Introduction
Cowpea (Vigna unguiculata L.) is one of the most important legume crops that is fairly adapted to harsher environments especially drought and high temperature. In India it is used as a dual-purpose crop used for food and fodder. In the North-Western Himalayan region especially Kashmir valley it is a niche crop that fetches premium price on account of its broader adaptability and nutritional value viz. high protein content as well as its ability to fix nitrogen. However, a number of production constraints such as pod shattering severely implicate its yield potential especially under changing climate with drier and warmer days around maturity period of the crop. Cowpea was domesticated in Africa from its wild ancestor V. unguiculata var. dekindtiana) some 5000–6000 years ago (Coulibaly et al., Reference Coulibaly, Pasquet, Papa and Gepts2002). The loss of yield from pod shattering can be either on account of the pre-harvest pod shattering in the field, and the yield loss from pod shattering during manual or mechanical stress. This problem is exacerbated in dry arid environments where pods become brittle and fracture easily. There is no published information available in cowpea in terms of losses caused by shattering, but losses as high as 20–60% have been reported in soybean, broad bean, brassica and canola (Tiwari and Bhatnagar, Reference Tiwari and Bhatnagar1991; Price et al., Reference Price, Hobson, Neale and Bruce1996; Child et al., Reference Child, Chauvaux, John, Onckelen and Ulvskov1998; Dong et al., Reference Dong, Jahufer, Dong, Wang and Liu2016). Pod shattering also has a metabolic cost as it limits the seed size (Murgia et al., Reference Murgia, Attene, Rodriguez, Bitocchi, Bellucci, Fois and Rau2017). In fact, 100-seed weight of shattering resistant types in V. unguiculata subsp. sesquipedalis was reported to be higher than the wild types (Takahashi et al., Reference Takahashi, Kongjaimun, Muto, Kobayashi, Kumagai, Sakai and Naito2019). The domesticated cowpea has fairly lesser level of pod shattering, despite the fact that the level of shattering in cultivated germplasm is fairly high with persistence of shattering in some varieties. As such shattering assumes significant importance as a cause of yield losses especially under dry hot environments (Zhang and Singh, Reference Zhang and Singh2020; Parker et al., Reference Parker, Lo and Gepts2021). Therefore, there is an urgent need to develop shattering resistant varieties to avoid such losses.
Despite the fact that human selection under domestication of crops has driven selection of desired plant traits including adaptive traits such as reduced shattering, plants have still retained fair amount of seed shattering to provide for ease of harvesting (Gregory and Ellis, Reference Gregory and Ellis2009; Flint-Garcia, Reference Flint-Garcia2013). In wild plants, shattering is desirable as it allows seed dispersal, but in commercial agriculture, it is undesirable as it leads to substantial loss of yields. Pod shattering is the most important domestication syndrome that has evolved independently in legume crops during the domestication process, and has driven adaptation of the legumes plants to diverse agro-ecosystem (Tang et al., Reference Tang, Cuevas, Das, Sezen, Zhou, Guo, Goff, Ge, Clemente and Paterson2013).
In cowpea, the major domestication driven changes have progressively reduced pod dehiscence and seed hardness. Also, there has been an increase in size of pods and seeds by increased rate of dry weight accumulation (Lush and Evans, Reference Lush and Evans1981; Andargie et al., Reference Andargie, Pasquet, Gowda, Muluvi and Timko2014). Shattering of pods is mainly governed by hygroscopic movements within the pod valves upon dehydration (Lo et al., Reference Lo, Muñoz-Amatriaín, Boukar, Herniter, Cisse, Guo, Roberts, Xu, Fatokun and Close2018). The accumulation of tension during dehydration causes splitting of the valves along their suture lines (Elbaum and Abraham, Reference Elbaum and Abraham2014). Shattering invariably occurs at the final stages of pod development and is triggered by senescence and desiccation of lignified cells in the pods (Roberts et al., Reference Roberts, Elliott and Gonzalez-Carranza2002). In cowpea, pod shattering has been found to be influenced by the thickness and strength of the fibre layer of the pod that generates twisting force (Takahashi et al., Reference Takahashi, Kongjaimun, Muto, Kobayashi, Kumagai, Sakai, Satou, Teruya, Shiroma, Shimoji, Hirano, Isemura, Saito, Baba-Kasai, Kaga, Samta, Tomooka and Naito2020).
Several pod physical traits such as pod wall weight and pod wall water content, pod number, thickness of the pod wall, and seed weight/pod weight ratio, pod thickness, pod length have been implicated in shattering response with most of the work reported from soybean and common bean (Kuai et al., Reference Kuai, Sun, Liu, Zhang, Zhou, Wu and Zhou2016; Krisnawati and Adie, Reference Krisnawati and Adie2017; Zhang et al., Reference Zhang, Tu, Liu and Liu2018; Krisnawati et al., Reference Krisnawati, Soegianto and Waluyo2020; Sofi et al., Reference Sofi, Mir, Bhat, Mir, Fatima, Rani and Zargar2022). The present study was the first comprehensive phenotyping study in Western Himalayan region using national core set of cowpea for pod shattering in relation to various pod physical traits with the broader aim of identifying sources of resistance to shattering using a modified screening method that is based on weighted level scores. The broad hypothetical framework of present study was that the pod shattering has a definite physical basis in terms of being affected by pod physical traits.
Materials and methods
Site of the experiment
The experiment was conducted in 2022 and 2023 at the research fields of Division of Genetics and Plant Breeding, Faculty of Agriculture Wadura, SKUAST-K, Sopore (34°17′ North and 74°33′ E at an altitude of 1594 masl). The experimental site soil is a typical inceptisol with predominantly clay loam texture. The pH is almost neutral (7.2), with organic carbon 0.65%, electrical conductivity of 0.18 dS/m and CEC of 16 meq/kg. All the accessions were grown as single rows of four-meter length, with a spacing of 20 cm × 60 cm, in an augmented block design with four checks. The minimum and maximum temperatures during the growing season in 2022 varied from 10.50–17.91 and 26–32°C and in 2023 it ranged from 9.10–18.01 and 23.2–33.50°C, respectively. The total precipitation during the crop period in 2022 and 2023 was 194.60 and 388.40 mm respectively. Data was recorded on ten competitive plants from each genotype for pod length, pod breadth, pod thickness, seeds per pod, 100-seed weight, pod weight, pod wall weight and shattering score as per the standard procedures (Sofi, Reference Sofi2019; Fatima et al., Reference Fatima, Rani, Shafi, Zaffar, Zargar and Sofi2023).
Experimental material
The material for the present study comprised of a core set of 248 lines from national cowpea core set developed by NBPGR (National Bureau of Plant Genetic Resources) including six checks (Shalimar cowpea-1, Shalimar cowpea-2, Local Bold-1, WFC-1, WFC-2 and WFC-3), representing diverse market classes in cowpea. The core set comprised of a vast diversity of growth habits, seed colour, shape and eye pattern. All the accessions were grown as single replicates in an augmented block design (ABD) with a 2.5 m row with spacing of 20 cm within rows and 60 cm between rows, except the checks that were replicated in each block. Each block of the ABD comprised of 31 accessions and six checks.
Crop management
The management practices were uniform and homogeneous and comprised of seed treatment with the fungicide (Captan 50WP) and the insecticide (Imidacloprid 17.8%) at the rate of 2 ml/kg seed, application of the pre-emergent herbicide Pendimethalin at a dose of 1.25 l/ha as well as timely manual weeding, recommended dose of fertilizers (NPK) comprising a basal dose and a top dressing of urea at the V3 stage (first open trifoliate leaf). The crop was irrigated intermittently to avoid drought stress that would have confounded the results. The pods were harvested manually at the R9 (maturation stage), when 95% of pods were physiologically mature.
Pod physical traits recorded in the present study
-
Pod length (cm): Average length of pods was calculated in cm using a measuring scale from base of pod to the tip except the beak if any
-
Pod breadth (cm): Average breadth of pods was calculated in mm using vernier calliper
-
Pod thickness (cm): Average thickness of pods in mm using vernier calliper
-
Seeds per pod: Number of seeds per pod averaged over 10 pods
-
100-seed weight (g): Weight of 100 seeds averaged over three randomly drawn samples.
-
Pod weight (g): Average weight of pods of including seeds using weighing balance in grams
-
Pod wall weight (mg): Average weight of pods excluding seeds using weighing balance in grams
-
Shattering score: Weighted average of number of pods in various types of shattering
Manual screening for pod shattering using random impact method
Field phenotyping of pod shattering requires fully grown plants, and it is a time-consuming and labour-intensive procedure (Kim et al., Reference Kim, Kim, Jung, Kang, Lee, Ha and Kang2020). Moreover, the fluctuations in weather parameters at the time of pod maturation cause bias in the results. However, a test procedure, namely Random Impact Apparatus (RIA) has been devised that exposes pods to random impacts in a similar manner to those that occur in the crop canopy during harvest (Bruce et al., Reference Bruce, Farrent, Morgan and Child2002; Murgia et al., Reference Murgia, Attene, Rodriguez, Bitocchi, Bellucci, Fois and Rau2017). In the present study, screening of pod shattering was done in laboratory using RIM method suggested by Murgia et al. (Reference Murgia, Attene, Rodriguez, Bitocchi, Bellucci, Fois and Rau2017) with modifications using an in house designed RIA comprising of a 20 cm diameter cylinder with six steel balls of 12 mm diameter. Random impact method (RIM) enables the rapid comparison of shattering response of fully mature pods from individual plants (Bruce et al., Reference Bruce, Farrent, Morgan and Child2002). The pods were harvested at physiological maturity and were equilibrated for moisture for 10 d in paper bags (20 × 10 cm) followed by oven drying at 80° for 2 d. Ten sampled pods were put in RIA and shaken on a shaker at 250 rpm for 10 s using a stopwatch. Each treatment was done in triplicate. The method subjects all samples to uniform conditions as well as manual pressure. In addition, Murgia et al. (Reference Murgia, Attene, Rodriguez, Bitocchi, Bellucci, Fois and Rau2017) observed that almost 98% of the variance for shattering was genetic and there was a very limited role of environmental factors influencing this trait.
Development of a modified screening index
We developed a modified screening index following the protocol for level of shattering as proposed by Murgia et al. (Reference Murgia, Attene, Rodriguez, Bitocchi, Bellucci, Fois and Rau2017). In the conventional method, the shattering score is usually estimated as percentage of shattered pods from a given sample, without any delineation of type of shattering. In this study, we measured the shattering response of pods in terms of 1–9 scale where 1 = indehiscent pods, 2 = fissured pods, 3 = split pods, 4 = split pods with 20% twist, 5 = split pods with 40% twist, 6 = split pods with 60% twist, 7 = split pods with 80% twist, 8 = split pods with 100% twist, and 9 = Abscised pods. In order to build the shattering score index, the number of pods in each ith class (type of shattering) was multiplied by the ith scale (numerical description of the scale) and the final shattering score (SS) was taken as weighted average of all the values as follows:

Results
Variability for pod physical traits and shattering score
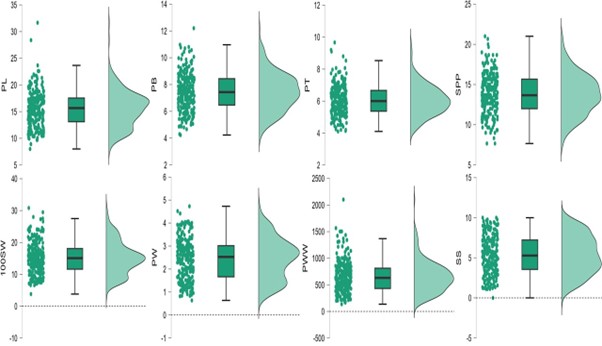
There was substantial variability in seven pod physical traits and shattering score in 254 genotypes of cowpea core collection indicating significant diversity of the material in respect of pod traits (Table 1, online Supplementary Figs. S1–S4). Pod length had a mean value of 15.56 cm (8.00–31.67 cm). Highest pod length was recorded in case IC471926 (31.67 cm), followed by IC58905 (28.40 cm) and WFC-3 (23.67 cm), whereas IC202710 has smallest pods measuring 8.00 cm. Pod breadth had a mean value of 7.44 mm (4.23–12.20 mm). Highest pod breadth was recorded in case EC724805 (12.20 mm), followed by EC723822 (10.97 mm) and EC724239 (10.80 mm), whereas pod breadth was smallest in case of IC397618 (4.23 mm). Pod thickness had a mean value of 6.06 mm with a range of 4.10–9.67 mm. Highest pod thickness was recorded in case IC413324 (9.67 mm), followed by EC244130 (9.20 mm) and EC724805 (9.10 mm), whereas pod thickness was smallest in case of IC383461 (4.10 mm). Pod weight had a mean value of 2.43 g with a range of 0.63 to 4.73 grams. Highest pod weight was recorded in case IC282020 (4.73 g), followed by IC413324 (4.52 g) and EC244025 (4.42 g) while as lowest value for pod weight was recorded in IC202710 (0.63 g). Pod wall weight had a mean value of 649.74 mg (136.00 to 2103 mg). Highest pod wall weight was recorded in case IC282020 (2103.00 mg), followed by EC725103 (1506.00 mg) and EC724319 (1504.00 mg) while as lowest value for pod wall weight was recorded in IC397618 (136.00 mg). Seeds per pod had a mean value of 13.67 with a range of 7.67–21.00. Highest number of seeds per pod were recorded in case of EC98661 (21.00), followed by EC98678 (20.67) and IC97807 (20.00), whereas the lowest number of seeds per pod were recorded in case of EC16205 and IC372722 (7.67). Similarly, for 100-seed weight a broader range of 3.80–30.82 g was observed with the highest 100-seed weight recorded in case of C-196 (30.82 g), followed by IC336836 (29.48 g) and EC18734 (28.00 g), whereas lowest 100-seed weight was recorded for EC738277 (3.80 g).
Table 1. Descriptive statistics of pod physical traits in cowpea core collection

PW, Pod weight; PL, Pod length; PB, Pod breadth; PT, Pod thickness; SPP, Seeds per pod; 100SW, 100 seed weight; PWW, Pod wall weight; SS, Shattering score.
There was substantial variability in pod shattering score in 254 genotypes of cowpea indicating significant diversity of the material in respect of shattering response (Fig. 1). Using the conventional scoring method of shattering response based on percentage of sampled pods shattered, regardless of the type of shattering, out of 254 genotypes, 18 were highly resistant, 22 were moderately resistant, 40 were moderately susceptible and 178 were highly susceptible. Shattering score had a mean value of 5.39 with a range of 1.00 in resistant genotypes to 10.00 in highly susceptible genotypes. Highest shattering score of 10.00, that indicates highest susceptibility to shattering, was recorded in case EC738122 and EC738159, followed by a score of 9.78 recorded for EC7511 and EC528687. One accession with a shattering score of 1 viz., EC332352 was completely indehiscent even under manual shattering force, followed by highly resistant accessions IC58905, IC413324 and IC361790 with a score of 1.11. Out of 254 genotypes, 34 were resistant (SS = 0–2.5), 83 were moderately resistant (2.51–5), 82 were moderately susceptible (5.1–7.5) and 55 were highly susceptible (SS = 7.51–10).

Figure 1. Response of 254 cowpea genotypes to manual shattering based on modified screening system.
Trait association among pod physical traits in cowpea core set
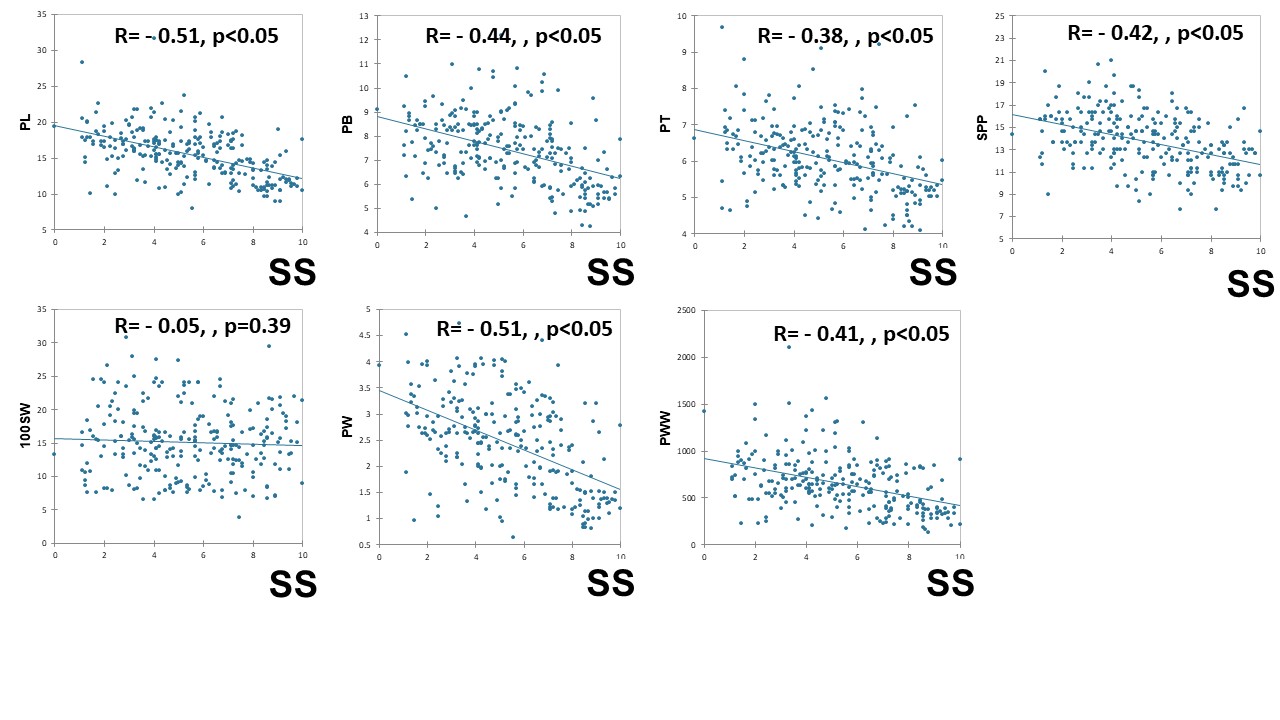
The trait associations of eight pod physical traits are depicted in Fig. 2. Shattering score had significant negative correlation with pod length (r = −0.513) followed by pod weight (r = −0.510), pod breadth (r = −0.440), seeds per pod (r = −0.420), pod wall weight (r = −0.410) and pod thickness (r = −0.383). Among other pod traits, significant positive correlations were recorded with notable correlations between pod breadth and pod weight (r = 0.830), followed by pod length and pod weight (r = 0.810), pod weight and pod thickness (r = 0.770), pod weight and pod wall weight (r = 0.760), pod breadth and pod thickness (r = 0.740), pod length and pod breadth (r = 0.700), pod length and pod wall weight (r = 0.640), pod breadth and pod thickness with pod wall weight (r = 0.620). There were no significant correlations recorded among pod traits except with shattering score.

Figure 2. Heat map of pod physical traits in cowpea, Where, PW, Pod weight; PL, Pod length; PB, Pod breadth; PT, Pod thickness; SPP, Seeds per pod; 100SW, 100 seed weight; PWW, Pod wall weight; SS, Shattering score.
Principal component analysis
The PCA is a useful data reduction technique that helps plant breeder to reduce the data dimensions and exclude the traits that either have non-significant contribution towards variation or have non-significant correlation with the trait of interest. In the present study PCA was done based on eight pod physical traits (Table 2) scored in the cowpea core set. The number of PCA was derived from correlation matrix and is equal to the number of traits. Based on the Eigen value and the cumulative variance accounted for the PCA, the PCA concentrated 69.60% variability in the first two principal components which were significant based on Eigen values greater than unity. The Eigen value was 4.497 for PC1 and 1.07 for PC2. Rest of the PCs were not considered as the Eigen value was less than unity. The first two PC's were used for construction of biplot that accounted for 69.60% of total variation (Fig. 3). The traits that significantly contributed to the PC1 were pod weight, followed by pod length, pod breadth, pod thickness and pod wall weight. The PC2 was largely contributed by 100-seed weight (Table 3).
Table 2. Eigen values and variances accounted by principal components


Figure 3. PCA biplot of various pod physical traits, Where, PW, od weight; PL, Pod length; PB, Pod breadth; PT, Pod thickness; SPP, Seeds per pod; 100SW, 100 seed weight; PWW, Pod wall weight; SS, Shattering score.
Table 3. Component loadings of pod physical traits

PW, Pod weight; PL, Pod length; PB, Pod breadth; PT, Pod thickness; SPP, Seeds per pod; 100SW, 100 seed weight; PWW, Pod wall weight; SS, Shattering score.
Discussion
In the present study substantial variability in pod physical traits in the cowpea core set was observed in respect of pod length, pod breadth, pod thickness, pod weight and pod wall weight as well as shattering score. Several earlier works have reported substantial diversity for pod length in cowpea ranging from 12.3–74.5 cm (Xu et al., Reference Xu, Wu, Muñoz-Amatriain, Wang, Wu, Hu and Li2017), 10.44–20.12 cm (Sofi et al., Reference Sofi, Mir, Bhat, Mir, Fatima, Rani and Zargar2022), 7.50–20.00 cm (Fatima, Reference Fatima2023). Pod length, breadth and pod wall weight have significant role in shattering response (Krisnawati and Adie, Reference Krisnawati and Adie2017). Pods with greater pod wall weight (thicker pods) are fairly resistant to shattering (Gioia et al., Reference Gioia, Giuseppina, Giovanna, Elisa, Benedettelli, Negri, Papa and Zeuli2013; Fatima, Reference Fatima2023) as the degree of pod coiling of pod walls is strongly influenced by thickness of the wall (Krisnawati and Adie, Reference Krisnawati and Adie2017; Takahashi et al., Reference Takahashi, Kongjaimun, Muto, Kobayashi, Kumagai, Sakai and Naito2019). The range of trait dispersion as depicted by range and C.V. value showed that highest C.V. value was observed in case of pod wall weight (35.40%) followed by shattering score (34.40%), pod weight and pod thickness (26.60%), 100-seed weight (23.80%) and pod length (21.80%), while as the lowest value of C.V. was observed in seeds per pod (18.20%).
Pod shattering score had a broad range from almost resistant (SS = 0) to completely shattered (SS = 10). Out of 254 genotypes, 34 were resistant (SS = 0–2.5), 83 were moderately resistant (SS = 2.51–5), 82 were moderately susceptible (5.1–7.5) and 55 were highly susceptible (SS = 7.51–10). Several earlier workers have reported substantial variation for shattering score in several legume species. Guo et al. (Reference Guo, Zhu, Li, Liu, Wu, Wang and Li2022) has reported wide variation of pod shattering score in Medicago ranging from 7.4–76%. Krisnawati and Adie (Reference Krisnawati and Adie2017) reported highly significant difference for pod shattering in soybean, indicated the high variation in shattering resistance among genotypes. Similarly, Murgia et al. (Reference Murgia, Attene, Rodriguez, Bitocchi, Bellucci, Fois and Rau2017) has also reported wide variation in a common bean diversity panel of 267 genotypes comprising a susceptible (MG38) and resistant (MIDAS) and their introgression lines with shattering percentage of 0–82.6% with a mean of 31.6%. Kaur et al. (Reference Kaur, Akhatar, Goyal, Kaur, Kaur, Mittal, Kumar, Banga and Banga2020) reported that black mustard was more susceptible to shattering as compared to Indian mustard and rapeseed and reported significant variation in terms of rupture energy. In our earlier studies on common bean pod shattering, Fatima et al. (Reference Fatima, Rani, Shafi, Zaffar, Zargar and Sofi2023) reported shattering score ranging from 0–10 with a mean of 6.098 in a common bean diversity panel. However, all these studies were based on conventional method of pod shattering screening without any distinction to different types of shattering.
The trait association revealed that shattering score had significant negative correlation with pod length followed by pod weight, pod breadth, seeds per pod, pod wall weight and pod thickness. Based on these trait associations it can be premised that pod length, pod weight, pod breadth, seeds per pod, pod wall weight and pod thickness are major drivers of shattering resistance in cowpea. Thus, longer pods with wider diameter and thicker pod wall (resulting in higher pod wall weight) are more resistant to shattering. Such pods have pod walls containing higher amounts of starch, cellulose, pectin and lignin that improve shattering (Fatima et al., Reference Fatima, Rani, Shafi, Zaffar, Zargar and Sofi2023). Similar results have been reported by Krisnawati et al. (Reference Krisnawati, Adie, Soegianto and Waluyo2019, Reference Krisnawati, Soegianto and Waluyo2020) in soybean and Fatima et al. (Reference Fatima, Rani, Shafi, Zaffar, Zargar and Sofi2023) in common bean. In fact, Krisnawati et al. (Reference Krisnawati, Soegianto and Waluyo2020) reported highly significant negative correlation between pod shattering and seeds per pod in soybean, but shattering had positive correlation with dorsal length of pods that indicates greater curvature of pods. Murgia et al. (Reference Murgia, Attene, Rodriguez, Bitocchi, Bellucci, Fois and Rau2017) also reported more pronounced pod shattering in lines with smaller pods, lower pod weight and pods with lower seed to pod ratio. The greater influence of pod length on shattering response in cowpea might be on account of parallel changes in in cowpea under domestication viz., increased strength of dehiscence zone in pod sutures, increase in size of pods and seeds as well as reduction in dehiscence (Lush and Evans, Reference Lush and Evans1981). Similarly, Kataliko et al. (Reference Kataliko, Kimani, Muthomi, Wanderi, Olubayo and Nzuve2019) reported that pod shattering resistance in soybean was negatively correlated with number of seeds per pod. The degree of pod coiling has a strong positive relationship with the pod wall thickness as reported by Takahashi et al. (Reference Takahashi, Kongjaimun, Muto, Kobayashi, Kumagai, Sakai, Satou, Teruya, Shiroma, Shimoji, Hirano, Isemura, Saito, Baba-Kasai, Kaga, Samta, Tomooka and Naito2020). However, some workers such as Suzuki et al. (Reference Suzuki, Fujino and Funatsuki2009) and Dong et al. (Reference Dong, Yan, Dong, Liu, Wang and Liu2017) reported that there was no significant correlation of shattering response with length, width, and thickness of pods and the thickness: width ratio was not associated with pod shattering.
The PCA concentrated 69.60% variability in the first two principal components which were significant based on Eigen values greater than unity. The traits that significantly contributed to the PC1 were pod weight, followed by pod length, pod breadth, pod thickness and pod wall weight. The PC2 was largely contributed by 100-seed weight.
In terms of GT biplot pod shattering score was significantly and negatively correlated with pod length, pod weight, pod breadth, seeds per pod, pod wall weight and pod thickness. The results are fairly in agreement with the correlation analysis even though sometimes the relationship based on PCA biplot are different than correlation analysis as it captures only a part of variation (69.60% in present case). This is the first report of multivariate analysis of pod physical traits in relation to pod shattering in cowpea. Tu et al. (Reference Tu, Liu, Wang, Li, Zhang, Liu and Herbert2019) used principal component analysis among various anatomical traits of ventral suture in soybean in relation to pod shattering and reported that first two axes explained 93.6% of the total variance in the shatter-susceptible and three shatter-resistant soybean cultivars. Similarly, Fatima (Reference Fatima2023) reported in common bean diversity panel that first two PC's accounted for 50.74% of variation largely contributed by pod wall thickness, breadth thickness ratio, string per cent, length breadth ratio, pod length, pod breadth, pod thickness, ventral pod length and filled pod weight.
The conventional screening methods as proposed by Murgia et al. (Reference Murgia, Attene, Rodriguez, Bitocchi, Bellucci, Fois and Rau2017) and one reported by Fatima (Reference Fatima2023) are efficient in delineating genotyping differences for pod shattering on the basis of number of pods shattered out of the sampled pods subjected to manual shattering pressure using a RIM apparatus. However, it is based on level of shattering and does not take into account various types of shattering such as fissured, split, twisted or abscised (online Supplementary Fig. S3). It has been observed that in several cases, fissured and partially split pods do not lead to complete loss of seed as compared to twisted and abscised pods. In order to quantify the relative contribution of each type of shattering response within sampled pods, we used an average of weighted level scores as described above. The method takes into account the number of pods in a sampled set from each class of shattering to derive the shattering score that is more reliable than earlier methods. We propose that this method should be used for screening shattering response under manual pressure methods to overcome the ambiguity of conventional methods. It creates fairly uniform conditions for pods as they are equilibrated for moisture and subjected to similar manual pressure.
Conclusion
The present study is the first comprehensive screening report of a large-scale germplasm evaluation for pod shattering. We used Random Impact Assessment, which is a useful approach for characterising large germplasm collections for traits like shattering, whose screening under filed conditions is implicated by weather changes as well as moisture status of pods. The method creates a fairly uniform screening system for shattering response and removes all subjectivities. We identified important pod physical attributes that are important determinants of shattering response in cowpea. In the present study, we screened a set of 254 cowpea genotypes of national core collection using pod physical traits to identify effective traits for improving shattering resistance. We identified pod length, pod weight, pod breadth, seeds per pod, pod wall weight and pod thickness as important drivers of shattering response in cowpea. We identified several shattering resistant genotypes (EC332352, IC471926, WFC-3, IC58905, IC413324 and IC361790, EC109493-2744-1, EC109493-1047-1, EC738109 and EC724319) that can be used in crossing programme to develop mapping populations for molecular characterization of pod shattering. As for the ideal trait combinations for improving shattering response breeders should select for relatively longer, thicker and straighter pods. The findings of the study are useful not only from the point of view of identifying potential sources of resistance to shattering, but can also be used to map genetic loci underlying shattering response in cowpea.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S1479262124000388.
Acknowledgements
The support of Dr Kuldeep Tripathi for providing the seeds of the national core set is acknowledged.
Competing interests
The authors declare they have no conflicts of interest.