, Setti, Figueira, Machado, Iaconelli and Borges2013, Reference Braga, Zanetti, Setti, Iaconelli and Borges2020; Sousa et al., Reference Sousa, Teixeira da Silva, Silva, Cunha, Viana, Oliveira, Sá, Soares, Oliveira and Barros2015, Reference Sousa, Cunha, Silva, Oliveira, Pinho, Almeida, Sá, da Silva, Oliveira and Barros2016a; Stigliani et al., Reference Stigliani, Moretti, Casciano, Canepa, Remorgida, Anserini and Scaruffi2018), due to maturation defects (Windt et al., Reference Windt, Coetzee, Kruger, Marino, Kitshoff and Sousa2001; Tripathi et al., Reference Tripathi, Kumar and Chaube2010; Conti and Franciosi, Reference Conti and Franciosi2018; Ozturk, Reference Ozturk2020). Some immature oocytes may resume meiosis during in vitro culture, but show reduced developmental competence (Ebner et al., Reference Ebner, Moser, Sommergruber and Tews2003, 2006; Shu et al., Reference Shu, Gebhardt, Watt, Lyon, Dasig and Behr2007; Azevedo et al., Reference Azevedo, Pinho, Silva, Sá, Thorsteinsdóttir, Barros and Sousa2014; De Vos et al., Reference De Vos, Grynberg, Ho, Yuan, Albertini and Gilchrist2021; Karavani et al., Reference Karavani, Wasserzug-Pash, Mordechai-Daniel, Bauman, Klutstein and Imbar2021).
Oocyte competence can be evaluated through morphologic and molecular analyses. The ultrastructural morphology of human oocytes at different maturation stages was evaluated in oocytes obtained from ART cycles (Baca and Zamboni, Reference Baca and Zamboni1967; Zamboni et al., Reference Zamboni, Thompson and Smith1972; Sathananthan and Trounson, Reference Sathananthan and Trounson1982, Reference Sathananthan and Trounson2000; Sathananthan, Reference Sathananthan1985, Reference Sathananthan1994, Reference Sathananthan1997; Reference Sathananthan, Trounson and Gosden2003; Reference Sathananthan2013; Sundström et al., Reference Sundström, Nilsson, Liedholm and Larsson1985; Motta et al., Reference Motta, Nottola, Micara and Familiari1988, Reference Motta, Nottola, Makabe, Heyn and Jansen2000; Familiari et al., Reference Familiari, Heyn, Relucenti, Nottola and Sathananthan2006; Sathananthan et al., Reference Sathananthan, Selvaraj, Girijashankar, Ganesh, Selvaraj and Trounson2006; Morimoto, Reference Morimoto2009; Nottola et al., Reference Nottola, Macchiarelli and Familiari2014). Qualitative morphological studies have also been recently conducted in donor oocytes (Trebichalská et al., Reference Trebichalská, Kyjovská, Kloudová, Otevřel, Hampl and Holubcová2021). Molecular research involved the analysis of oocyte and cumulus cells transcriptome (Cillo et al., Reference Cillo, Brevini, Antonini, Paffoni, Ragni and Gandolfi2007; Gasca et al., Reference Gasca, Pellestor, Assou, Loup, Anahory, Dechaud, De Vos and Hamamah2007; Wells and Patrizio, Reference Wells and Patrizio2008; Ouandaogo et al., Reference Ouandaogo, Frydman, Hesters, Assou, Haouzi, Dechaud, Frydman and Hamamah2012; Coticchio et al., Reference Coticchio, Ophir, Yung, Baum, Dal Canto, Mignini-Renzini, Brambillasca, Fadini and Hourvitz2017), proteome (Virant-Klun et al., Reference Virant-Klun, Leicht, Hughes and Krijgsveld2016; Shen et al., Reference Shen, Liu, Zhu, Zhang, Wang, Wang, Wang, Liu, Li and Liu2017) and metabolome (Venturas et al., Reference Venturas, Yang, Kumar, Wells, Racowsky and Needleman2021), or zona pellucida gene mRNA expression (Canosa et al., Reference Canosa, Adriaenssens, Coucke, Dalmasso, Revelli, Benedetto and Smitz2017), with development of oocyte quality markers (Sirait et al., Reference Sirait, Wiweko, Jusuf, Iftitah and Muharam2021).
Despite the importance of descriptive morphological studies, the qualitative evaluation of oocyte morphological characteristics can be incomplete or erroneous. As knowledge of the real morphology of the human oocyte is critical for the clinical development of new methods of cryopreservation and in vitro maturation, Pires-Luís et al. (Reference Pires-Luís, Rocha, Bartosch, Oliveira, Silva, Barros, Sá and Sousa2016) and Coelho et al. (Reference Coelho, Pires-Luís, Oliveira, Alves, Leal, Cunha, Barreiro, Barros, Sá and Sousa2020) conducted pioneer quantitative studies with a stereological approach in GV and MI oocytes, respectively. In these studies, the relative volumes of the different organelles were measured and the distribution of organelles in the different regions of the oocyte, cortex, subcortex and inner cytoplasm (IC), were evaluated.
At this time, the use of donor oocytes showed an exponential increase due to the large improvements in the cryopreservation technique and luteal phase endometrium preparation (Walker and Tobler, Reference Walker and Tobler2022). Although with the use of donor oocytes live birth rates are high, there is still a place for implantation failures. This has been attributable to recipient uterine conditions or partner male factors [European IVF-Monitoring Consortium (EIM) for the European Society of Human Reproduction and Embryology (ESHRE), 2021; Sciorio et al., Reference Sciorio, Antonini and Engl2021; Williams et al., Reference Williams, Ellis, Wilkinson, Kramer, Datta and Guzick2022], with the competence of donor oocytes being admitted to be optimized.
To our knowledge, this ultrastructural study is the first for donor MII oocytes using a stereological approach, in which we intended to quantify the spatial distribution of organelles. Surprisingly, we found a set of donor oocytes showing nuclear maturity but cytoplasmic immaturity. The results of the present report therefore provide evidence that even donor oocytes can show impaired competence.
Materials and methods
Ethical approval
Ethical guidelines were followed when conducting the research. According to the National Law on Medically Assisted Procreation (Law 32/2006) and the National Council for Medically Assisted Procreation guidelines (CNPMA, 2018), the use of clinical databases and surplus gametes for research may be used under strict individual anonymity and after patient written informed consent. Surplus donor oocytes were used after patients signed an informed consent agreeing to share their own gamete samples for the present study. The present laboratory experiments were executed under the Joint Ethics Committee of the Hospital and University, CHUP/ICBAS approval number 2019/CE/P017 (266/CETI/ ICBAS). This work did not involve human or animal experiments and therefore the provisions of the Declaration of Helsinki as revised in Tokyo 2004 do not apply.
Patients
This research was performed on five MII oocytes recovered from a 22-year-old oocyte donor after controlled ovarian stimulation during a donation programme performed at the Centro de Procriação Medicamente Assistida (CPMA), Centro Materno-Infantil do Norte (CMIN) Albino Aroso, Centro Hospitalar do Porto (CHUPorto), which is the National Gamete Bank.
Ovarian-controlled hyperstimulation
The oocyte donor underwent controlled ovarian hyperstimulation with a gonadotropin-releasing hormone antagonist protocol (GnRH; MSD, Hertfordshire, UK). Recombinant follicle-stimulating hormone (rFSH; Teva Biotech, Germany) was used for ovarian stimulation. A GnRH agonist (0.2 mg triptorelin; Ipsen Pharma Biotech, Signes, France) was administrated for triggering oocyte maturation, 36 h before follicular puncture (Huirne et al., Reference Huirne, Homburg and Lambalk2007; Pinto et al., Reference Pinto, Oliveira, Cardoso, Teixeira-da-Silva, Silva, Sousa and Barros2009).
Gamete and embryo handling
Procedures were performed on a K-Systems laminar flow chamber with a thermal base at 37°C (Cooper Surgical, Malöv, Denmark). Cumulus–oocyte complexes (COC) were collected in 1-well culture dishes (Falcon, Corning, New York, NY, USA) with SynVitro Flush medium (without heparin; Origio, Malöv, Denmark) and their handling and culture were performed under paraffin oil (Ovoil-100, Vitrolife, Frölunda, Sweden). After collection, COCs were washed and then cultured in Sequential Fert medium (Origio) in an Esco incubator (MRI-6A10, EscoMedical, Singapore, Singapore) (37°C, 5% O2, 6% CO2, 89% N2) for 2 h. After denudation with recombinant hyaluronidase (ICSI Cumulase, Origio), oocytes were transferred to 4-well culture dishes (Nunc, Thermo Scientific, Denmark) with Sequential Fert medium (Origio) and processed for transmission electron microscopy (TEM). The remaining donor oocytes were vitrified under the National Gamete Bank programme according to Kitazato methodology (Kitazato Corporation, Japan).
Transmission electron microscopy
Oocytes were fixed in Karnovsky solution (2.5% glutaraldehyde, 4% paraformaldehyde, 0.15 M sodium cacodylate buffer) (Sigma-Aldrich, St. Louis, USA; Merck, Darmstadt, Germany) at room temperature for 30 min, followed by 2 h at 4°C. After washing in 0.15 M sodium cacodylate buffer, pH 7.3 (Merck) for 2 h at 4°C, oocytes were post-fixed with 2% osmium tetroxide (Merck) in buffer containing 0.8% potassium ferricyanide (Merck) for 2 h at 4°C, washed in a buffer for 10 min, serially dehydrated in ethanol (Panreac, Barcelona, Spain), equilibrated with propylene oxide (Merck), and embedded in Epon (Sigma). Semithin and ultrathin sections were cut using diamond knives (Diatome, Hatfield, Switzerland) in an LKB ultramicrotome (Leica Microsystems, Weltzlar, Germany). A random number table was used to select the initial cut, and then oocytes were serially sectioned and sampled every 10 μm. Ultrathin sections were collected on 100 mesh formvar carbon-coated copper grids (Taab, UK) and stained with 3% aqueous uranyl acetate for 20 min (BDH, Poole, UK) and Reynolds lead citrate for 10 min (Merck) at room temperature in a light-protected environment. Ultrathin sections were observed on a JEOL 100CXII transmission electron microscope (JEOL, Tokyo, Japan) at 60 kV (Sousa and Tesarik, Reference Sousa and Tesarik1994; El-Shafie et al., Reference El-Shafie, Windt, Kitshoff, McGregor, Sousa, Wranz, Kruger, El-Shafie, Sousa and Kruger2000).
Stereological and statistical analysis
Systematic sampling was performed on each microscope grid, with photographs taken at alternate TEM field spaces when more than 50% of the field was occupied by the oocyte cytoplasm. Images were taken at ×5300 magnification and printed at 20.2 cm × 20.2 cm. A classical manual stereological technique based on point counting with a suitable stereological grid was used. The grid was placed over the printed photographs and the number of grid points placed over each organelle was noted. The relative volume (Vv) of each organelle was obtained by applying the formula Vv (organelle, oocyte) = [number of dots (organelle)/number of dots (oocyte)] × 100 (%) (Weibel et al., Reference Weibel, Kistler and Scherle1966).
Organelles included in the present evaluation were: cortical vesicles (CV), mitochondria, dictyosomes, lysosomes, medium-sized vesicles containing granular materials (VZP), smooth endoplasmic reticulum (SER) small vesicles (SER-SV), SER medium vesicles (SER-MV), SER large vesicles (SER-LV), SER very large vesicles (SER-VLV), SER isolated tubules (SER-IT), SER tubular aggregates (aSERT), and total SER without VLV (SV, MV, LV, IT, aSERT).
Each oocyte was divided into three regions, from the oolemma up to the cell centre: cortex (5 μm), subcortex (5–10 μm), and IC (>10 μm) (Figure 2A). A stereological procedure was adopted, applying the formula Vv (organelle, cortex/subcortex/IC) = [number of points (organelle)/number of points (cortex/subcortex/IC)] × 100 (%) (Pires-Luís et al., Reference Pires-Luís, Rocha, Bartosch, Oliveira, Silva, Barros, Sá and Sousa2016).

Figure 1. Metaphase II (MII) oocyte. (A) Live cell observed in the inverted microscope and (B, C) its correspondent semithin sections. The oocyte (O) was separated from the zona pellucida (ZP) by the perivitelline space (pvs), where resided the first polar body (PB1). In all cases, the ooplasm presented a homogeneous appearance. In semithin sections, all oocytes presented a metaphase place (mp) slightly dislocated towards the upper inner region. Chromosomes (chr) and cortical vesicles (cv) appeared evident in the PB1. The oocyte surface displayed numerous microvilli (mv). In the ooplasm, cortical vesicles (cv), mitochondria (mi) and large vesicles (V) can be observed. Bars: (A) 20 μm; (B, C) 10 μm.
Statistical analysis was performed using Microsoft® Excel 2022 and SPSS version 27.0 software (IBM Corp, Foster City, California, CA, USA). The results are presented as mean, standard error of the mean (SEM = standard deviation/n1/2), and coefficient of variation (CVar = standard deviation/mean). Normal distribution was tested using the Kolmogorov–Smirnov test. The samples did not follow a normal distribution, and therefore non-parametric tests were used. To compare the means of Vv (organelle, oocyte), Vv (organelle, cortex), Vv (organelle, subcortex) and Vv (organelle, IC) the Kruskal–Wallis test and the Mann–Whitney U-test with Bonferroni correction were used. The level of statistical significance was set at P < 0.05.
Results
When observed under the inverted microscope, live MII oocytes displayed a rounded shape and a uniform fine granular cytoplasm, being surrounded by a thick translucent zona pellucida (ZP) and separated from the ZP by a short and narrow perivitelline space, where the PB1 was located (Figure 1A). In semithin sections, the oocyte evidenced the same structures, being additionally evident microvilli, CV and large vesicles. The oocyte metaphase II plate was also discernible, being placed slightly away from the cortical region. The PB1 showed chromosomes and the characteristic organelles of the cortical ooplasm (Figure 1B,C).

Figure 2. Ultrastructural images of metaphase II (MII) oocytes. (A, B) MII oocytes, showing the three regions, cortex (C), subcortex (SC) and inner (IC) cytoplasm. The oocyte surface is separated from the zona pellucida (ZP) by the perivitelline space (pvs), which contains oocyte microvilli (mv). Rows of cortical vesicles (cv) are observed under the oolemma and as isolated cortical vesicles in the cortex and subcortex regions. In the ooplasm mitochondria (mi) and smooth endoplasmic reticulum (SER) small (SV) and medium (MV) sized vesicles and isolated tubules (IT) can be observed. SER vesicles were barely associated with mitochondria. (B) The first polar body (PB1) exhibits chromosomes (chr) and cortical ooplasm elements, such as cortical vesicles (cv) and isolated SER tubules (IT). (C) The chromosomes (chr) of the oocyte metaphase II plate appeared slightly displaced towards the upper inner region. Bars: 1 μm.
At the ultrastructural level, the oocyte surface exhibited numerous microvilli (Figure 2A,B), and in the perivitelline space the PB1 evidenced chromosomes and cortical ooplasm characteristics, including CV (Figure 2B). In the ooplasm, rows of electron-dense CV were observed under the oolemma and as isolated vesicles in the oocyte subcortex (Figure 2A). The metaphase II plate was slightly displaced towards the upper IC (Figure 2C). Mitochondria, SER-IT, SER-SV, SER-MV (Figure 2A) and SER-LV (Figure 3A) were observed in all three regions of the oocyte. The SER-VLV were found in the IC of one oocyte (Figure 3B). The aSERT was observed in all oocyte regions and also in one oocyte (Figure 4A,B). The VZP were observed in the oocyte cortex of two oocytes (Figure 4C). Lysosomes were observed in the cortex and inner ooplasm (Figure 4D). Dictyosomes were observed in the subcortex and IC of two oocytes (Figure 4E). The oocyte surface contained numerous tiny coated vesicles (Figure 4F). Multivesicular bodies, annulate lamellae, rough endoplasmic reticulum cisternae, lipid droplets and polyribosomes were not observed.

Figure 3. Ultrastructural images of metaphase II oocytes at the inner oocyte region. (A) In the inner cytoplasm mitochondria (mi) and smooth endoplasmic reticulum (SER) small (SV), medium (MV) and large (LV) sized vesicles and isolated tubules (IT) can be observed. Note that SER vesicles are barely associated with mitochondria. (B) The inner cytoplasm also presented SER very large vesicles (VLV), with their characteristic bridges (*).

Figure 4. Ultrastructural images of metaphase II oocytes. (A, B) Aggregates of smooth endoplasmic reticulum tubules (aSERT) in cortical (A) and inner (B) oocyte regions, surrounded by mitochondria (mi). (C) Vesicles containing granular materials (VZP) in the oocyte cortical region. (D) Lysosome (LY) in the inner ooplasm. (E) Dictyosome (Di) in the inner ooplasm. (F) The oocyte surface showed numerous tiny coated vesicles (arrows). ZP: zona pellucida; pvs: perivitelline space; mv: microvilli; SV, LV: small (SV) and large (LV) smooth endoplasmic reticulum vesicles. Bars: (A, C, D) 1 μm; (B) 1 μm; (E, F) 0.5 μm.
The present stereological analysis was performed on 204 photographs, with 96 representations of the cortex, 96 of the subcortex and 176 of the IC. The following organelles were quantified: mitochondria, dictyosomes, lysosomes, CV, VZP and SER elements (SER-SV, SER-MV, SER-LV, SER-VLV, SER-IT and aSERT).
The most predominant organelles in the total oocyte (Table 1) were SER-SV (Vv 8.84%), SER-IT (Vv 7.16%) and mitochondria (Vv 5.49%). This was followed by SER-MV (Vv 2.01%), LV-SER (Vv 0.79%), aSERT (Vv 0.61%), lysosomes (Vv 0.33%), CV (Vv 0.31%), SER-VLV (Vv 0.30) and dictyosomes (Vv 0.01%).
Table 1. Relative volume (Vv) of organelle per total MII oocyte and per MII oocyte regions: cortex, subcortex and inner cytoplasm

Results presented as mean, SEM (standard error of the mean: standard deviation/n1/2), and CVar (coefficient of variation: standard deviation/mean).
Mi: mitochondria; Di: dictyosomes; Ly: lysosomes; CV: cortical vesicles; VZP: vesicles containing zona pellucida-like material; SER: smooth endoplasmic reticulum; SV: SER small vesicles; MV: SER medium vesicles; LV: SER large vesicles; VLV: SER very large vesicles; IT: SER isolated tubules; aSERT: SER tubular aggregates.
The most common organelles per oocyte region were (Table 1): in the cortex, SER-SV (Vv 9.67%) followed by SER-IT (Vv 8.19%) and mitochondria (Vv 5.66%); in the subcortex, SER-SV (Vv 8.74%), SER-IT (Vv 7.22%) and mitochondria (Vv 5.79%); and in the IC, SER-SV(Vv 8.40%), SER-IT (Vv 7.2%) and mitochondria (Vv 5.33%).
The three oocyte regions were compared using the Kruskal–Wallis test (Table 2) and no significant differences were found for mitochondria (P = 0.677), dictyosomes (P = 0.317), SER-SV (P = 0.677), SER-MV (P = 0.102), SER-VLV (P = 0.368), SER-IT (P = 0.336) and aSERT (P = 0.980). Conversely, significant differences were found for lysosomes (P = 0.003), CV (P = 0.002), VZP (P = 0.032) and SER-LV (P = 0.009). Strict pairwise comparisons for the three regions using the Mann–Whitney U-test, Bonferroni corrected (Table 2), showed that there were significant differences in the Vv of CV between the three regions of oocytes (P = 0.008), lysosomes showed significant differences between cortex vs IC and subcortex vs IC (P = 0.008), and SER-LV between cortex vs IC (P = 0.032) and subcortex vs IC (P = 0.008).
Table 2. Comparison of the means of organelle relative volume (Vv) between MII oocyte regions: Vv (organelle, cortex), Vv (organelle, subcortex) and Vv (organelle, inner cytoplasm)

* Significance set at P < 0.05
Mi: mitochondria; Di: dictyosomes; Ly: lysosomes; CV: cortical vesicles; VZP: vesicles containing zona pellucida-like material; SER: smooth endoplasmic reticulum; SV: SER small vesicles; MV: SER medium vesicles; LV: SER large vesicles; VLV: SER very large vesicles; IT: SER isolated tubules; aSERT: SER tubular aggregates.
Note: Mann–Whitney U-test was performed when the Kruskal–Wallis test gave a significant P-value.
Discussion
Donor oocytes are an important procedure in ART and in the last 30 years have become a more prevalent therapy of choice for many infertile patients (Yeh et al., Reference Yeh, Steward, Dude, Shah, Goldfarb and Muasher2014). This study is based on the use of donor oocytes from a young woman by a recipient woman in ART procedures. Indications for donor oocytes include advanced reproductive age, diminished ovarian reserve, recurrent poor oocyte or embryo quality in IVF attempts, hypothalamic hypogonadism, and in some cases with the possibility of transmitting a genetic defect (Taylor, Reference Taylor and Stuart2019; Sciorio et al., Reference Sciorio, Antonini and Engl2021; Williams et al., Reference Williams, Ellis, Wilkinson, Kramer, Datta and Guzick2022). In oocyte donor cycles, in which ovarian hyperstimulation is used, it is necessary that nuclear and cytoplasmic maturation occurs in a coordinated manner to ensure optimal conditions for subsequent fertilization (Conti and Franciosi, Reference Conti and Franciosi2018), but which does not always occur, resulting in nuclear, cytoplasmic or extracytoplasmic morphological abnormalities (Ten et al., Reference Ten, Mendiola, Vioque, de Juan and Bernabeu2007; Setti et al., 2021).
In previous works we presented the stereological analysis of human immature oocytes at the GV stage (Pires-Luís et al., Reference Pires-Luís, Rocha, Bartosch, Oliveira, Silva, Barros, Sá and Sousa2016) and MI stage (Coelho et al., Reference Coelho, Pires-Luís, Oliveira, Alves, Leal, Cunha, Barreiro, Barros, Sá and Sousa2020). The present study was designed to apply the same technique to mature MII oocytes. To this end, we obtained 13 surplus donor MII oocytes, retrieved from six donors during oocyte donor programmes. In one case, the oocyte donor donated five oocytes for research, three cases gave two oocytes each, and two cases gave one oocyte each. In all cases, nuclear maturity was verified through inspection under an inverted microscope (live oocytes), which confirmed the presence of the first polar body, and in semithin sections (fixed oocytes), which confirmed the presence of the first polar body and the metaphase II plate. Also, in all cases, cytoplasmic integrity was verified through inspection under an inverted microscope and in semithin sections, which confirmed the absence of intracytoplasmic or extracytoplasmic dimorphisms. We started with the donor that provided five oocytes, but TEM observations revealed cytoplasmic immaturity characteristics. Thereafter, we looked at the remaining MII oocytes from the donor to ensure their cytoplasmic maturity. However, all five MII oocytes revealed cytoplasmic immaturity, evidencing an unconformity between nuclear and cytoplasmic maturity. The current study focuses on the examination of these five oocytes obtained from a single donor.
Previous ultrastructural studies of mature MII oocytes showed the presence of PB1 in the perivitelline space, containing chromosomes and the organelles of the cortical ooplasm (Zamboni et al., Reference Zamboni, Thompson and Smith1972; Sathananthan, Reference Sathananthan1994). An organized metaphase II plate could also be observed in the cortical/subcortical region near the PB1 (Zamboni et al., Reference Zamboni, Thompson and Smith1972; Sathananthan, Reference Sathananthan2013). The oocyte surface presented numerous microvilli (Morimoto, Reference Morimoto2009) and CV formed one or more layers next to the oolemma (Sathananthan and Trounson, Reference Sathananthan and Trounson1982; Motta et al., Reference Motta, Nottola, Micara and Familiari1988), with some cortical regions presenting a higher density of CV than others (Sathananthan and Trounson, Reference Sathananthan and Trounson1982; Sousa and Tesarik, Reference Sousa and Tesarik1994; El-Shafie et al., Reference El-Shafie, Windt, Kitshoff, McGregor, Sousa, Wranz, Kruger, El-Shafie, Sousa and Kruger2000; Sousa et al., Reference Sousa, Oliveira, Barros, Barros and Sá2016b). The oocyte cortex and subcortex were enriched in SER vesicles of various sizes, SER-IT and aSERT (Sousa and Tesarik, Reference Sousa and Tesarik1994; El-Shafie et al., Reference El-Shafie, Windt, Kitshoff, McGregor, Sousa, Wranz, Kruger, El-Shafie, Sousa and Kruger2000; Sathananthan, Reference Sathananthan2013; Sousa et al., Reference Sousa, Oliveira, Barros, Barros and Sá2016b). Mitochondria were described as evenly distributed, surrounding most SER vesicles and aSERT, and forming necklace-like complexes (Sundström et al., Reference Sundström, Nilsson, Liedholm and Larsson1985; Motta et al., Reference Motta, Nottola, Micara and Familiari1988, Reference Motta, Nottola, Makabe, Heyn and Jansen2000; El-Shafie et al., Reference El-Shafie, Windt, Kitshoff, McGregor, Sousa, Wranz, Kruger, El-Shafie, Sousa and Kruger2000; Sathananthan and Trounson, Reference Sathananthan and Trounson2000; Morimoto, Reference Morimoto2009; Nottola et al., Reference Nottola, Macchiarelli and Familiari2014; Sousa et al., Reference Sousa, Oliveira, Barros, Barros and Sá2016b; Trebichalská et al., Reference Trebichalská, Kyjovská, Kloudová, Otevřel, Hampl and Holubcová2021). Lysosomes were also described (Sathananthan, Reference Sathananthan, Trounson and Gosden2003; Trebichalská et al., Reference Trebichalská, Kyjovská, Kloudová, Otevřel, Hampl and Holubcová2021), and dictyosomes (Sathananthan et al., Reference Sathananthan, Selvaraj, Girijashankar, Ganesh, Selvaraj and Trounson2006; Nottola et al., Reference Nottola, Macchiarelli and Familiari2014) and annulate lamella (Sathananthan et al., Reference Sathananthan, Selvaraj, Girijashankar, Ganesh, Selvaraj and Trounson2006) were rarely observed. Some organelles and cytoplasmic structures were not found in mature MII oocytes, such as rough endoplasmic reticulum and multivesicular bodies (Sundström et al., Reference Sundström, Nilsson, Liedholm and Larsson1985), polyribosomes and lipid droplets (Sathananthan, Reference Sathananthan1994), just as in donor oocytes (Trebichalská et al., Reference Trebichalská, Kyjovská, Kloudová, Otevřel, Hampl and Holubcová2021).
We here describe the presence of cortical tiny coated vesicles, which were previously observed in GV and MI oocytes (Pires-Luís et al., Reference Pires-Luís, Rocha, Bartosch, Oliveira, Silva, Barros, Sá and Sousa2016; Coelho et al., Reference Coelho, Pires-Luís, Oliveira, Alves, Leal, Cunha, Barreiro, Barros, Sá and Sousa2020), and in mature MII oocytes (El-Shafie et al., Reference El-Shafie, Windt, Kitshoff, McGregor, Sousa, Wranz, Kruger, El-Shafie, Sousa and Kruger2000; Sousa et al., Reference Sousa, Oliveira, Barros, Barros and Sá2016b), indicating exchanges of materials with the exterior by receptor-mediated endocytosis (Hinduja et al., Reference Hinduja, Kumar and Anand Kumar1990; Pires-Luís et al., Reference Pires-Luís, Rocha, Bartosch, Oliveira, Silva, Barros, Sá and Sousa2016; Sousa et al., Reference Sousa, Oliveira, Barros, Barros and Sá2016b; Coelho et al., Reference Coelho, Pires-Luís, Oliveira, Alves, Leal, Cunha, Barreiro, Barros, Sá and Sousa2020).
Our present observations confirm those previous findings, but several characteristics of immaturity were also observed in the ooplasm. These immature characteristics were the following: CV were observed in the oocyte cortex and subcortex, as in GV and MI oocytes (Pires-Luís et al., Reference Pires-Luís, Rocha, Bartosch, Oliveira, Silva, Barros, Sá and Sousa2016; Coelho et al., Reference Coelho, Pires-Luís, Oliveira, Alves, Leal, Cunha, Barreiro, Barros, Sá and Sousa2020), being that they were expected to be only present in the cortical ooplasm; granular vesicles, suspected of containing ZP-like materials, were observed in the oocyte cortex, as in GV oocytes (Pires-Luís et al., Reference Pires-Luís, Rocha, Bartosch, Oliveira, Silva, Barros, Sá and Sousa2016), and although observed in only two oocytes (oocytes 2 and 3), it was expected they would not be found similarly to MI oocytes (Coelho et al., Reference Coelho, Pires-Luís, Oliveira, Alves, Leal, Cunha, Barreiro, Barros, Sá and Sousa2020); dictyosomes were observed in the oocyte subcortex, as in GV oocytes (Pires-Luís et al., Reference Pires-Luís, Rocha, Bartosch, Oliveira, Silva, Barros, Sá and Sousa2016) and in the IC, as in GV and MI oocytes (Pires-Luís et al., Reference Pires-Luís, Rocha, Bartosch, Oliveira, Silva, Barros, Sá and Sousa2016; Coelho et al., Reference Coelho, Pires-Luís, Oliveira, Alves, Leal, Cunha, Barreiro, Barros, Sá and Sousa2020), and although observed in only two oocytes (oocytes 1 and 3), they were expected to be not found, although there are two previous reports in mature MII oocytes that observed rare dictyosomes (Sathananthan et al., Reference Sathananthan, Selvaraj, Girijashankar, Ganesh, Selvaraj and Trounson2006; Nottola et al., Reference Nottola, Macchiarelli and Familiari2014); SER-VLV were observed in the IC in only one oocyte (oocyte 4), but these are a GV characteristic (Pires-Luís et al., Reference Pires-Luís, Rocha, Bartosch, Oliveira, Silva, Barros, Sá and Sousa2016); aSERT were missing in oocytes 1–4, and the oocyte exhibiting aSERT (oocyte 5) presented this organelle in all three oocyte regions, as in MI oocytes (Coelho et al., Reference Coelho, Pires-Luís, Oliveira, Alves, Leal, Cunha, Barreiro, Barros, Sá and Sousa2020). The aSERT in mature MII oocytes are limited to the oocyte cortex and subcortex, being critical for the oocyte to develop calcium oscillations at fertilization (Sousa et al., Reference Sousa, Barros and Tesarik1996, Reference Sousa, Barros, Silva and Tesarik1997), without which the oocyte is not activated and no embryo development occurs (Dale et al., Reference Dale, Wilding, Coppola and Tosti2010).
Therefore, all oocytes exhibited abnormal cortical vesicle positioning in the subcortex and all presented absence or abnormal positioning of aSERT. Additionally, in all cases, mitochondria appeared barely associated with SER-MV and SER-LV, and the metaphase II plate was slightly displaced from the cortical/subcortical region towards the IC. This renders the ooplasm of these donor MII oocytes immature.
By using a quantitative approach, when MII oocytes were compared in their different regions (Table 2), it was shown that there were no significant differences in the Vv of mitochondria among the three oocyte regions, corroborating what was reported in descriptive studies, in which a homogeneous distribution throughout the ooplasm was observed. Although mitochondria were described as the most predominant organelle in the ooplasm (Motta et al., Reference Motta, Nottola, Micara and Familiari1988, Reference Motta, Nottola, Makabe, Heyn and Jansen2000; Sathananthan and Trounson, Reference Sathananthan and Trounson2000; Trebichalská et al., Reference Trebichalská, Kyjovská, Kloudová, Otevřel, Hampl and Holubcová2021), this was not observed in the present study (Table 2), with quantitative analyses showing that the SER is the most predominant organelle (Table 1) (Mi Vv 5.49% vs total SER Vv 26.8%).
Using strict pairwise comparisons for the three oocyte regions and the Mann–Whitney U-test, Bonferroni corrected (Table 2), significant differences were observed for the following and the SER-LV presented a higher prevalence in the IC. This finding contradicts morphological study descriptions, in which lysosomes and SER-LV were seen to be distributed uniformly (Zamboni et al., Reference Zamboni, Thompson and Smith1972; Sundström et al., Reference Sundström, Nilsson, Liedholm and Larsson1985; Motta et al., Reference Motta, Nottola, Micara and Familiari1988).
Applying a stereological analysis similar to that previously used in human immature oocytes, GV (Pires-Luís et al., Reference Pires-Luís, Rocha, Bartosch, Oliveira, Silva, Barros, Sá and Sousa2016) and MI (Coelho et al., Reference Coelho, Pires-Luís, Oliveira, Alves, Leal, Cunha, Barreiro, Barros, Sá and Sousa2020), it was possible to compare quantitatively immature oocytes with the present studied MII oocytes.
Comparing GV with MII oocytes (Table 3), no significant differences were observed in the Vv of mitochondria, lysosomes, CV, VZP and SER-LV. For mitochondria, the Vv was lower in MII oocytes, except in the cortex, and was only significantly lower in the IC; for lysosomes and VZP, no significant differences in the Vv were found along all oocyte regions; in relation to CV, the Vv were similar in the cortex and subcortex; the significant difference observed in the IC were due to the absence of CV in that region of MII oocytes; for SER-LV, the Vv were similar but nevertheless higher in the cortex and IC of MII oocytes, and significantly lower in the subcortex of MII oocytes. Significant differences in all oocyte regions were observed in the Vv of dictyosomes, SER-SV, SER-MV, SER-VLV, SER-IT and total SER. For dictyosomes, the Vv was lower in all regions of MII oocytes; for SER-SV, the Vv was higher in all regions of MII oocytes; for SER-MV, the Vv was higher in all regions of MII oocytes, although not significant in the oocyte subcortex; for SER-VLV, the Vv was lower in all regions of MII oocytes; for SER-IT, the Vv was higher in all regions of MII oocytes; and for total SER (P < 0.001), the Vv was higher in all regions of MII oocytes.
Table 3. Comparison of the means of organelle relative volume (Vv) per oocyte region (organelle, cortex), Vv (organelle, subcortex) and Vv (organelle, inner cytoplasm) between GV oocytes and MII oocytes
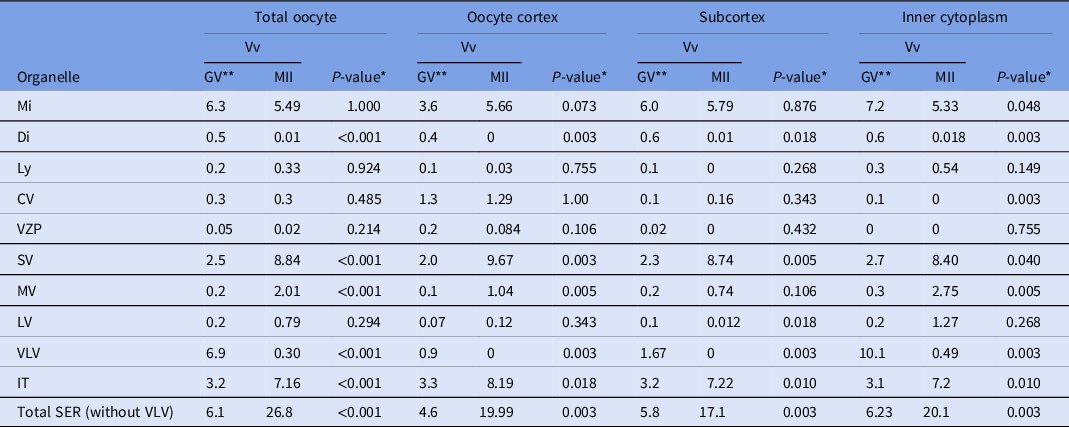
* Significance set at P < 0.05.
** Data are presented as: organelle relative volume (Vv) found in GV oocytes (from Pires-Luís et al., Reference Pires-Luís, Rocha, Bartosch, Oliveira, Silva, Barros, Sá and Sousa2016), organelle Vv found in MII oocytes (from Table 1), and the P-value obtained after comparison between the GV-organelle Vv and the MII-organelle Vv.
GV: prophase I oocytes (germinal vesicle oocytes); MII: metaphase II oocytes.
Mi: mitochondria; Di: dictyosomes; Ly: lysosomes; CV: cortical vesicles; VZP: vesicles containing zona pellucida-like material; SER: smooth endoplasmic reticulum; SV: SER small vesicles; MV: SER medium vesicles; LV: SER large vesicles; VLV: SER very large vesicles; IT: SER isolated tubules.
Note: aSERT (SER tubular aggregates) not included as not found in GV oocytes.
Comparing MI with MII oocytes (Table 4), no significant differences in the Vv were found in relation to dictyosomes (although not significant, the Vv was higher in MII oocytes in the subcortex and IC), lysosomes (except for a significantly lower Vv in the cortex of MII oocytes and a non-significant higher Vv in the IC), CV (although not significant, the Vv was higher in MII oocytes, except for a significantly lower Vv in the IC) and SER-LV (although not significant, the Vv was higher in MII oocytes). Significant differences in the Vv were observed regarding mitochondria, SER-SV, SER-MV, SER-IT, aSERT and total SER. The Vv of mitochondria, SER-IT and aSERT were significantly lower in MII oocytes only when evaluating the total oocyte, with all regions displaying non-significantly lower Vv in MII oocytes. The Vv of total SER was significantly higher when evaluating the total oocyte and IC of MII oocytes, with non-significantly higher Vv in the cortex and subcortex of MII oocytes. The Vv of SER-SV was significantly higher in all regions of MII oocytes. The Vv of SER-MV was significantly lower in the cortex and subcortex of MII oocytes, and not significantly lower in the IC of MII oocytes.
Table 4. Comparison of the means of organelle relative volume (Vv) per oocyte region (organelle, cortex), Vv (organelle, subcortex) and Vv (organelle, inner cytoplasm) between MI oocytes and MII oocytes

* Significance set at P < 0.05.
** Data are presented as: organelle relative volume (Vv) found in MI oocytes (from Coelho et al., Reference Coelho, Pires-Luís, Oliveira, Alves, Leal, Cunha, Barreiro, Barros, Sá and Sousa2020), organelle Vv found in MII oocytes (from Table 1), and the P-value obtained after comparison between the MI-organelle Vv and the MII-organelle Vv.
Mi: mitochondria; Di: dictyosomes; Ly: lysosomes; CV: cortical vesicles; SER: smooth endoplasmic reticulum; SV: SER small vesicles; MV: SER medium vesicles; LV: SER large vesicles; IT: SER isolated tubules: aSERT: SER tubular aggregates.
Note: VZP (vesicles containing zona pellucida-like material) and VLV (SER very large vesicles) not included. as not found in MI oocytes.
Considering all these quantitative data, a gradual increase in SER elements, as described in morphological studies (Sundström et al., Reference Sundström, Nilsson, Liedholm and Larsson1985; Nottola et al., Reference Nottola, Macchiarelli and Familiari2014) and a decrease in dictyosomes (Sathananthan, Reference Sathananthan, Trounson and Gosden2003; Sathananthan et al., Reference Sathananthan, Selvaraj, Girijashankar, Ganesh, Selvaraj and Trounson2006), was noted specifically from GV to MI stages. However, an increase in CV was expected along with oocyte maturation, as well as in aSERT and mitochondria–SER complexes in the oocyte cortex (Nottola et al., Reference Nottola, Macchiarelli and Familiari2014), which was not observed. Therefore, the cytoplasm of the oocytes analyzed in this study exhibited both GV and MI features, implying that they contained an immature cytoplasm.
Although a stereological analysis of mature (nuclear and cytoplasmic) MII oocytes is in progress, we think that the presentation of this anomaly (discrepant nuclear and cytoplasmic maturation in MII donor oocytes) deserves anticipated publication due to its immediate clinical importance.
In conclusion, we describe a case of donor MII oocytes exhibiting nuclear maturity simultaneously with cytoplasmic immaturity. If used in a donor programme, these oocytes would not be fertilized, and therefore this finding is the first demonstration of a possible cause of ART failure in oocyte donor cycles, in which oocyte quality is usually taken for granted.
Ethics approval and consent to participate
The authors declare that they followed all rules of ethical conduct regarding originality, data processing and analysis, duplicate publication and biological material. All authors critically read the final manuscript and agreed with the author order.
Ethical guidelines were followed conducting the research. According to the National Law on Medically Assisted Procreation (Law 32/2006) and the National Council for Medically Assisted Procreation guidelines (2018), the use of clinical databases and surplus gametes for research may be used under strict individual anonymity and after patient written informed consent. Surplus donor oocytes were used after the patient signed an informed consent agreeing to share their own gamete samples for the present study. The present laboratorial experiments were executed under the Joint Ethics Committee of the Hospital and University, CHUP/ICBAS approval number 2019/CE/P017 (266/CETI/ ICBAS). This work did not involve human or animal experiments and therefore the provisions of the Declaration of Helsinki as revised in Tokyo 2004 do not apply.
Data availability statement
The authors confirm that the data supporting the findings of this study is available upon request.
Acknowledgements
Not applicable.
Author contributions
T.S. was responsible for the stereology and statistic work, data analysis, critical discussion, a draft of the manuscript and a critical review of the final manuscript. A.S.P.L. was responsible for study design, stereology and statistic training, electron microscopy image acquisition, data analysis, critical discussion and critical review of the final manuscript. A.A. and E.O. were responsible for oocyte processing to electron microscopy and critical review of the final manuscript. C.L. was responsible for the embryologic work, donor oocyte selection and critical review of the final manuscript. M.F. was responsible for the psychological evaluation of the donor and the critical review of the final manuscript. E.V.F. and M.B. were responsible for clinical donor selection, ovarian stimulation, oocyte retrieval and critical review of the final manuscript. A.M.C. was responsible for critical review of the final manuscript. R.S. was responsible for critical discussion and review of the final manuscript. M.S. was responsible for study conceptualization, electron microscopy image acquisition, data analysis, critical discussion and writing of the final manuscript. All authors have read and agreed to the order of appearance and the published version of the manuscript.
Funding
ICBAS-UP and UMIB/ITR. The UMIB-Unit for Multidisciplinary Research in Biomedicine is funded by the Foundation for Science and Technology (FCT) Portugal (grant numbers UIDB/00215/2020 and UIDP/00215/2020) and the ITR-Laboratory for Integrative and Translational Research in Population Health (LA/P/0064/2020).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Institutional review board statement
Not applicable.
Informed consent statement
Not applicable








